 2019, Vol. 39
2019, Vol. 39文章信息
- 何学敏, 吕光辉, 秦璐, 李岩, 杨晓东, 杨建军, 于恩涛
- HE Xuemin, LÜ Guanghui, QIN Lu, LI Yan, YANG Xiaodong, YANG Jianjun, YU Entao
- 荒漠杜加依林冠层水热变化及CO2交换特征
- Carbon interfacial hydrothermal variation and CO2 exchange characteristics of the Tugai forest in a desert environment
- 生态学报. 2019, 39(3): 1052-1062
- Acta Ecologica Sinica. 2019, 39(3): 1052-1062
- http://dx.doi.org/10.5846/stxb201712052188
-
文章历史
- 收稿日期: 2017-12-05
- 网络出版日期: 2018-11-15

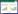




2. 新疆大学生态学博士后科研流动站, 乌鲁木齐 830046;
3. 新疆绿洲生态教育部重点实验室, 乌鲁木齐 830046;
4. 新疆环境保护科学研究院, 乌鲁木齐 830011;
5. 新疆环境污染监控与风险预警重点实验室, 乌鲁木齐 830011;
6. 新疆大学资源与环境科学学院, 乌鲁木齐 830046;
7. 中国科学院大气物理研究所, 北京 100029
2. Post-Doctoral Research Center for Ecology, Xinjiang University, Urumqi 830046, China;
3. Xinjiang Key Laboratory of Oasis Ecology, Urumqi 830046, China;
4. Xinjiang Academy of Environment Protection Science, Urumqi 830011, China;
5. Xinjiang Key Laboratory of Environmental Pollution Monitoring and Risk Warning, Urumqi 830011, China;
6. College of Resources and Environment Science, Xinjiang University, Urumqi 830046, China;
7. Institute of Atmospheric Physics, Chinese Academy of Sciences, Beijing 100029, China
森林生态系统存储了陆地植被80%的碳, 在调控大气CO2浓度和稳定全球气候方面扮演主导角色[1]。在陆地生态系统中, 森林冠层是由植物的枝、叶之间聚合和组合而构成的整体, 是植物与外界环境进行相互作用中最直接和最活跃的界面, 冠层控制着植物的多种生物物理和生理过程, 如光合、呼吸、降水截留等, 对生态系统的能量流动、生物多样性以及气候变化等均具有重要影响[2-3]。研究冠层内气象要素的时空变化规律, 是准确模拟地气系统物质能量交换过程的重要前提[4]。同时, 开展森林表面大气湍流结构和空气动力学特征等研究, 对于深入了解森林冠层物质和能量交换过程具有重要的现实意义。
高大的森林冠层内, 微气象要素的垂直变化规律比大气中复杂的多, 直接影响了植物-大气界面的物质循环和能量传输过程, 同时亦显著影响大气的层结状况。前人对森林冠层的研究发现, 温带混交林冠层内温度比其上空气温度高4.5—5.0℃[5], 林冠内的温度日较差低于冠外, 而湿度明显高于冠外[6-7], 在干旱区同样发现温度、湿度及风速等环境因子在垂直高度和林冠上存在明显的垂直变化特征[8-9]。植物冠层结构与组成对干旱荒漠生态系统地表水热和碳收支具有重要的影响[10-12], 目前, 关于森林冠层微气象与地表水热和碳收支相互作用的研究鲜见, 仍然是生态学和气象学等交叉学科领域研究的重点和难点。
艾比湖流域植物区系属古北界蒙新区北疆荒漠亚区准噶尔荒漠小区, 在湖滨区和补给河流沿岸分布着大量由荒漠植物组成的天然植物群落—杜加依林(Tugai forest)[13], 该群落是由干旱荒漠中沿河岸分布的乔木、灌木和草本组成[14], 不仅是荒漠地区动植物重要栖息地和生物多样性的维持地, 同时在防风固沙、水源涵养和河岸防护等方面具有重要作用[15-16]。对于艾比湖流域内荒漠杜加依林而言, 此类生态系统往往对全球气候变化具有较高的敏感性, 同时在干旱区碳循环中起着重要的作用[17-18]。因此, 本研究利用通量观测塔连续观测艾比湖流域内荒漠杜加依林数据, 分析CO2通量典型日变化与季节变化规律, 同时比较林冠层界面湍流碳通量、植物冠层储存碳通量以及CO2净交换量(NEE), 在此基础上探讨冠层界面温湿廓线与CO2交换过程的相互关系, 以期探索荒漠杜加依林不同时间和垂直空间序列下温湿变化对碳收支的影响, 揭示复杂下垫面模式中杜加依林冠层界面物质和能量交换规律。
