 2019, Vol. 39
2019, Vol. 39文章信息
- 严光荣, 杨永清, 唐安军
- YAN Guangrong, YANG Yongqing, TANG Anjun
- 前自交型小红花寄生种子特性及寄生关系的建立
- Investigating seed characteristics and parasitic relationship establishment of Scurrula parasitica var. graciliflora characterized by prior selfing
- 生态学报. 2019, 39(3): 1019-1029
- Acta Ecologica Sinica. 2019, 39(3): 1019-1029
- http://dx.doi.org/10.5846/stxb201710181874
-
文章历史
- 收稿日期: 2017-10-18
- 网络出版日期: 2018-11-15

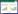




寄生是一种比较常见的生物有机体之间的互作关系, 广泛存在于自然界。在被子植物种, 寄生植物的种类大约有3000种, 约占被子植物总数的1%, 分属于16个科[1];抑或有4744种, 分属于32个科[2]。寄生植物的共性之一就是通过吸器从寄主获取水分、营养等生长发育所需物质[3-4]。种子萌发之后, 吸器逐渐形成, 并由此与寄主建立寄生关系, 实现从自养阶段到寄生阶段的转变[5-6]。一旦建立了寄生关系, 寄生植物不仅能改变寄主的行为, 影响寄主与非寄主之间的相互作用, 而且能驱动生态系统中生物与非生物因子的变化, 从而影响生物多样性, 进而改变生态系统的结构和功能[7-8]。
繁殖是生物进化的关键, 也是研究各种生物进化问题的重要节点[9]。一般地, 植物的有性繁殖涉及三个连续事件:传粉、种子传播和幼苗建成;而且, 传粉和种子传播通常依赖于动植物之间的相互作用[10]。传粉是种子植物受精的必经阶段, 其运动方式在很大程度上限定了植物个体间的基因流和群体的交配方式, 从而影响后代的遗传组成和适合度[11]。传粉者服务的减少能直接影响植物的繁殖产出, 会导致果实和种子产量与质量的下降, 甚至促进植物转向自交[12]。此外, 种子传播受限会严重地影响种子萌发和幼苗建成, 进而加速自交衰退[13]。就寄生植物而言, 种子的有效传播对于种子萌发是非常重要的。越来越多的研究表明, 鸟或其他动物的取食作用不仅能传播种子, 而且对某些种子的萌发有促进作用[6, 14]。种子休眠与萌发是植物适合度的重要组成部分[9, 12]。许多寄生植物的种子是休眠的, 需经过化学物质的刺激才能萌发[5, 15-16]。在某些情况下, 由于干燥失水或高温等逆境胁迫, 脆弱的幼苗由于未能及时与寄主建立寄生关系而死亡[17]。无论是植物的传粉方式, 还是种子萌发模式, 抑或是寄生植物与寄主之间的相互作用, 都是在自然选择的作用下使生物个体的适合度趋向最大[18]。
桑寄生科(Loranthaceae)是一类比较常见的寄生植物, 约有22属700余种, 其中一些种类有多个寄主, 如Dendrophthoe falcate[19]。长期以来, 尽管在桑寄生植物的分类鉴定、种子传播与种子萌发等方面取得了长足进展[1, 19], 但是, 许多寄生植物的繁育系统和传粉机制等方面的研究亟待弥补[6, 20]。譬如, 小红花寄生(Scurrula parasitica var. graciliflora)有多种寄主, 具有重要的药用价值[21], 有关研究主要集中于物种分类[22]以及果实的传播等方面[14], 尚无繁殖生态及进化等方面的科学报道。调查发现, 在重庆缙云山下的杨家沟和肖家沟, 成熟的小红花寄生果实在5—6月散落于树枝上和地面, 但缺乏寄生的小苗。那么, 小红花寄生幼苗罕见的原因是什么呢?又是哪些因素影响其寄生成功的呢?为了充分地理解上述问题, 我们对其繁育系统、种子萌发特性、种子的脱水耐性、寄生关系建立等方面进行了系统调查与研究。
