 2019, Vol. 39
2019, Vol. 39文章信息
- 成向荣, 邢文黎, 苑海静, 熊静, 虞木奎
- CHENG Xiangrong, XING Wenli, YU Haijing, XIONG Jing, YU Mukui
- 披针叶茴香对变化光环境的表型可塑性
- Phenotypic plasticity of Illicium lanceolatum in response to varied light environments
- 生态学报. 2019, 39(6): 1935-1944
- Acta Ecologica Sinica. 2019, 39(6): 1935-1944
- http://dx.doi.org/10.5846/stxb201809192044
-
文章历史
- 收稿日期: 2018-09-19
- 修订日期: 2019-02-26

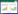




森林生态系统中地上(如光照)、地下资源(如养分)具有较大的时空变异性, 影响植物组成、生长和更新。光是影响森林中林下植物成活、生长和分布的最主要环境因素[1]。植物适应变化光环境的能力大小对其生存至关重要, 表型可塑性的大小是衡量植物对异质光环境适应能力的重要指标[2-3]。植物表型可塑性指植物对不同环境因子(如光照)变化做出响应而产生的不同特征表现[4], 是植物对异质生境的生态适应策略[5]。通过表型可塑性变化, 植物在异质生境中改变和调整各种形态或内在生理性状来适应环境, 增加对环境资源的获取能力, 以最大限度地利用资源, 从而降低环境对植物生长的负面影响[6-7]。Portsmut和Niinemets [8]将描述植物生长的相关性状归为生理性状, 而将光能捕获相关性状分为结构性状, 两者共同决定植物可塑性变化。据此, 将植物表型可塑性分为生理可塑性与结构可塑性。一些研究表明, 耐荫植物生理和结构可塑性低于非耐荫植物[9, 10], 因为高的可塑性可以让非耐荫植物获得较高的生长率。叶片性状与植物光合作用和光能利用直接相关, 是表征植物对弱光环境可塑性的关键表型。已有的多数研究集中在叶片形态和生理性状及其相对重要性如何适应变化光环境, 而对植物整体可塑性方面的研究相对较少, 这种整体可塑性决定了植物对地上、地下资源的利用, 其对植物在异质光环境中生存更为重要[11, 12]。近期的研究也发现, 随光照强度变化, 5种热带树种幼苗的叶片和细根面积(表面积)、氮、磷含量具有显著相关性[13]。
披针叶茴香(Illicium lanceolatum), 是八角科八角属的一种常绿小乔木, 广泛分布于我国南方丘陵山地。它是我国传统的药用植物, 具有活血止痛、祛风除湿的功效, 其所含的莽草酸可抑制动、静脉血栓及脑血栓形成, 也是一些抗病毒和抗癌药物的中间体[14]。披针叶茴香虽分布较广, 但多为零星散生, 随着野生资源的不断开发利用, 资源日渐稀少, 亟待加强种质资源保护和栽培[15]。披针叶茴香具有较强耐荫性, 光照条件显著影响其生长发育。基于叶绿素荧光动力学分析发现, 50%光照下的披针叶茴香光能利用效率接近100%光照处理, 而20%相对光照处理光能利用效率显著降低[16-17]。其他有关披针叶茴香的研究主要涉及其药用活性成分分析等方面[18, 19]。披针叶茴香光生态幅的大小, 以及对变化光环境的生态适应机制尚不清楚。因此, 系统开展不同光环境下披针叶茴香形态、生理和生物量分配等的研究, 有助于明确其在不同光环境下整体表型可塑性的变化, 揭示披针叶茴香对变化光环境的生态适应机制, 为今后披针叶茴香的高效栽培和利用提供参考。
1 材料与方法 1.1 试验地概况试验地位于浙江省杭州市富阳区高桥镇千家村(119°86′ E, 30°14′N)。试验区属亚热带季风气候, 年平均气温16.4 ℃, 年降水量1814 mm, 无霜期252 d, 年日照总时数1334.1 h。试验地地势平坦, 土壤为红黄壤, pH值为6.5。2002年之前为农地, 后改为试验苗圃。
1.2 试验方法在千家村平整田地上采用遮阳网模拟不同光照环境, 开展披针叶茴香对不同光环境表型可塑性的研究。选取150株生长良好的2年生披针叶茴香幼苗, 随机分成5组, 在3月中旬分别栽植于5个处理区内, 苗木间距大于50 cm。苗木生长1个月后, 开始进行光照控制实验, 采用遮阳网搭建遮荫棚设置4个光照处理, 光照强度分别为全光照的52%、33%、15%和6%, 另外设置1个自然光照处理(100%光照, 不遮荫)。光照强度采用光照计(TES 1339R, 中国台湾)在晴天中午测定5次, 然后取平均值。所有光照强度处理下生长的幼苗采用相同的抚育管理措施[20]。
1.3 采样和测定连续遮荫试验于11月中旬结束, 在各处理中随机选择6株幼苗整株收获, 用于形态和生物量等指标测定, 在剩余的植株中随机采集适量成熟叶片用于生理和细胞超微结构分析。整株收获的每个样株分叶、枝、干和根称取鲜重, 将每个光照处理下每个样株的叶片和根系采用WinFOLIA(Regent Company, Canada)分析系统测定叶片和根系形态特征, 包括叶长(leaf length, LL)、叶宽(leaf width, LW)、叶长/叶宽(LL/LW)、叶面积(leaf area, LA)、细根(fine root FR, φ≤2 mm)和粗根(Coarse root CR, φ>2 mm)的长度、表面积。然后每个样株叶、枝、干和根在105℃杀青1小时后在70℃下烘干至恒重, 分别计算单株叶、枝、干和根生物量。同时计算下列相关参数:根冠比(root biomass/aboveground biomass, R/A)=根生物量/地上部分生物量, 比叶面积(specific leaf area, SLA)=单株总叶面积/单株总叶重, 比根长(specific root length, SRL)=单株总根系长度/单株总根重, 比根表面积(specific root surface area, SRSA)=单株总根系表面积/单株总根重。取适量烘干的叶片和根系样品粉碎过2 mm筛, 进行全氮(total nitrogen, TN)和全磷(total phosphorus, TP)含量测定, TN含量用浓硫酸-过氧化氢消煮-半微量凯氏定氮法测定, TP含量用浓硫酸-过氧化氢消煮-钼锑抗比色法测定[21]。