1 材料与方法 1.1 研究区概括研究区位于我国新疆博尔塔拉蒙古自治州艾比湖湿地国家级自然保护区内(图 1), 地理位置为82°30′47″—83°50′21″E, 44°37′05″—45°10′35″N, 属北温带大陆性干旱气候, 夏季高温干旱, 冬季寒冷。艾比湖通量观测塔(2012XJ-AiBiHu-OPEC)架设于保护区内的东大桥管护站处, 于2012年1月调试后正式运行。监测对象为流域内荒漠杜加依林, 植被群落类型以胡杨(Populus euphratica Oliv.)和芦苇(Phragmites australis (Cav.) Trin. ex Steud)为优势种, 伴生种有琵琶柴(Reaumuria soongonica (PalL) Maxim)、盐豆木(Halimodendron halodendron (Pall.) Voss. var. halodendron)、黑果枸杞(Lycium ruthenicum Murr.)、罗布麻(Apocynum venetum Linn.)、光果甘草(Radix glycyrrhiza glabra)以及猪毛菜属等。森林结构为矮乔木-灌木-草本, 多呈稀疏型分布特征, 其中, 矮乔木平均高度约6.61 m, 平均冠幅为24.76 m2, 灌木和草本散布在乔木林间, 灌木平均高度为0.65 m, 平均冠幅为1.56 m2。

|
| 图 1 研究区示意图 Fig. 1 Study samples schemes |
利用艾比湖通量观测塔配套仪器(表 1)获取研究区2012年1—12月的碳通量及相关数据, 涡度相关系统采样频率为10 Hz, 气象与土壤梯度系统数据采集周期为10 min。
| 序号 Number |
仪器名称 Instrument name |
仪器型号 Instrument model |
厂商与国家 Manufacturer and country |
观测项目 Observation project |
观测高度 Observed altitude |
| 1 | 开路涡动相关系统 | CSAT3, EC150 | Campbell, USA | CO2、感热、潜热通量等 | 15 m |
| 2 | 空气温/湿度传感器 | HMP155A | Vaisala, FIN | 空气温度、湿度 | 2, 3, 6, 12, 15, 21, 27 m |
| 3 | 风速传感器 | 010C | Met One, USA | 风速 | 2, 3, 6, 12, 15, 21, 27 m |
| 4 | 风向传感器 | 020C | Met One, USA | 风向 | 27 m |
| 5 | 数据采集装置 | CR3000 | Campbell, USA | 存储数据 | 1.5 m |
对于获取的湍流通量数据, 分别进行频率响应校正、WPL校正, 在此基础上进行质量控制与评价, 剔除因传感器故障或异常天气等原因产生的不合理“野点”。部分异常或缺失的梯度数据, 使用通量观测塔旁架设的气象站同步观测数据替代。此外, 本文数据分析时间均采用地方时。
1.3 数据质量评价与插补参考我国通量数据处理标准对站点观测数据进行坐标轴旋转、WPL校正、存储项计算以及数据质量评价, 去除地形、水热、植物冠层等因素的影响[19], 处理后总缺失数据比例达45.58%, 采用非线性回归法分别对缺失数据进行插补[20-21]。
1.4 主要计算公式依据Monin-Obukhov相似理论, 利用超声风速仪观测数据, 考虑粗糙度和零平面位移的影响, Monin-Obukhov稳定度参数计算公式如下:
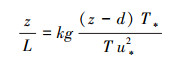
|
(1) |
式中, z为观测高度;L表示湍流混合的垂直尺度;d为零平面位移;T为气温;u*为摩擦风速;T*表示湍流尺度温度;k为Karman常数;g为重力加速度。
NEE采用以下计算公式, 其中冠层储存项采用单点浓度变化估算[22-23], 即:
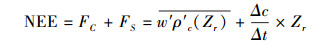
|
(2) |
式中, NEE为生态系统净碳交换量;FC为大气和生态系统界面的湍流通量;FS为冠层内的碳储存通量;w′为垂直风速的脉动量;ρ′c为CO2密度的脉动量;Zr为通量观测高度;Δc为前、后两次相邻时刻测定的CO2浓度差;Δt为前、后两次测定的时间间隔。
2 结果与分析 2.1 荒漠杜加依林冠层界面大气稳定状况艾比湖流域内荒漠杜加依林冠层界面上的摩擦风速(μ*)从1月到12月表现出先增加后减小的趋势(表 2), 1月最小, 为0.125 m/s, 最大值出现在6月, 为0.353 m/s, 杜加依林冠层全年平均μ*为0.266 m/s。湍流动能(TKE)与摩擦风速变化趋势相同, 亦随月份表现出先增后减的趋势, 年平均TKE为0.762 m2/s2。
| 月份Month | μ*/(m/s) | TKE/(m2/s2) | z/L | |
| 1 | 0.125 | 0.178 | -0.337 | |
| 2 | 0.196 | 0.448 | -0.269 | |
| 3 | 0.244 | 0.640 | -0.185 | |
| 4 | 0.338 | 1.168 | -0.131 | |
| 5 | 0.331 | 1.091 | -0.168 | |