1 材料和方法 1.1 小红花寄生及其生境在重庆市沙坪坝区的杨家沟(29°36′49″N, 106°18′07″E, 海拔275 m)和肖家沟(29°36′48″N, 106°18′07″E, 海拔353 m), 小红花寄生(S. parasitica var. graciliflora)分别寄生在数十株桂花树上。小红花寄生是桑寄生科中的一种常绿灌木, 总状花序有花3—7朵, 密集于叶腋;花冠黄色, 长1—1.2 cm, 裂片披针形, 长约3 mm;果梨形, 红黄色;花果期4—12月;主要分布于云南、四川、贵州、广西等省区的一些山谷或阔叶林, 其海拔介于850—2100 m, 常以桃树(Prunus persica)、锥栗(Castanea henryi)等种类为寄主;尼泊尔、缅甸、孟加拉等国也有分布[21]。经过2年的连续观察, 研究发现, 寄生于桂花树上的小红花寄生的花期是3—5月, 果期是5—6月。在果实成熟期间, 红胸啄花鸟(Dicaeum ignipectus)和白头鹎(Pycnonotus sinensis)等会偶尔取食小红花寄生的成熟果实。
在相邻的杨家沟和肖家沟, 与小红花寄生的寄主桂花伴生的植物有桑树(Morus alba)、紫叶李(Prunus Cerasifera)、桃树(P. persica)、小叶女贞(Ligustrum quihoui)和黄葛树(Ficus virens var. sublanceolata)等多种植物, 它们共同构成了一个小型的“常绿-落叶阔叶林”, 伴生的草本植物有千里光(Senecio scandens)、狗牙根(Cynodon dactylon)、蛇莓(Duchesnea indica)和喜旱莲子草(Alternanthera philoxeroides)等种类。杨家沟和肖家沟的气候类型属于中亚热带季风性湿润气候, 四季分明, 雨量充沛, 且降雨多发于5—10月(约占全年降雨量的80%)。2005—2016年, 此地的最冷月(1月)的平均气温8.2℃, 最热月(7月, 有的年份是8月)的平均气温29.3℃, 年平均气温19.2℃, 年均降水量为1086 mm(数据源自沙坪坝区气象局)。
1.2 花部特征与开花物候的测量与观察2015年3—6月, 首先观察和分析了小红花寄生的开花动态及其开花比例, 以此确定其开花物候。其次, 随机标记20个花蕾, 连续观察, 以确定单朵花的持续时间;第三, 在花蕾期标记30个花蕾, 分别测量花开之后的花筒长度、花筒基部直径、花筒顶部直径、花瓣裂片的长度、花丝长度、花药长度以及花梗的长度。2016年3—6月, 重复观察和记录小红花寄生的开花动态, 并测量上述参数。
1.3 传粉处理与昆虫传粉行为的观察2015年5—6月, 为了查明小红花寄生的繁育系统类型, 分别进行如下处理:(1)自然授粉的对照组(CK)。在花蕾期, 随机选择50朵花, 挂上塑料标记牌, 让其自然开放与授粉。(2)花蕾期挂牌标记, 套袋(marked and bagging before corolla tubes split, MBB)。在花冠筒直立(花冠筒闭合)时, 在5株小红花寄生上随机标记50朵花, 并套上小的茶叶袋。(3)花开放之前挂牌标记, 去雄, 不套袋(marked, emasculated, and non-bagging before corolla tubes split, MENBB)。在花冠筒直立时, 用小镊子将花冠筒打开, 去除雄蕊, 处理朵50朵花。(4)花开放之前挂牌标记, 去雄, 花开之后异花授粉, 并套袋(marked, emasculated, outcrossing and bagging before corolla tubes dehisce, MEOBB)。在花冠筒尚未弯曲前, 对50朵随机标记的花予以去雄并套袋, 然后在花开之后从其他开放的花上收集花粉, 辅以人工授粉, 继续套袋;(5)在花冠筒开裂之前挂牌标记, 去雄, 套袋(marked, emasculated and bagging before corolla tubes split, MEBB)。在花冠筒直立闭合时, 对标记的50朵花进行去雄处理, 然后套袋。2016年5—6月, 在同一个种群内重复上述5类处理, 进一步探究其繁育系统的特性。
此外, 在2015年和2016年的开花高峰期, 分别连续3天对小红花寄生进行传粉昆虫的监测。观察时间为6:00—19:00, 每次观察20 min, 1小时内随机观察1次;同时, 如果有昆虫前来访花, 我们就对这些昆虫进行拍照与捕捉, 将捕捉的昆虫放入装有一团医用棉花和少量乙酸乙酯的玻璃瓶中, 而后在实验室进行昆虫种类的鉴定。