叶片叶绿素a(chlorophyll a, Chl a)、叶绿素b(chlorophyll b, Chl b)和类胡萝卜素(carotenoids, Car)含量采用乙醇-丙酮比色法测定, 可溶性蛋白质(soluble protein, SP)采用考马斯亮蓝G-250染色法测定, 硝酸还原酶(nitrate reductase, NR)含量用分光光度法测定, 淀粉(starch, S)含量采用高氯酸水解-蒽酮比色法测量, 可溶性糖(soluble sugar, SS)含量采用蒽酮比色法测定, 超氧化物歧化酶(superoxide dismutase, SOD)活性的测定采用氮蓝四唑法, 硝酸盐(nitrate, NI)含量采用比色法进行测定[22]。
将处理后的披针叶茴香叶片采用2.5%戊二醛溶液与1%的锇酸溶液进行固定后, 用0.1 mol/L的磷酸缓冲液漂洗3次, 每次30 min。接着用30%、50%、70%、90%、100%的丙酮逐级脱水, 每次15 min。样品脱水后迅速移入丙酮、包埋剂中, 后按丙酮、包埋剂3:1、1:3混合液, 逐级渗透, 每级4 h, 最后移入纯包埋剂过夜渗透。采用瑞典LKB-V型超薄切片机进行切片, 切片经醋酸双氧铀和柠檬酸铅溶液分别染色, 最后用H-600型透射电子显微镜(日本日立)观察、拍照[23]。
1.4 数据处理采用Excel 2003和SPSS 22.0软件对数据进行统计分析。不同光照强度下披针叶茴香形态、生理和生物量等的差异采用SPSS 22.0软件的单因素方差分析(one way ANOVA)检验, 并进行Duncan多重比较法进行不同处理之间差异性检验分析(P < 0.05)。披针叶茴香功能性状的主成分分析(Principal Component Analysis, PCA)采用R软件包(R 3.5.1)完成。根据Valladares等[10]的方法计算某一指标的可塑性指数(Plasticity indices, PI):PI=(某指标在不同光强下的最大平均值-最小平均值)/最大平均值。表中数据为平均值±标准差。
2 结果与分析 2.1 不同光照强度下叶片性状变化对不同光照强度下披针叶茴香叶片形态特征分析表明, 遮荫处理增加了叶片平均长度, 弱光环境下(6%—15%相对光照处理)叶片平均宽度高于其他光照处理, 叶长/叶宽比值在不同光照条件下差异不显著(P>0.05), 平均单叶面积和比叶面积均随光照强度减弱呈增大趋势(表 1)。中等光照强度下(33%—52%相对光照处理), 单叶面积和比叶面积分别比全光照增加46.6%和18.9%;弱光环境下单叶面积和比叶面积分别比全光照增加76.9%和45.7%。尽管遮荫处理单株总叶面积比全光照增加了7.1%—24.8%, 但5种光照处理之间单株总叶面积没有显著差异(P>0.05)。叶片形态指标中单叶面积的可塑性指数(PI)较高, 其他指标PI均较低, 叶片形态指标的平均PI为0.34。
| 指标 Index | 处理 Treatment | 可塑性指数 PI Plasticity indices |
||||
| 100% | 52% | 33% | 15% | 6% | ||
| 叶长 Leaf length/g | 7.9±2.2b | 11.0±3.1a | 10.3±2.1a | 11.4±1.5a | 10.9±2.3a | 0.31 |
| 叶宽 Leaf width/g | 3.5±0.9b | 3.9±1.5b | 3.5±0.7b | 5.4±2.1a | 4.7±1.5ab | 0.35 |
| 叶长/叶宽 Leaf length/leaf width | 2.4±0.7 | 3.0±0.9 | 3.0±1.1 | 2.2±1.1 | 2.5±0.5 | 0.27 |
| 单叶面积 Leaf area/cm2 | 13.2±0.9c | 19.3±2.2b | 19.4±1.3b | 24.9±2.4a | 21.8±2.3ab | 0.47 |
| 总叶面积 Total leaf area/cm2 | 2623±406 | 3111±261 | 3273±408 | 2808±268 | 3143±474 | 0.31 |
| 比叶面积 Specific leaf area/cm2 | 65.9±5.3c | 77.3±4.9b | 79.4±8.2b | 96.2±7.8a | 95.8±9.6a | 0.31 |
叶绿素含量是衡量光合能力的重要指标, 遮荫处理的叶绿素a、叶绿素b和类胡萝卜素含量均比全光照处理有不同程度增加。极弱光照处理下(6%相对光照处理), 叶绿素a、叶绿素b、叶绿素a+b和类胡萝卜素含量最高, 叶绿素a/b比值最低。叶绿素和类胡萝卜素含量在15%、33%和52%相对光照处理之间没有显著差异(P>0.05), 但均显著高于全光照(100%)处理(P < 0.05)。
叶片非结构性碳水化合物可溶性糖含量随光照强度减弱显著降低, 6%—52%相对光照处理的淀粉含量均显著低于全光照处理, 但52%、33%、15%和6%相对光照处理之间淀粉含量差异不显著(P>0.05)(表 2)。叶片可溶性蛋白含量随光照强度的变化趋势与淀粉一致。6%—52%相对光照处理下叶片氮和磷含量均显著高于全光照处理, 叶氮含量随光照强度减弱呈增加趋势, 叶磷含量在52%相对光照处理下最高。6%—52%相对光照处理叶氮/叶磷比值与全光照处理之间没有显著差异(P>0.05), 52%相对光照处理下叶氮/叶磷比值显著低于33%、15%和6%相对光照处理(P < 0.05)。硝酸盐含量与硝酸还原酶活性的变化特征基本一致, 在33%和52%相对光照处理下, 硝酸盐含量与硝酸还原酶活性最低, 其次为15%相对光照处理, 在100%和6%相对光照处理下硝酸盐含量与硝酸还原酶活性最大。超氧化物岐化酶活性在不同光照处理之间差异不显著(P>0.05)。叶片生理指标中叶绿素a、叶绿素b, 叶绿素a+b、类胡萝卜素、可溶性糖、硝酸盐和硝酸还原酶活性的PI均大于0.5, 其他指标的PI相对较低, 叶片生理指标的平均PI为0.43。