| 6 | 0.353 | 1.129 | -0.059 | |
| 7 | 0.289 | 0.889 | 0.030 | |
| 8 | 0.319 | 1.028 | 0.020 | |
| 9 | 0.250 | 0.674 | 0.045 | |
| 10 | 0.237 | 0.608 | 0.352 | |
| 11 | 0.307 | 0.881 | -0.019 | |
| 12 | 0.202 | 0.413 | -1.605 | |
| 平均Average | 0.266 | 0.762 | -0.194 | |
| μ*:摩擦风速, Friction velocity;TKE:湍流动能, Turbulent kinetic energy;z/L:大气稳定度参数, Monin-Obukhov atmospheric stability | ||||
参考大气稳定度分类标准[24], 规定z/L < -0.1为不稳定大气, z/L > 0.1时为稳定大气, -0.1≤z/L≤0.1之间为中性大气。由表 2可以看出, 1—5月以及12月流域内荒漠杜加依林近地层大气表现为不稳定状态(z/L < -0.1), 6—9月大气稳定度逐渐转为中性状态, 10月z/L达到0.352, 为2012年仅有的大气稳定状态, 同时全年z/L均值为-0.194, 表明就全年水平而言, 艾比湖流域荒漠杜加依林大气稳定度总体呈不稳定水平。
2.2 荒漠河岸林冠层界面水热变化特征水热因子作为影响干旱区物质和能量交换的最重要因素, 其交互作用对荒漠杜加依林CO2通量的变化影响重大。由近地层0—30 m温度廓线变化情况(图 2)可以看出, 1—12月近地层气温随着高度的升高均表现为递增趋势, 除10月温度变幅超过4℃以外, 其他月份均变幅均小于2—3℃, 特别是5、6、7月垂直温度变幅在2℃以内, 此时期荒漠杜加依林与大气界面间热传输较弱, 大气层结较为稳定。

|
| 图 2 2012年荒漠杜加依林冠层温度廓线特征 Fig. 2 Characteristics of canopy interface temperature profiles of Tugai forest in desert during 2012 |
荒漠杜加依林空气相对湿度廓线总体呈秋冬湿春夏干的特点(图 3)。冬季空气相对湿度较高, 其中1月达到全年最大值为78.17%(27 m处), 冬季到春季空气相对湿度迅速下降, 在5月达到全年最低值为34.03%(27 m处), 此时天气特征多为大风沙尘天气。伴随土壤冻融以及植物蒸散发, 空气相对湿度逐渐回升, 在整个生长季多维持在40%—50%之间, 到秋冬季逐渐回归到高值水平。

|
| 图 3 2012年荒漠杜加依林冠层湿度廓线特征 Fig. 3 Characteristics of canopy humidity profiles of Tugai forest in desert during 2012 |
将2012年按新疆气候特征划分为冬(12—翌年2月)、春(3—5月)、夏(6—8月)、秋(9—11月)4个季节[25], 分别选取天气状况良好、通量信号强度大于0.85且无通量数据缺失的典型日2月21日、4月22日、8月21日和10月21日进行不同季节典型日内CO2浓度和通量日变化特征分析, 如图 4所示:

|
| 图 4 荒漠杜加依林冠层CO2浓度和碳通量典型日变化特征 Fig. 4 The typical diurnal variation characteristic of canopy interface carbon dioxide and carbon flux for Tugai forest in desert |
夏季典型日荒漠杜加依林CO2浓度变化范围在633.316—762.660 mg/m3, 对应碳通量为-0.807—0.587 mg m-2 s-1, 总体呈“U”型, 远大于稀疏干旱植被的-0.023 mg m-2 s-1[26]。夏季杜加依林夜间多有微风, 此时碳通量多以正值为主且呈波动趋势, 昼间碳通量为负且呈不规则“波谷”, 9:00左右为正负值交替的时段, 日平均碳通量为-0.102 mg m-2 s-1。研究区秋季昼夜温差较大, 大气CO2浓度变化范围为707.624—760.023 mg/m3, 对应碳通量为-0.218—0.141 mg m-2 s-1。夜间通量波动强烈且正值居多, 昼间则表现为较小的碳吸收, 碳通量平均值约为0.008 mg m-2 s-1。
从全年水平来看(图 5), 流域内荒漠杜加依林全年CO2浓度阈值为594.800—894.101 mg/m3, 整体表现为平缓的“U型”变化趋势, 1月和11月在一年中具有较高值, 其中1月略高于11月, 7月达到全年最低谷值。湍流通量阈值为-0.235—0.196 mg m-2 s-1, 1月初至3月中下旬, 以及10月初到12月底均表现为在0值附近波动, 4月和5月表现出CO2湍流通量的较高峰值, 为冻融作用影响结果[27-28], 而以6、7、8月为代表的夏季整体表现为谷值。图中6月底出现通量断点情况为仪器校准与维护导致通量与气象数据同时缺失。

|