在果实成熟期间(5月下旬), 收集以上5种处理的果实, 统计座果率(%)(即正常发育的果实数与标记处理的总花数的百分比)。一般地, 一个正常发育的果实含有一粒种子。
1.4 花粉活力和柱头可授性的检测2015年5月, 随机标记100朵花, 以收集花粉和柱头。在花冠略弯曲且尚未裂开而花药已裂开时, 取一次花粉并检测活力;花冠裂开之后, 每隔24 h取一次花粉并检测其活力, 连续取4次。离体花粉活力的检测参照叶要妹等所描述的方法进行[23]。对每一次所取的花粉样品设置4个重复培养, 通过体视显微镜观察花粉的萌发情况, 以花粉管长度大于花粉直径作为萌发的标准。
随机取样柱头并检测其可授性。花开放之前(花弯曲, 花冠未开裂), 但花药已开裂, 连续3天于10:00取10个花柱。花开放之后(此时花冠裂片反折), 连续3天取花柱, 分别是10:00点和5:00各取10个花柱。取样之后, 立即用联苯胺-过氧化氢溶液检测柱头的可授性[24]。具体操作如下:将新采集的柱头浸入联苯胺-过氧化氢溶液(1%联苯胺:3%过氧化氢:水= 4:11:22, V/V)中(该溶液加注于凹面载玻片的凹陷处)。10—15 min后, 在体视显微镜(舜宇体视显微镜/SZM45, 浙江宁波舜宇仪器有限公司)下观察柱头染色的部位, 若柱头有2/3及其以上的面积呈现深蓝色并伴有大量气泡, 则视为可授性柱头, 否则视为无活性的柱头。
1.5 花粉扫描电镜观察首先, 从花冠筒已开裂的10朵花中分别采集花粉, 然后将花粉混合。其次, 从花冠筒尚未开裂的10朵花中分离出柱头。随后, 用双面胶在铜台上粘样, JFC-1200离子溅射镀膜仪喷金镀膜, 用SU3500日立扫描电子显微镜观察与拍照。随机选取20离花粉, 测量臂长及高度(μm);分别观察和测量10个柱头的表面直径(μm), 并拍照。
1.6 果实的采集、特征参数测量及萌发实验2015年5月, 从小红花寄生的植株上采集成熟的果实, 一部分果实用于测量果实的大小(cm)、千粒重(g)和含水量(%)等基本特征值;其余的果实用于萌发(即带果肉的萌发实验)和剥取种子。随机取30粒果实, 用数显游标卡尺(精度为0.01 mm)测量果实的长度与宽度。随机取200粒果实, 用电子天平(精度为0.1 mg)称其重量(g), 3次重复。在测定果实的含水量时, 参照ISTA的方法进行[25]:取10粒果实, 称其重量(W0), 然后将其放入103℃的烘箱中进行干燥脱水, 17 h后取出, 放入密闭的干燥器中冷却至室温, 然后称重(W1), 并以鲜重为基础计算果实的初始含水量(%), 重复3次。
在“果实内的种子萌发实验”前, 用2%的次氯酸钠溶液对果实进行表面消毒10 min, 然后进行萌发检测:(1)在邻近小红花寄生的生境(距离小红花寄生母株约3 m)中, 将50粒种子播种于大小为1 m×1 m的小样块的土壤表面, 4个重复。(2)在实验室, 将25粒种子摆放于培养皿内湿润的滤纸上, 然后将其放入25℃、30/20℃或20℃的周期性光照培养箱内。每个温度下设立4个重复。以胚根突破“果皮”作为萌发的标准, 实验持续时间为28 d。
1.7 温度和光照对带果肉种子萌发的影响2015年5月, 在5个恒温(15、20、25、30℃和35℃)和3个变温(20/10℃、25/15℃以及30/20℃)下检测小红花种子的萌发特性, 均给予周期性光照(12 h/d), 光照强度约为40 μmol m-2 s-1。用于萌发的种子是去除了果皮而具粘液物质的种子, 即是“带果肉的种子”。变温的高温时段与光照同步, 其低温时段是黑暗的。检测种子萌发时, 在玻璃培养皿(Φ=9 cm)内置2层滤纸, 并加入9 mL灭菌水, 然后放置25粒带果肉的种子。每一个温度下设立4个重复。以胚根长至2 mm作为种子萌发的标准, 观测持续时间为28 d(其他的萌发实验与此相同)。为了避免系统位置效应, 观察时随机地改变培养皿的位置(下同)。每天计数萌发的种子, 并将萌发的种子移出培养皿。
2016年5月, 检测种子在黑暗下的萌发情况。在培养皿(Φ=6 cm)的滤纸上放置25粒种子, 加入5 mL蒸馏水, 然后用双层铝箔纸包裹培养皿;萌发温度为25℃和30/20℃, 每个温度下设置4个重复, 其他的操作和萌发标准与光照下的相同。在实验的第28 d查看种子萌发情况, 以胚根长至2 mm作为萌发的标准。