| 指标 Index | 处理 Treatment | PI | ||||
| 100% | 52% | 33% | 15% | 6% | ||
| 叶绿素 a Chlorophyll a/(mg/g) | 0.88±0.18c | 1.27±0.09b | 1.17±0.01b | 1.24±0.22b | 1.83±0.05a | 0.52 |
| 叶绿素 b Chlorophyll b/(mg/g) | 0.17±0.04c | 0.27±0.01b | 0.26±0.01b | 0.29±0.05b | 0.46±0.02a | 0.63 |
| 叶绿素 a+b Chlorophyll a+b/(mg/g) | 1.05±0.21c | 1.53±0.10b | 1.44±0.01b | 1.53±0.27b | 2.29±0.07a | 0.54 |
| 叶绿素 a/b Chlorophyll a/ chlorophyll b | 5.18±0.71a | 4.76±0.37ab | 4.46±0.19bc | 4.25±0.09bc | 3.99±0.12c | 0.23 |
| 类胡萝卜素 Carotenoid/(mg/g) | 0.22±0.05c | 0.34±0.04b | 0.30±0.01b | 0.29±0.05b | 0.46±0.02a | 0.52 |
| 可溶性糖 Soluble sugar/(mg/g) | 95.36±13.53a | 55.72±8.16b | 58.66±3.68b | 28.47±5.67c | 29.80±3.10c | 0.70 |
| 淀粉 Starch/(mg/g) | 28.30±5.24a | 19.91±1.75b | 21.58±5.19b | 20.49±3.72b | 17.22±4.09b | 0.39 |
| 可溶性蛋白 Soluble protein/(mg/g) | 8.72±1.67a | 6.05±0.56b | 6.58±1.65b | 6.24±1.18b | 5.20±1.30b | 0.40 |
| 叶氮 Leaf nitrogen/(mg/g) | 13.20±0.57c | 15.40±0.14b | 17.05±0.21a | 16.15±0.35b | 17.15±0.07a | 0.23 |
| 叶磷 Leaf phosphorus/(mg/g) | 1.24±0.12d | 1.56±0.05a | 1.40±0.09bc | 1.34±0.02c | 1.45±0.06b | 0.21 |
| 叶氮/叶磷 Leaf nitrogen/leaf phosphorus | 10.76±1.51ab | 9.91±0.41b | 12.25±0.96a | 12.10±0.07a | 11.88±0.57a | 0.19 |
| 硝酸还原酶 Nitrate reductase/(μg g-1 h-1) | 21.53±3.37a | 10.27±0.67b | 10.33±1.35b | 16.32±5.22a | 20.87±9.64a | 0.52 |
| 硝酸盐 Nitrates/(mg/g) | 177.27±26.71a | 87.90±5.34c | 88.34±10.68c | 155.89±41.44b | 205.32±29.47a | 0.57 |
| 超氧化物岐化酶 Superoxide dismutase/(U/g) | 502.60±72.0a | 287.80±132.8a | 310.80±120.5a | 349.80±141.5a | 385.20±139.2a | 0.43 |
叶片细胞中叶绿体、细胞核等担负着重要的生理功能。不同光照强度处理披针叶茴香细胞超微结构观察结果如下:①在100%光照处理下(图 1A), 叶肉细胞内物质丰富, 细胞器结构完整, 膜系统完整, 叶绿体数量较多, 沿着细胞壁分布, 紧密排列成一圈, 呈椭圆形, 其上的黑色嗜锇颗粒数量很多, 白色的淀粉颗粒数量较少; ②在52%相对光照处理下(图 1B), 细胞器完整, 叶绿体沿着细胞壁紧密排列, 其上淀粉粒含量增多, 细胞核增大; ③在33%相对光照处理下(图 1C), 叶绿体沿着细胞壁紧密排列, 呈长梭形, 其上淀粉粒含量减少; ④在15%相对光照处理下(图 1D), 尽管叶绿体仍沿细胞壁紧密排列, 但叶绿体之间排列松散, 部分叶绿体体积增大, 其上白色淀粉粒含量进一步降低; ⑤在6%相对光照处理下(图 1E), 细胞内细胞器数量减少, 叶绿体排列紧密, 多成半圆形, 其上分布的嗜锇颗粒极少, 淀粉粒分布也较少, 细胞核减小。

|
| 图 1 不同光照强度下披针叶茴香叶片细胞超微结构 Fig. 1 Leaf cell ultrastructure of I. henryi in different light intensities (A), 100%光照处理; (B), 52%相对光照处理; (C), 33%相对光照处理; (D), 15%相对光照处理; (E), 6%相对光照处理; C, 叶绿体Chloroplast; N, 细胞核Nucleus; OG, 嗜锇颗粒Osmiophilic granules; SG, 淀粉粒Starch grain |
不同光照强度下根系性状的分析表明, 细根、粗根的长度和表面积以及比根长和比表面积在5种光照处理之间均没有显著差异(P>0.05)(表 3)。低光处理(15%和6%)根系氮含量显著降低(P < 0.05), 根系磷含量在52%相对光照处理下最高, 其他4种光照处理之间根系磷含量没有显著差异(P>0.05)。根系氮/磷比值在52%相对光照处理下最低, 其次为6%相对光照处理, 100%、33%和15%相对光照处理之间差异不显著(P>0.05)。根系性状中形态指标的PI(0.36)略高于根系养分含量PI(0.29), 根系形态和养分指标的平均PI为0.34。
| 指标 Index | 处理 Treatment | PI | ||||
| 100% | 52% | 33% | 15% | 6% | ||