| 图 5 荒漠杜加依林冠层CO2浓度和碳通量季节变化特征 Fig. 5 The typical seasonal variation characteristic of canopy interface carbon dioxide and carbon flux for Tugai forest in desert |
由图 6可以看出, 1月, 荒漠杜加依林CO2湍流通量均值为-0.006 mg m-2 s-1, 对应CO2储存通量为0.009 mg m-2 s-1, 此月平均NEE为较小正值(0.003 mg m-2 s-1), 表现为微弱的碳源, 2月CO2湍流通量与储存通量均为较小负值, 平均NEE为-0.008 mg m-2 s-1, 则为弱的碳汇, 而3月则转为弱的碳源(NEE=0.003 mg m-2 s-1), 4月, 伴随着气温的升高, 土壤释放出大量CO2, 表现为较强的碳源, 而5—9月为生长季, 生态系统光合作用大于呼吸作用, 因此整个生长季表现为荒漠杜加依林从大气界面吸收CO2, 5—9月随着气温的升高、土壤湿度的增加, 植物的生长达到峰值, 碳吸收速率也相对高于其他月份, 不同地区不同生态系统NEE在月尺度上略有差异[29]。而10—12月随着气温的逐渐降低, 整个荒漠杜加依林几乎停止光合作用, 而表现为较强的碳源。

|
| 图 6 2012年荒漠杜加依林冠层湍流通量、CO2储存通量及NEE比较 Fig. 6 Compare of canopy interface turbulent flux, CO2 storage flux and NEE for Tugai forest in desert during 2012 Fc:湍流通量, turbulent flux;NEE:生态系统净碳交换量, Net ecosystem exchange;Fs:二氧化碳储存通量, CO2 storage flux |
荒漠杜加依林冠层内温度与湿度在时空尺度上变化规律复杂, 将观测期温湿度数据集分别与NEE进行回归分析时无明显规律性。因此, 研究中从两方面分析荒漠杜加依林冠层界面水热变化对CO2交换的影响:(1)在冠层垂直空间尺度上, 选取生长季典型日(8月21—23日)2 m和27 m垂直温湿度差与NEE进行回归分析;(2)在季节时间尺度上, 利用冠层内全年日平均温湿度与NEE进行回归分析。结果如图 7所示。

|
| 图 7 荒漠杜加依林冠层NEE随垂直梯度温湿度差的变化趋势 Fig. 7 Canopy interface NEE change with the vertical atmospheric temperature and humidity gradient deference for Tugai forest in desert |
由图 7可以看出, 在冠层垂直空间尺度上, 垂直气温差介于-1—1℃之间时, 杜加依林往往表现为较强烈的碳吸收, 而垂直温差在1℃附近表现为拐点, 在1—12℃范围内对应NEE呈现出在0值附近波动趋势。对于艾比湖流域内荒漠杜加依林而言, 空气相对湿度对CO2交换过程的影响不容忽视, 与温度相似, 垂直梯度湿度差仅在日尺度上与NEE拟合关系较好(R2=0.3627, P < 0.01)(图 7), 梯度湿度差在5%左右表现为拐点, 小于5%表现为生态系统吸收CO2能量较强, 在5%附近多表现为CO2排放, 而较大的垂直湿度差下NEE变化平缓。
季节尺度下荒漠杜加依林冠层内气温与NEE具有较好的拟合结果(图 8), 其规律与垂直气温差规律相反, 在-20—18℃之间时, 全年日均NEE整体呈0值附近波动趋势, 而气温在18℃左右为NEE的下降点, 对应NEE多为负值, 表现为下垫面植被吸收CO2能力逐渐增强, 这一规律解释了图 6中5月上旬荒漠杜加依林NEE呈负增长现象。在季节尺度上NEE与空气相对湿度拟合关系较差(图 8), 但值得注意的是, 研究区空气相对湿度小于55%时, 生态系统NEE正负交替且多为负值, 而大于55%时, 荒漠杜加依林NEE则对应为较小负值或较大正值, 表现为生态系统弱的碳吸收和较强的碳排放过程。

|
| 图 8 荒漠杜加依林冠层NEE随季节温湿度的变化趋势 Fig. 8 Canopy interface NEE change with the seasonal atmospheric temperature and humidity for Tugai forest in desert |
植物冠层空间结构对近地面摩擦风速具有显著影响[30-31], 荒漠杜加依林非生长期冠层较小而间隙较大, 此时期摩擦风速较小, 如1月摩擦风速仅为0.125 m/s。当杜加依林进入生长期, 冠层逐渐增大增高, 叶片增多填补了冠层空隙, 导致下垫面摩擦风速随冠层高度和形状的变化而增大, μ*增大表明杜加依林冠层摩擦应力增加, 即对应的阻力增大[32], 此时期荒漠杜加依林大气稳定度同样表现为增强趋势。荒漠杜加依林在6月达到最大μ*, 而生长旺季的7、8月反而略小, 表明茂盛的冠层结构相对于生长阶段的冠层而言, 冠层粗糙度较低而表面更加平滑, 摩擦风速随之减小。荒漠杜加依林冠层大气稳定度在6—9月为中性, 表明生长季大气垂直运动平稳, 同时, 生长季内冠层高温干燥的环境有利于大气趋于中性。