1.8 种子的干燥脱水与萌发检测2016年6月, 基于萌发率对小红花寄生种子进行脱水耐性的评价。首先, 从所采集的成熟果实中手工分离种子(即是“不带果皮和果肉的种子”), 并清洗干净, 室温下晾干, 然后贮藏于15℃的恒温箱中(相对湿度为65%)。其次, 在带盖的玻璃干燥器内的活性硅胶的表面放置120粒种子, 进行脱水处理(温度控制在26±2℃), 共用5个干燥器处理了600粒种子, 分别脱水2、12、48、72 h和96 h。对脱水72 h和96 h的干燥器内的硅胶分别更新2次和3次, 以获得更好的脱水效果。当种子脱水至既定的时间后, 取出任意一个干燥器中的种子, 其中20粒用于含水量(%)的测定, 其余100粒用于萌发实验。测定含水量时, 参照国际种子检测协会(ISTA)制定的方法进行[25], 4次重复。以未干燥脱水的种子作为对照。萌发温度为25℃。
1.9 与桂花和桃树的寄生关系的建立2016年5月, 我们将分离的具有粘液物质的种子放于桂花(Osmanthus fragrans)和桃树(P. persica)的不同部位, 以评估其寄生关系建立的难易度。具体操作如下:在5株成年桂花的茎干(胸径约110—140 mm)表面(包括茎与枝的连接处)分别放置16粒, 共计80粒种子。在10株桂花幼树的茎顶叶腋(茎的直径约3—4 mm)分别放置8粒种子, 共计80粒种子。同时, 在10株桂花幼树的叶片上也放置80粒种子, 每一株上有种子8粒。类似地, 在5株成年桃树茎干、叶腋和叶片上分别放置80粒种子, 即分为茎干、茎顶叶腋和叶片3组。特别地, 在放置种子前, 用钻孔器在桂花和桃树的茎干上打一个小孔(孔的直径与深度约为2—3 mm), 种子被置于孔内。每2天观察一次种子的发芽动态及幼苗的生长状况。在观察计数时喷洒少量的蒸馏水, 防止种子因干燥脱水而死亡。
1.10 数据统计分析花粉活力(%)和柱头可授性(%)以重复实验的平均值±标准偏差(SD)表示。运用单因子方差分析(One-way ANOVA)和最小显著差数法(LSD)检验各种传粉处理对座果率(%)的影响;利用单因子方差分析检验温度、脱水处理或人工接种位点对种子萌发的作用效应。因为只有桂花幼树茎顶叶腋的幼苗与寄主桂花以较低比例的建立了寄生关系, 故未对寄生率进行统计检验。在进行方差分析时, 先将萌发率(%)和寄生率(%)进行反正弦转换, 但萌发与寄生的结果仍以未转换前的数值(%)表示。所用数据统计分析软件是SPSS 19.0 for Windows。
种子萌发率(%)以平均数±标准偏差(SD)表示。种子平均萌发时间(MGT)的计算公式如下:
MGT=∑(t×n)/∑n[26]
式中, t是萌发时间(d), 计时从0 d开始;n为第t天所萌发的种子数。MGT的值越大, 则种子萌发速率越慢。
2 结果 2.1 小红花寄生的开花物候与花部特征小红花寄生的花序为总状花序, 4—7朵生于叶腋处, 外被白色短绒毛(图 1A, 1B)。在2015年3月21—28日期间, 较多绿色的花蕾形成, 称之为“花早期”或“花蕾期”。在3月29日—4月17期间, 约有10%的花开放, 称为“始花期”。在4月18日—5月29日期间, 约有95%的花开放, 称之为“盛花期”或“开花高峰期”。在5月30日—6月14日期间, 大部分花已凋谢, 约有5%的花仍在开放, 称之为“晚花期”。2016年, 小红花寄生的开花物候没有明显变化, 故未赘述有关数据。

当花开放之后, 花柱呈紫红色, 柱头呈暗红色或红褐色(图 1B);花柱的长度约(26.67±0.25) mm, 柱头的表面宽度约(35.70±0.02) μm(图 1G)。与此同时, 略弯曲的花冠筒的长度约(27.94±0.22) mm, 花冠筒的4个裂片反折, 甚至紧贴花冠, 其裂片长度约(5.37±0.13) mm;花冠下部呈红色, 上部黄色或黄绿色(图 1B)。在花冠内壁, 紫红色条带与绿色条带相间排列, 4条紫红色条带实为雄蕊的扁平状花丝, 与花冠紧贴, 但可分离。花丝上端(约(1.75±0.03) mm)呈红色。花邻近开放时, 花冠筒的基部直径、中部直径和顶部直径分别为(2.20±0.08)、(2.66±0.10) mm和(1.46±0.08) mm。