| 细根长度 Fine root length/cm | 3356.4±576.3 | 3667.3±1968.2 | 4388.8±161.0 | 2589.0±1280.0 | 2791.2±726.5 | 0.41 |
| 粗根长度 Coarse root length/cm | 167.2±26.2 | 237.3±168.8 | 215.7±63.9 | 186.9±54.4 | 224.6±68.6 | 0.30 |
| 总根长 Total root length/cm | 3523.6±596.9 | 3904.6±2118.4 | 4604.5±1626.2 | 2776.0±1327.8 | 3015.8±792.9 | 0.40 |
| 细根表面积 Fine root surface area/cm2 | 661.2±90.5 | 699.4±395.0 | 794.5±301.6 | 456.7±234.6 | 498.5±141.5 | 0.43 |
| 粗根表面积 Coarse root surface area/cm2 | 182.9±90.5 | 290.9±200.5 | 267.4±80.6 | 259.1±52.6 | 296.5±92.8 | 0.38 |
| 总根表面积 Total root surface area/cm2 | 844.2±126.0 | 990.4±575.8 | 1062.0±313.0 | 715.8±273.3 | 795.0±231.1 | 0.33 |
| 比根长 Specific root length/(cm/g) | 106.9±27.6 | 87.6±38.8 | 133.4±46.2 | 89.9±43.3 | 92.8±28.9 | 0.34 |
| 比根表面积 Specific root surface area/(cm2/g) | 25.7±6.6 | 22.1±9.0 | 30.5±7.3 | 23.2±8.7 | 24.0±6.4 | 0.27 |
| 根系氮 Root nitrogen/(mg/g) | 8.22±0.21a | 8.31±0.37a | 8.52±0.55a | 7.35±0.07b | 6.89±0.09b | 0.19 |
| 根系磷 Root phosphorus/(mg/g) | 1.03±0.08b | 1.31±0.10a | 1.04±0.16b | 0.82±0.03b | 0.96±0.04b | 0.37 |
| 根系氮/磷 Root nitrogen/root phosphorus | 7.98±0.40ab | 6.34±0.21c | 8.26±0.72a | 8.93±0.39a | 7.20±0.18b | 0.29 |
不同光照条件下披针叶茴香单株各器官生物量的分析显示, 叶和根生物量在5种光照条件下均没有显著差异(P>0.05), 枝生物量在中等光照条件下(52%和33%)显著高于其他光照处理(P < 0.05), 干生物量随光强减弱呈增加趋势, 干生物量在6%—33%光照处理下显著高于100%和52%相对光照处理(P < 0.05)(表 4)。地上生物量和总生物量随光照条件的变化趋势基本一致, 6%—52%相对光照处理增加了单株地上生物量和总生物量, 仅52%和33%相对光照处理下生物量显著高于全光照(100%)。地上生物量/总生物量比值和根冠比在不同光照条件下差异不显著(P>0.05)。由此可见, 遮荫有利于披针叶茴香生物量积累, 尤其是中等光照条件下生长最佳。披针叶茴香单株各器官生物量指标中仅枝生物量PI为0.5, 其余指标PI均低于0.32, 生物量相关指标的平均PI为0.26。
| 指标 Index | 处理 Treatment | PI | ||||
| 100% | 52% | 33% | 15% | 6% | ||
| 叶生物量 Leaf biomass/g | 39.8±13.8 | 40.7±13.7 | 35.4±7.6 | 34.0±9.5 | 32.5±6.1 | 0.20 |
| 枝生物量 Branch biomass/g | 6.6±5.5b | 13.2±6.3a | 11.1±4.8a | 8.6±3.0b | 7.3±2.9b | 0.50 |
| 干生物量 Stem biomass/g | 35.3±11.2b | 42.5±9.7b | 48.2±10.3a | 48.2±4.1a | 50.0±10.3a | 0.29 |
| 根生物量 Root biomass/g | 32.1±9.4 | 46.0±21.7 | 35.7±11.2 | 31.1±5.3 | 32.5±10.1 | 0.32 |
| 地上生物量 Aboveground biomass/g | 81.7±29.7b | 96.3±25.5a | 94.7±21.4a | 90.9±7.8ab | 89.8±18.7ab | 0.15 |
| 总生物量 Total biomass/g | 113.8±30.1b | 142.3±39.3a | 130.3±29.8ab | 121.9±12.8ab | 122.3±27.5ab | 0.20 |
| 地上/总生物量 Aboveground biomass/total biomass |
0.72±0.15 | 0.68±0.24 | 0.73±0.08 | 0.75±0.03 | 0.73±0.08 | 0.09 |
| 根冠比 Root biomass/ aboveground biomass | 0.39±0.08 | 0.48±0.09 | 0.38±0.04 | 0.34±0.02 | 0.36±0.05 | 0.29 |
基于不同光照强度下披针叶茴香38个功能性状的主成分分析表明, 前2个主成分可以解释不同光照强度下表型变异总方差的78.5%, 其中PC1可以解释总方差的42.0%, 比叶面积、可溶性糖和叶绿素含量贡献较大; PC2可以解释总方差的36.5%, 总生物量、枝生物量和硝酸还原酶等的贡献较大(图 2)。100%光照强度下披针叶茴香功能性状与6%—52%相对光照处理处理差异较大, 52%和33%相对光照处理下性状随光照的变化特征接近, 15%和6%相对光照处理下性状随光照的变化特征接近。100%光照强度下可溶性糖、可溶性蛋白和超氧化物岐化酶等性状与其他光照处理之间差异较大, 52%和33%相对光照处理下根系形态学和生物量特征与其他光照处理之间差异较大, 15%和6%相对光照处理下比叶面积、硝酸盐等叶性状的差异较大。