植物冠层内不同高度处的温度波动产生了不同的微气象特征[33], 同时, 植被冠层稀疏程度与空气垂直温度变化及差异密切相关[34], 荒漠杜加依林冠层为稀疏型, 其冠层底部具有更大的风速, 有利于温度降低, 从而使得冠层界面上气温产生小于5℃的垂直变幅。吕萍等[9]对内蒙古腾格里沙漠研究发现空气相对湿度随高度的增加而减少, 地表水汽由下向高空蒸发输送, 表现出明显的季节特征, 地处干旱区的荒漠杜加依林与半干旱地区相似, 季节尺度上冠层湿度垂直变幅超过40%, 当杜加依林植物高度增加或覆盖程度增加时, 植物的水汽蒸腾作用增强, 同时, 摩擦风速的降低致使水平方向上水汽交换的减弱, 有利于下垫面湿环境的形成和增强[35], 此外, 季节性降水对湿度垂直变化也具有影响[36-37]。冠层相对稀疏的杜加依林, 垂直高度上的温湿度变化明显小于密集型冠幅, 其仍然表现出温湿梯度低层大高层小, 不存在明显逆温逆湿现象, 有利于冠层表面大气层结的稳定。
3.2 荒漠杜加依林冠层碳收支特征荒漠杜加依林不同季节典型日碳通量总体呈秋冬小而春夏大的规律, 夏季夜间通量多以正值为主且呈波动趋势, 秋季夜间通量亦表现为较小的正值和波动特征, 主要原因在于夏、秋季夜间风速多变, 通过影响冠层内温湿和储存通量影响了碳通量, 导致夜间碳通量波动较大。现有研究中普遍发现冬春时段不同生态系统均表现为碳源[38-40], 荒漠杜加依林冬季表现为碳源, 但仅在个别时段出现碳吸收现象。盐生荒漠生态系统生长季日碳通量最大值为-0.12 mg m-2 s-1[10], 与本研究结果相近。季节性碳通量在冬季变化较小, 在早春时期变化较大, 高浓度的CO2更加有利于碳的吸收, 并且随着季节变化这种吸收能力呈减弱趋势[41]。日平均最大碳通量多发生在维管束植物覆盖的高峰时期或土壤水分较高的时期, 春季土壤湿润程度的增加可能导致白天碳通量释放能力的增强[42], 季节性的土壤水分变化影响下垫面碳通量的收支, 对于荒漠杜加依林而言表现为春季的释放峰值和夏季的吸收峰值。
森林冠层由于树高、冠幅、林隙等空间结构特征的差异, 导致不同类型森林冠层碳存储量值和季节变化规律上差异明显[43], 荒漠杜加依林冠层CO2储存没有明显的季节规律性, 仅1月存在正值, 其他时期均表现为弱的碳储存。已有研究表明, 我国荒漠地区的NEE日均值约为0.006 mg m-2 s-1, 非生长季介于-0.034—0.022之间(11月到翌年4月), 在生长季(5月到10月)植被覆盖区域表现为碳汇, 此时日最大NEE变幅出现在9月从中午最高的-0.10 mg m-2 s-1, 到夜晚最低的0.035 mg m-2 s-1[44]。荒漠杜加依林日平均NEE为-0.026 mg m-2 s-1, 高于荒漠平均, 其较高的植被覆盖和高冠层特征决定了其生长季的高碳汇能力。
3.3 荒漠杜加依林冠层水热变化对碳收支影响大气温度通过影响下垫面光合和呼吸对生态系统碳收支产生影响[45], 同时, 植物冠层界面内的空气温度变化, 可在不同程度上影响生态系统呼吸的变化[46]。本研究发现, 荒漠杜加依林生长季2 m和27 m的垂直温度差与NEE存在较好的拟合关系(R2=0.7350, P < 0.01), 冠层垂直温差介于-1—1℃时对应生态系统较高的碳吸收。荒漠杜加依冠层内气温在季节尺度上与NEE具有较好的拟合效果(R2=0.5221, P < 0.01), 季节性温度升高伴随着较大的NEE负值, 表明季节尺度上温度的升高是驱动荒漠杜加依林NEE碳汇功能增加的主要环境因子。张法伟等[47]在青海湖北岸高寒草甸草原生态系统生长季研究中发现, 昼夜温差(Td)小于14.8℃时, NEE随着Td增大而减少, 即有利于生态系统碳积累, 反之则不利于生态系统碳积累, 适度的Td既不影响植被白天的光合作用又降低系统夜间呼吸。对于荒漠杜加依林而言, 冠层尺度上的温度梯度差和季节尺度上的气温变化均能强烈影响生态系统NEE, 垂直温差通过影响生态系统呼吸增加碳释放, 而季节气温变化则对生态系统光合和呼吸均具有影响, 表现为生态系统碳积累。
荒漠杜加依垂直冠层界面上湿度变幅较大, 生长季典型日垂直湿度差在5%以内时杜加依林具有较高的碳吸收, 而高的垂直湿度差则会抑制生态系统碳吸收, 主要是由于高的湿度差通过影响下垫面植物叶片气孔的开闭降低植物光合[48-49], 进而降低生态系统碳吸收。季节尺度上的空气湿度变化可影响大气凝结水, 进而对植物生理产生影响[50]。苗雨春[51]研究发现, 大兴安岭湿地生态系统NEE随着相对湿度的增加呈线性增大的趋势, 说明在光合有效辐射不再是生态系统NEE的限制因素时, 较高的大气湿度也有利于生态系统净碳吸收, 同时高湿环境下(60.6%—60.8%)植物最大羧化速率增加, 通过增加植物光合能力而间接增加生态系统碳吸收[52], 而荒漠杜加依林中较高的湿度条件下(> 55%)反而会抑制生态系统对大气CO2的吸收, 这与冠层垂直湿度变化对碳收支的影响相似, 除由于高湿环境对荒漠植物光合生理的影响外, 与高湿环境对群落呼吸的激发同样密切相关。
4 结论本文利用涡度相关技术对艾比湖流域内荒漠杜加依林冠层界面气象环境特征和CO2交换过程进行了分析研究, 主要结论如下:
(1) 荒漠杜加依林植物冠层上方摩擦风速与湍流动能均表现为随月份变化的先增加后减小的趋势, 生长季(6—9月)大气稳定度为中性, 非生长季(1—5月、12月)为不稳定, 全年大气稳定度为-0.194, 呈现不稳定水平。