|
| 图 2 不同光照强度下披针叶茴香功能性状的主成分分析(PCA) Fig. 2 Principal Component Analysis (PCA) of functional traits of I. henryi in different light treatments Ⅰ, 100%光照处理; Ⅱ, 52%相对光照处理; Ⅲ, 33%相对光照处理; Ⅳ, 15%相对光照处理; Ⅴ, 6%相对光照处理; LL, 叶长Leaf length; LW, 叶宽Leaf width; LL/LW, 叶长宽比Leaf length/leaf width; LA, 叶面积Leaf area; TLA, 总叶面积Total leaf area; SLA, 比叶面积Specific leaf area; Chla, 叶绿素a Chlorophyll a; Chlb, 叶绿素b Chlorophyll b; Chla+b, 叶绿素a+b Chlorophyll a + chlorophyll b; Chla/b, 叶绿素a/b Chlorophyll a/chlorophyll b; Car, 类胡箩卜素Carotenoid; SS, 可溶性糖Soluble sugar; S, 淀粉Starch; SP, 可溶性蛋白Soluble protein; LN, 叶氮Leaf nitrogen; LP, 叶磷Leaf phosphorus; NR, 硝酸还原酶Nitrate reductase; NI, 硝酸盐Nitrates; SOD, 超氧化物岐化酶Superoxide dismutase; FRL, 细根长Fine root length; CRL, 粗根长Coarse root length; TRL, 总根长Total root length; FRSA, 细根表面积Fine root surface area; CRSA, 粗根表面积Coarse root surface area; TRSA, 总根系表面积Total root surface area; SRL, 比根长Specific root length; SRSA, 比根表面积Specific root surface area; RN, 根系氮Root nitrogen; RP, 根系磷Root phosphorus; RN/RP, 根系氮/磷Root nitrogen/root phosphorus; LB, 叶生物量Leaf biomass; BB, 枝生物量Branch biomass; SB, 干生物量Stem biomass; RB, 根生物量Root biomass; AB, 地上生物量Aboveground biomass; TB, 总生物量Total biomass; AB/TB, 地上生物量/总生物量Aboveground biomass/total biomass; R/A, 根冠比Root biomass/aboveground biomass |
叶片是植物进行光合作用及与外界环境进行物质和能量交换的主要场所, 其形态结构和生理功能与生长环境密切相适应, 是植物对外界环境变化最敏感的器官[24], 因而, 植物对环境变化的多数响应主要体现在叶性状上[10]。研究表明, 随光照有效性降低, 植物会通过增大叶长和叶宽, 来增加光合作用的叶面积, 进而增加对光能的捕获能力[25, 26]。本研究也发现随光照强度减弱, 叶长和单叶面积均有一定程度增加。比叶面积能够反映植物对资源获取和利用的权衡关系[12]。通常, 在低光环境中, 植物分配更多的生物量给叶片来增加受光面积, 一般具有较高的比叶面积; 在高光环境中, 植物为减少蒸腾、增加碳的固定, 形成小而厚的叶片, 比叶面积较低[27]。本研究披针叶茴香比叶面积随光照有效性降低显著增加, 较大的比叶面积表明植物在低光环境中具有较高的光能捕获效率。其他一些研究表明, 尽管比叶面积随光强减弱显著增大, 但植物总叶面积显著减小, 导致整株的受光面积大大减少, 这将影响其光合产物的积累[28]。本研究中总叶面积随光照强度减弱并没有发生显著变化, 这与叶生物量与比叶面积的变化有关。随光照强度减弱, 叶生物量呈减少趋势, 而比叶面积逐渐增大, 因而总叶面积在不同光照处理之间的变异较小。总体上, 披针叶茴香通过比叶面积大小来适应变化光环境, 其叶片形态的表型可塑性相对较低。
植物的光合作用主要依靠叶绿素和光合酶来捕获和利用光能, 合成光合产物。在低光环境中, 植物为有效利用光能, 通常叶绿素含量增加, 叶绿素比例发生变化[24]。叶绿素a含量大小可反映叶片光能利用能力, 而总叶绿素含量尤其是叶绿素b与光能捕获能力密切相关。随光照有效性降低, 植物为了更有效地捕获光, 叶绿素含量尤其是叶绿素b的含量增加, 同时叶绿素a/b下降[29, 30]。本研究也得出类似结论。对山栀子(Gardenia jasminoides)幼苗遮荫试验发现, 叶绿素b含量和叶绿素a/b在6%—100%光照环境中没有显著差异[28]; 而麻栎(Quercus acutissima)幼苗在中等光照条件下(66%相对光照)叶绿素a和叶绿素b的含量与全光照处理之间没有显著差异, 低光环境(10%相对光照)叶绿素a和叶绿素b含量显著高于全光照, 叶绿素a/b在3种光照处理之间没有显著变化[31]。这些不同的研究结果可能与树种差异及其对光环境的不同适应机制有关。本研究中披针叶茴香的地上生物量和总生物量在极弱光照下也没有显著降低, 这可能主要与叶绿素和类胡萝卜素含量的大幅增加, 进而提高了其对光的捕获和利用能力有关(表 2)。