(2) 气温随着高度升高呈递增趋势, 变幅均小于5℃, 空气相对湿度呈秋冬湿春夏干特点, 垂直变幅超过40%。
(3) 大气碳通量呈现出秋冬小而春夏大的规律, 春季和夏季日碳通量分别为0.188 mg m-2 s-1和-0.102 mg m-2 s-1;荒漠杜加依林年碳收支表现为碳汇。
(4) 在日尺度上垂直温湿度差与NEE拟合结果较好, 对应拐点分别为1℃和5%, 而在季节尺度上温度与NEE拟合较好, 对应拐点分别为18℃和55%。总体而言, 生长季较小的冠层垂直温差有利于杜加依林对大气CO2的吸收, 而冠层高湿环境则会抑制杜加依林对CO2的吸收。
| [1] |
Pan Y D, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988-993. DOI:10.1126/science.1201609 |
| [2] |
李德志, 臧润国. 森林冠层结构与功能及其时空变化研究进展. 世界林业研究, 2004, 17(3): 12-16. DOI:10.3969/j.issn.1001-4241.2004.03.003 |
| [3] |
Keenan T, García R, Friend A D, Zaehle S, Gracia C, Sabate S. Improved understanding of drought controls on seasonal variation in Mediterranean forest canopy CO2 and water fluxes through combined in situ measurements and ecosystem modelling. Biogeosciences, 2009, 6(8): 1423-1444. DOI:10.5194/bg-6-1423-2009 |
| [4] |
杨亚丽.森林冠层气象要素廓线模型构建及应用研究[D].沈阳: 沈阳农业大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10157-1016144702.htm
|
| [5] |
Leuzinger S, Körner C. Tree species diversity affects canopy leaf temperatures in a mature temperate forest. Agricultural and Forest Meteorology, 2007, 146(1/2): 29-37. |
| [6] |
任引, 薛建辉. 中亚热带甜槠常绿阔叶林小气候特征分析. 南京林业大学学报:自然科学版, 2008, 32(3): 14-18. |
| [7] |
马鸿儒, 吉春容, 李新建, 邹陈, 袁玉江. 天山中部白杨沟天然林区森林小气候观测与分析. 干旱区研究, 2011, 28(2): 251-254. |
| [8] |
何清, 缪启龙, 张瑞军, 艾力·买买提明, 刘新春, 霍文. 塔克拉玛干沙漠肖塘地区空气动力学粗糙度分析. 中国沙漠, 2008, 28(6): 1011-1016. |
| [9] |
吕萍, 董治宝, 张正偲, 赵爱国. 腾格里沙漠近地面层风、气温、湿度特征. 中国沙漠, 2009, 29(5): 977-981. |
| [10] |
刘冉, 王勤学, 唐立松, 李彦. 盐生荒漠地表水热与二氧化碳通量的季节变化及驱动因素. 生态学报, 2009, 29(1): 67-75. DOI:10.3321/j.issn:1000-0933.2009.01.009 |
| [11] |
Koerner B A, Klopatek J M. Carbon fluxes and nitrogen availability along an urban-rural gradient in a desert landscape. Urban Ecosystems, 2010, 13(1): 1-21. DOI:10.1007/s11252-009-0105-z |
| [12] |
Slot M, Garcia M N, Winter K. Temperature response of CO2 exchange in three tropical tree species. Functional Plant Biology, 2016, 43(5): 468-478. DOI:10.1071/FP15320 |
| [13] |
龚雪伟, 吕光辉. 艾比湖流域杜加依林荒漠植物群落多样性及优势种生态位. 生物多样性, 2017, 25(1): 34-45. |
| [14] |
Thevs N. Ecology, Spatial Distribution, and Utilization of the Tugai Vegetation at the Middle Reaches of the Tarim River, Xinjiang, China. Greifswald, Germany: Ernst-Moritz-Arndt-University Greifswald, 2006.
|
| [15] |
Zerbe S, Halik V, Küchler J. Urban greening in the oases of continental arid Southern Xinjiang (NW China)-an interdisciplinary approach. Die Erde, 2005, 136(3): 245-266. |
| [16] |
Thevs N, Zerbe S, Schnittler M, Abdusalih N, Succow M. Structure, reproduction and flood-induced dynamics of riparian Tugai forests at the Tarim River in Xinjiang, NW China. Forestry, 2008, 81(1): 45-57. DOI:10.1093/forestry/cpm043 |
| [17] |
Leemans R, Eickhout B. Another reason for concern:regional and global impacts on ecosystems for different levels of climate change. Global Environmental Change, 2004, 14(3): 219-228. DOI:10.1016/j.gloenvcha.2004.04.009 |
| [18] |
陈亚宁, 李稚, 范煜婷, 王怀军, 方功焕. 西北干旱区气候变化对水文水资源影响研究进展. 地理学报, 2014, 69(9): 1295-1304. |
| [19] |
于贵瑞, 孙晓敏. 中国陆地生态系统碳通量观测技术及时空变化特征. 北京: 科学出版社, 2008: 135-161.
|
| [20] |
Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Clement R, Dolman H, Granier A, Gross P, Grünwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Lai C T, Law B E, Meyers T, Moncrieff J, Moors E, Munger J W, Pilegaard K, Rannik V, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T, Wilson K, Wofsy S. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 2001, 107(1): 43-69. DOI:10.1016/S0168-1923(00)00225-2 |
| [21] |
黄昆, 王绍强, 王辉民, 仪垂祥, 周蕾, 刘允芬, 石浩. 中亚热带人工针叶林生态系统碳通量拆分差异分析. 生态学报, 2013, 33(17): 5252-5265. |
| [22] |
Hollinger D Y, Kelliher F M, Byers J N, Hunt J E, McSeveny T M, Weir P L. Carbon dioxide exchange between an undisturbed old-growth temperate forest and the atmosphere. Ecology, 1994, 75(1): 134-150. |
| [23] |
Griffis T J, Black T A, Morgenstern K, Barr A G, Nesic Z, Drewitt G B, Gaumont-Guay D, McCaughey J H. Ecophysiological controls on the carbon balances of three southern boreal forests. Agricultural and Forest Meteorology, 2003, 117(1/2): 53-71. |
| [24] |
杨振, 张一平, 于贵瑞, 宋清海, 高举明, 孙晓敏, 窦军霞, 赵双菊, 谭正洪. 西双版纳热带季节雨林大气稳定度特征. 生态学杂志, 2008, 27(1): 130-134. |
| [25] |
于跃飞.新疆局地气候研究[D].杭州: 浙江大学, 2006. http://cdmd.cnki.com.cn/Article/CDMD-10335-2007099011.htm
|
| [26] |
王超, 韦志刚, 高晓清, 李振朝, 侯旭宏, 刘慧, 魏红. 夏季敦煌稀疏植被下垫面物质和能量交换的观测研究. 高原气象, 2012, 31(3): 622-628. |
| [27] |
Holst J, Liu C, Yao Z, Brüggemann N, Zheng X, Giese M, Butterbach-Bahl K. Fluxes of nitrous oxide, methane and carbon dioxide during freezing-thawing cycles in an Inner Mongolian steppe. Plant and Soil, 2008, 308(1/2): 105-117. |
| [28] |
秦璐, 吕光辉, 何学敏, 张雪妮, 张雪梅, 孙景鑫, 李尝君, 杨晓东. 艾比湖地区土壤呼吸对季节性冻土厚度变化的响应. 生态学报, 2013, 33(22): 7259-7269. |
| [29] |
王国印.半干旱区陆气相互作用的观测与研究[D].兰州: 兰州大学, 2013.