不同的光照强度影响叶片的碳氮代谢过程。在高光环境下, 光合产物和可溶性蛋白含量较多, 硝酸还原酶活性较高[32, 33]。而生长在弱光环境中的植物分配较多的碳给叶片, 以获取更多的光能, 但叶片中光合产物积累减少, 硝酸还原酶活性降低, 导致叶片中硝酸盐的积累[32, 33]。本研究中遮荫处理的可溶性糖、淀粉和可溶性蛋白含量均显著低于全光照处理, 表明遮荫后光合产物的积累减少, 尤其是可溶性糖含量随光照强度减弱显著降低。6%—52%相对光照处理下叶氮含量显著增加, 在33%和52%相对光照处理下, 硝酸盐积累较少; 而在全光照(100%)和极弱光照条件下(6%相对光照), 尽管硝酸还原酶活性较高, 但硝酸盐积累也较多, 这与已有的一些研究结果不同。这表明在不利光照条件下, 披针叶茴香为缓解硝酸盐积累对生长的胁迫, 相应的增加硝酸还原酶活性来减少这一负面影响。遮荫也增加了叶磷含量, 但对叶氮/叶磷比值影响相对较小, 表明叶氮和叶磷随光照强度变化具有协同效应, 叶片氮磷化学计量比几乎不受光照强度变化的影响。
在适宜条件下, 植物叶片细胞内叶绿体呈纺锤形结构, 细胞器结构完整, 基粒片层发达、清晰可见, 排列整齐有序, 没有异常的细胞内含体或空泡化结构[34]。本研究5种光照强度下细胞结构基本保持完整, 52%相对光照处理下叶绿体内淀粉含量最高, 低光下细胞内叶绿体体积较全光照处理下增大, 这些结果表明披针叶茴香叶片细胞结构在所有光照处理下保持正常的生理代谢, 其对变化光环境的适应性较强。而对其他一些植物的研究发现, 低光下植物生理代谢活动降低, 细胞受损、细胞器破坏严重, 进而导致光合作用效率下降[35]。
3.2 根系性状对变化光环境的表型可塑性光环境的变化不仅影响植物叶片性状, 也可能影响植物根系性状及其对养分资源的利用。Pooter等[27]指出, 在低光环境, 植物具有较高的叶面积比来获取光照, 而在高光环境下, 较高的根生物量比更有利于水分和养分的获取。在土壤养分有效性较低时, 植物通过增加比根长和比根表面积来吸收更多养分, 而养分有效性较高时, 植物增加比叶面积和叶面积比获得更多光照来促进生长。本研究中披针叶茴香细根、粗根的形态特征在5种光照处理之间均没有显著差异, 表明短期遮荫处理对根系生长发育的影响较小。尽管如此, 低光处理(15%和6%)下, 根系氮含量显著降低, 这可能与低光下植物氮需求减少(如叶片氮积累较多)有关。在52%相对光照处理下, 根系磷利用效率最高, 根系氮/磷比值最低, 因此, 适宜的光照环境可能有利于磷的利用。Gong等[13]也发现, 5种热带树种幼苗在4%和20%相对光照处理之间细根的比根长和比叶面积没有显著差异, 但这些树种的叶面积和细根表面积呈显著的正相关, 叶片和细根的氮磷化学计量比具有显著相关性。本研究披针叶茴香叶片和根系形态及生态化学计量特征均没有显著相关性, 表明随光照强度变化, 披针叶茴香对地下资源的获取受地上资源利用的影响较小。
3.3 整体结构的表型可塑性当植物受到光照条件限制时, 植物分配更多的资源供给地上部分, 增加叶、茎生物量的分配来获得更多的光照资源[36]。本研究发现6%—52%相对光照处理有利于披针叶茴香单株总生物量的积累, 尤其是52%相对光照处理总生物量最大。随光照强度减弱, 披针叶茴香分配给叶片和根系的生物量比例没有显著变化, 但分配给枝条和树干的生物量比例逐渐增加, 这表明在低光条件下, 植物采取保守策略, 光合同化产物主要用于枝、干生物量的积累。对欧洲山毛榉(Fagus sylvatica)[37]、闽楠(Phoebe bournei)[38]幼树的研究也发现, 叶片或地上生物量比、根系生物量比在不同光照处理之间没有显著差异。一些研究表明, 低光环境下植物为了存活尽量减少呼吸损失[39], 通过可塑性适应环境的改变, 增加碳在枝干和根系的储存, 而不是为了获取资源[40]。一般具有较宽生态幅和拓殖能力的物种拥有较高生物量分配可塑性, 由此缓解不同生境的选择压力[41]。但披针叶茴香生物量及其分配特征的可塑性较低, 说明其并不依赖生物量分配策略来适应变化光环境。相对于叶片形态和生物量可塑性, 叶片结构和生理可塑性对披针叶茴香适应变化光环境发挥了更为重要的作用。本研究中披针叶茴香比叶面积、光合色素和叶氮含量随光照强度减弱逐渐增大, 均有利于披针叶茴香提高光能的捕获和利用效率, 进而维持其正常的生理代谢。披针叶茴香具有较宽的光生态幅, 在中等光照条件(52%相对光照处理)下生长最佳, 而在极弱光照条件下(6%相对光照处理)叶绿素含量增加幅度远大于高光环境中(100%光照)叶绿素含量降低幅度, 尤其是叶绿素b的变异水平更高, 表明低光环境中植物为获取更多的光照资源投入更大。披针叶茴香幼苗整体结构的可塑性相对较低, 叶片生理的可塑性对变化光环境适应可能发挥了更加重要的作用。
4 结论本文研究了披针叶茴香在5种光照强度下叶片、根系和生物量的表型可塑性, 初步明确了披针叶茴香对变化光环境的响应和适应策略。披针叶茴香具有较宽的光生态幅, 其在6%—100%光照强度下均可以正常生长, 荫庇环境有助于披针叶茴香地上生物量和总生物量积累, 在52%相对光照处理下生物量最大。披针叶茴香叶片和根系形态、生物量及氮磷化学计量特征在6%—100%光照强度下可塑性较低, 叶片生理指标(尤其是光合色素和可溶性糖含量)的调节在披针叶茴香适应不同光环境过程中发挥了关键作用。这些结论仅为一个生长季的结果, 不同光照环境下披针叶茴香长期适应机制还有待进一步研究。
| [1] |
Aleric K M, Kirkman L K. Growth and photosynthetic responses of the federally endangered shrub, Lindera Melissifolia (Lauraceae), to varied light environments. American Journal of Botany, 2005, 92(4): 682-689. DOI:10.3732/ajb.92.4.682 |
| [2] |
郭志华, 王荣, 肖文发. 不同光环境下喜树与四川大头茶幼苗的表型可塑性. 林业科学, 2009, 45(9): 6-12. DOI:10.3321/j.issn:1001-7488.2009.09.002 |
| [3] |
刘从, 田甜, 李珊, 王芳, 梁宇. 中国木本植物幼苗生长对光照强度的响应. 生态学报, 2018, 38(2): 518-527. |
| [4] |