|
| [30] |
Zhang G, Leclerc M Y, Karipot A. Local flux-profile relationships of wind speed and temperature in a canopy layer in atmospheric stable conditions. Biogeosciences, 2010, 7(11): 3625-3636. DOI:10.5194/bg-7-3625-2010 |
| [31] |
Walter B, Gromke C, Leonard K C, Manes C, Lehning M. Spatio-temporal surface shear-stress variability in live plant canopies and cube arrays. Boundary-Layer Meteorology, 2012, 143(2): 337-356. DOI:10.1007/s10546-011-9690-5 |
| [32] |
张强, 曾剑, 姚桐. 植被下垫面近地层大气动力状态与动力学粗糙度长度的相互作用及其参数化关系. 科学通报, 2012, 57(8): 647-655. |
| [33] |
Hirai T L, Carmona-Galindo V D, Samuelson S, Hale C, Braker E. Sub-Canopy temperature dynamics of a native tree plantation from a lowland tropical rainforest in Costa Rica. Open Journal of Forestry, 2014, 4(3): 191-196. DOI:10.4236/ojf.2014.43025 |
| [34] |
Tesfuhuney W A, Walker S, van Rensburg L D, Everson C S. Water vapor, temperature and wind profiles within maize canopy under in-field rainwater harvesting with wide and narrow runoff strips. Atmosphere, 2013, 4(4): 428-444. DOI:10.3390/atmos4040428 |
| [35] |
洪雯, 王毅勇, 聂晓. 湿地下垫面大气边界层温、湿廓线的数值模拟及影响因素分析. 气候与环境研究, 2011, 16(4): 494-504. DOI:10.3878/j.issn.1006-9585.2011.04.10 |
| [36] |
涂钢, 刘辉志, 董文杰. 半干旱区不同下垫面近地层湍流通量特征分析. 大气科学, 2009, 33(4): 719-725. DOI:10.3878/j.issn.1006-9895.2009.04.06 |
| [37] |
Matsuura S, Miyata A, Mano M, Hojito M, Mori A, Kano S, Sasaki H, Kohyama K, Hatano R. Seasonal carbon dynamics and the effects of manure application on carbon budget of a managed grassland in a temperate, humid region in Japan. Grassland Science, 2014, 60(2): 76-91. |
| [38] |
宋霞, 刘允芬, 徐小锋, 于贵瑞, 温学发. 红壤丘陵区人工林冬春时段碳、水、热通量的观测与分析. 资源科学, 2004, 26(3): 96-104. DOI:10.3321/j.issn:1007-7588.2004.03.015 |
| [39] |
Zhou X P, Wang X K, Tong L, Zhang H X, Lu F, Zheng F X, Hou P Q, Song W Z, Ouyang Z Y. Soil warming effect on net ecosystem exchange of carbon dioxide during the transition from winter carbon source to spring carbon sink in a temperate urban lawn. Journal of Environmental Sciences, 2012, 24(12): 2104-2112. DOI:10.1016/S1001-0742(11)61057-7 |
| [40] |
麦博儒, 安兴琴, 邓雪娇, 周凌晞, 王春林, 李菲, 黄建平, 陈玲, 尹淑娴. 珠江三角洲近地层CO2通量模拟分析与评估验证. 中国环境科学, 2014, 34(8): 1960-1971. |
| [41] |
Jasoni R L, Smith S D, Arnone J A. Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2. Global Change Biology, 2005, 11(5): 749-756. DOI:10.1111/gcb.2005.11.issue-5 |
| [42] |
Wohlfahrt G, Fenstermaker L F, Arnone Ⅲ J A. Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem. Global Change Biology, 2008, 14(7): 1475-1487. DOI:10.1111/j.1365-2486.2008.01593.x |
| [43] |
Vourlitis G L, de Souza Nogueira J, Filho N P, Hoeger W, Raiter F, Biudes M S, Arruda J C, Capistrano V B, de Faria J L B, de Almeida Lobo F. The sensitivity of diel CO2 and H2O vapor exchange of a tropical transitional forest to seasonal variation in meteorology and water availability. Earth Interactions, 2005, 9(27): 1-23. DOI:10.1175/EI124.1 |
| [44] |
Gao Y H, Li X R, Liu L C, Jia R L, Yang H T, Li G, Wei Y P. Seasonal variation of carbon exchange from a revegetation area in a Chinese desert. Agricultural and Forest Meteorology, 2012, 156: 134-142. DOI:10.1016/j.agrformet.2012.01.007 |
| [45] |
李婧, 刘树华, 茅宇豪, 张称意, 刘立超, 梁福明, 辛国君, 王建华. 不同生态系统CO2通量和浓度特征分析研究. 地球物理学报, 2006, 49(5): 1298-1307. DOI:10.3321/j.issn:0001-5733.2006.05.008 |
| [46] |
唐祥, 陈文婧, 李春义, 查天山, 吴斌, 王小平, 贾昕. 北京八达岭林场人工林净碳交换及其环境影响因子. 应用生态学报, 2013, 24(11): 3057-3064. |
| [47] |
张法伟, 李英年, 曹广民, 李凤霞, 叶广继, 刘吉宏, 魏永林, 赵新全. 青海湖北岸高寒草甸草原生态系统CO2通量特征及其驱动因子. 植物生态学报, 2012, 36(3): 187-198. |
| [48] |
Ogle K, Reynolds J F. Desert dogma revisited:coupling of stomatal conductance and photosynthesis in the desert shrub, Larrea tridentata. Plant, Cell & Environment, 2002, 25(7): 909-921. |
| [49] |
苏培玺, 严巧娣. C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征. 生态学报, 2006, 26(1): 75-82. |
| [50] |
秦璐.不同盐生境下盐穗木群落凝结水形成机制及其生态效应[D].乌鲁木齐: 新疆大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10755-1014384303.htm
|
| [51] |
苗雨春.大兴安岭多年冻土区泥炭沼泽地——气间净碳通量交换[D].北京: 中国科学院大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-80062-1013025779.htm
|
| [52] |
Bauerle W L, Daniels A B, Barnard D M. Carbon and water flux responses to physiology by environment interactions:a sensitivity analysis of variation in climate on photosynthetic and stomatal parameters. Climate Dynamics, 2014, 42(9/10): 2539-2554. |