Gao L, Li B, Liu W Y, Shen Y X, Liu W J. Inhibition effects of daughter ramets on parent of clonal plant Eichhornia crassipes. Aquatic Botany, 2013, 107: 47-53. DOI:10.1016/j.aquabot.2013.01.010 |
| [5] |
Bradshaw A D. Evolutionary significance of phenotypic plasticity in plants. Advances in Genetics, 1965, 13: 115-155. DOI:10.1016/S0065-2660(08)60048-6 |
| [6] |
施建敏, 叶学华, 陈伏生, 杨清培, 黎祖尧, 方楷, 杨光耀. 竹类植物对异质生境的适应——表型可塑性. 生态学报, 2014, 34(20): 5687-5695. |
| [7] |
胡启鹏, 郭志华, 李春燕, 马履一. 植物表型可塑性对非生物环境因子的响应研究进展. 林业科学, 2008, 44(5): 135-142. DOI:10.3321/j.issn:1001-7488.2008.05.025 |
| [8] |
Portsmuth A, Ni+inemets V. Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Functional Ecology, 2007, 21(1): 61-77. |
| [9] |
Sánchez-Gómez D, Valladares F, Zavala M A. Functional traits and plasticity in response to light in seedlings of four Iberian forest tree species. Tree Physiology, 2006, 26(11): 1425-1433. DOI:10.1093/treephys/26.11.1425 |
| [10] |
Valladares F, Wright S J, Lasso E, Kitajima K, Pearcy P W. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest. Ecology, 2000, 81(7): 1925-1936. DOI:10.1890/0012-9658(2000)081[1925:PPRTLO]2.0.CO;2 |
| [11] |
Valladares F, Sanchez-Gomez D, Zavala M A. Quantitative estimation of phenotypic plasticity:bridging the gap between the evolutionary concept and its ecological applications. Journal of Ecology, 2006, 94(6): 1103-1116. DOI:10.1111/jec.2006.94.issue-6 |
| [12] |
Gratani L. Plant phenotypic plasticity in response to environmental factors. Advances in Botany, 2014, 2014: 208747. |
| [13] |
Gong H D, Wang H, Jiao D Y, Cai Z Q. Phenotypic plasticity of seedlings of five tropical tree species in response to different light and nutrient availability. Tropical Ecology, 2016, 57(4): 727-737. |
| [14] |
郑成忠, 徐金良, 郑卫华. 优良林下药用观赏树种——披针叶茴香. 林业调查规划, 2014, 39(1): 98-100. DOI:10.3969/j.issn.1671-3168.2014.01.023 |
| [15] |
曹永慧, 周本智, 陈双林, 萧江华, 王小明. 不同产地披针叶茴香光合特性对水分胁迫和复水的响应. 生态学报, 2012, 32(23): 7421-7429. |
| [16] |
Cao Y H, Zhou B Z, Chen S L, Xiao J H, Wang X M. The photosynthetic physiological properties of Illicium lanceolatum plants growing under different light intensity conditions. African Journal of Agricultural Research, 2011, 6(26): 5736-5741. |
| [17] |
曹永慧, 周本智, 张汝民, 陈双林. 披针叶茴香叶绿素荧光参数对不同光环境的响应. 西北植物学报, 2012, 32(3): 525-531. DOI:10.3969/j.issn.1000-4025.2012.03.014 |
| [18] |
李慧娟, 王丽霞, 刘孟奇, 张雁冰, 毕跃峰, 柳继锋. 披针叶茴香果实的化学成分研究. 中国药学杂志, 2014, 49(8): 636-639. |
| [19] |
王国伟.披针叶茴香茎、叶化学成分及抗炎活性研究[D].上海: 第二军医大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-90024-1012404469.htm
|
| [20] |
颉洪涛, 虞木奎, 成向荣. 光照强度变化对5种耐阴植物氮磷养分含量、分配以及限制状况的影响. 植物生态学报, 2017, 41(5): 559-569. |
| [21] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2013.
|
| [22] |
张治安, 张美善, 蔚荣海. 植物生理学实验指导. 北京: 中国农业科学技术出版社, 2004.
|
| [23] |
吴月燕, 李波, 张燕忠, 崔鹏. 盐胁迫对杜鹃生理生化与叶绿体亚显微结构的影响. 浙江大学学报:农业与生命科学版, 2011, 37(6): 642-648. |
| [24] |
王艺, 韦小丽. 不同光照对植物生长、生理生化和形态结构影响的研究进展. 山地农业生物学报, 2010, 29(4): 353-359, 370-370. DOI:10.3969/j.issn.1008-0457.2010.04.017 |
| [25] |
徐高峰, 申时才, 张付斗. 异质环境下入侵植物薇甘菊的适应性与繁殖特性. 生态环境学报, 2014, 23(8): 1258-1264. DOI:10.3969/j.issn.1674-5906.2014.08.002 |
| [26] |
孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150-165. DOI:10.3321/j.issn:1005-264X.2007.01.019 |
| [27] |
Poorter L. Growth responses of 15 rain-forest tree species to a light gradient:the relative importance of morphological and physiological traits. Functional Ecology, 1999, 13(3): 396-410. DOI:10.1046/j.1365-2435.1999.00332.x |
| [28] |
成向荣, 周俊宏, 陈永辉, 武克壮, 虞木奎. 山栀子幼苗表型可塑性对不同光环境的响应. 江西农业大学学报, 2016, 38(1): 180-186. |
| [29] |
李冬林, 向其柏. 光照条件对浙江楠幼苗生长及光合特性的影响. 南京林业大学学报:自然科学版, 2004, 28(5): 27-31. |
| [30] |
刘文海, 高东升, 束怀瑞. 不同光强处理对设施桃树光合及荧光特性的影响. 中国农业科学, 2006, 39(10): 2069-2075. DOI:10.3321/j.issn:0578-1752.2006.10.018 |
| [31] |
徐飞, 郭卫华, 徐伟红, 王仁卿. 不同光环境对麻栎和刺槐幼苗生长和光合特征的影响. 生态学报, 2010, 30(12): 3098-3107. |
| [32] |
关义新, 林葆, 凌碧莹. 光、氮及其互作对玉米幼苗叶片光合和碳、氮代谢的影响. 作物学报, 2000, 26(6): 806-812. DOI:10.3321/j.issn:0496-3490.2000.06.026 |
| [33] |
Vos J, van der Putten P E L. Effects of partial shading of the potato plant on photosynthesis of treated leaves, leaf area expansion and allocation of nitrogen and dry matter in component plant parts. European Journal of Agronomy, 2001, 14(3): 209-220. DOI:10.1016/S1161-0301(00)00090-3 |
| [34] |
林多, 黄丹枫, 杨延杰, 陈宁. 钾素水平对网纹甜瓜叶片光合特性及叶绿体亚显微结构的影响. 应用生态学报, 2007, 18(5): 1066-1070. DOI:10.3321/j.issn:1001-9332.2007.05.021 |
| [35] |
陶巧静, 吴月燕, 付涛, 项锡娜, 李波. 弱光胁迫对西洋杜鹃生理特性和叶片超微结构的影响. 林业科学, 2015, 51(3): 84-92. |
| [36] |
Nishimura E, Suzaki E, Irie M, Nagashima H, Hirose T. Architecture and growth of an annual plant Chenopodium album in different light climates. Ecological Research, 2010, 25(2): 383-393. DOI:10.1007/s11284-009-0666-6 |
| [37] |
Curt T, Coll L, Prévosto B, Balandier P, Kunstler G. Plasticity in growth, biomass allocation and root morphology in beech seedlings as induced by irradiance and herbaceous competition. Annals of Forest Science, 2005, 62(1): 51-60. DOI:10.1051/forest:2004092 |
| [38] |
王振兴, 朱锦懋, 王健, 汪滢, 卢钰茜, 郑群瑞. 闽楠幼树光合特性及生物量分配对光环境的响应. 生态学报, 2012, 32(12): 3841-3848. |
| [39] |
Craine J M, Reich P B. Leaf-level light compensation points in shade-tolerant woody seedlings. New Phytologist, 2005, 166(3): 710-713. DOI:10.1111/nph.2005.166.issue-3 |
| [40] |
Reich P B, Wright I J, Cavender-Bares J, Craine J M, Oleksyn J, Westoby M, Walters M B. The evolution of plant functional variation:traits, spectra, and strategies. International Journal of Plant Sciences, 2003, 164(S3): S143-S164. DOI:10.1086/374368 |
| [41] |
武高林, 陈敏, 杜国祯. 三种高寒植物幼苗生物量分配及性状特征对光照和养分的响应. 生态学报, 2010, 30(1): 60-66. DOI:10.3969/j.issn.1674-764x.2010.01.008 |