花柱初始期为绿色。随着花的生长, 花柱逐渐由绿色变成黄色, 最后转变为红色。柱头由红色转变为红褐色, 具粘液。当花冠筒裂开柱头可见时, 柱头呈紫黑色或暗紫色, 粘液消失。雄蕊始终位于柱头之上, 略高于柱头(约高出0.75 mm), 4个花药(贴着药)呈黄色, 长约(2.04±0.02)mm, 瓣裂。在花冠裂开之前, 花粉粒已散出并落置于具粘液的柱头之上。黄色的花粉粒呈三沟型-三臂沟型, 中央有一小孔, 在电子显微镜下, 花粉粒外壁的表面密布瘤状突起(图 1E, F)。花粉的臂长为(20.37±1.52) μm, 花粉的中央高度为(19.73 ±1.77) μm。在花开之后呈暗红色, 其上有许多乳突(图 1G, H)。柱头表面的直径是(367 ± 21) μm。单花的寿命约7—8 d。在整个花期, 花不分泌花蜜。
2.2 花粉活力与柱头的可授性花粉活力随着开花时间的推移而减弱, 花粉活力与柱头可授性是同步变化的。花冠筒已弯曲但未开裂时, 花粉活力极高(98%)。在花开之后的第24、48、72 h和96 h, 花粉活力依次为94.5%、81.5%、38.3%和6.3%, 存在显著差异(F=340.507, df = 4, P < 0.001)。
随着开花时间的延长, 柱头可授性亦降低。花开之前, 3次取样的柱头可授性分别是100%、100%和95%±2.1%。花开之后, 6次取样的柱头可授性分别是73%±3.83%、65%±5.72%、49%±2.84%、50%±3.77%、25%±2.22%和4.7%±1.41%(F=81.900, df=8, P < 0.001)。
2.3 繁育系统与传粉特性传粉处理对小红花寄生的座果率有显著的影响(F = 40.722, df=4, P=0.001)。自然授粉的对照组(CK)的座果率为82.5%±4.08%;花开放之前标记且套袋的实验组(MBB)的座果率为83.1%±3.75%, CK与MBB的座果率没有显著差异(F=0.901, df =1, P=0.379)。花开之前, 去雄后异花授粉的实验组(MEOBB)的座果率为35%±4.56%, 与CK的座果率的差异显著(F=266.748, df=1, P < 0.001)。去雄的两个实验组(MENBB和MEBB)均未产生果实(图 2)。

|
| 图 2 不同传粉处理对小红花寄生座果率的影响 Fig. 2 Effects of different pollination treatments on fruit set (%) in Scurrula parasitica var. graciliflora CK, 对照, control check; MBB, 花冠筒开裂前标记和套袋, marked and bagging before corolla tubes split; MENBB, 花冠筒开裂前标记、去雄, 但不套袋, marked, emasculated, and non-bagging before corolla tubes split; MEOBB, 花冠筒开裂前标记、去雄、杂交并套袋, marked, emasculated, outcrossing and bagging before corolla tubes dehisce; MEBB, 花冠筒开裂前标记、去雄并套袋, marked, emasculated and bagging before corolla tubes split.不同小写字母表示不同处理的座果率在0.05水平上存在显著差异 |
在实验期间, 虽然有蜜蜂(Apis cerana cerana)和熊蜂(Bombus Spp.)在小红花寄生及寄主桂花的枝叶间飞行, 但是, 它们都没有停留在花朵之上取食花粉或花蜜。所以, 蜜蜂和熊蜂不是小红花寄生有效的传粉者。除了蜜蜂和熊蜂外, 尚未见到其他昆虫、鸟或其他动物来访花。
2.4 果实与种子的基本特征小红花寄生的果实成熟于5—6月, 浆果梨形, 红黄色或黄色(图 1C), 长约(9.42±0.03) mm, 直径(宽)是(4.69±0.16) mm, 千粒重为(112.32±9.76) g, 初始含水量是72.15%±1.71%。
小红花寄生的种子外被粘液物质, 主要由种皮、胚和胚乳等三部分构成(图 1)。种脐呈柄状, 位于种子底部, 长度为(4.58±0.07)mm;种脐向上延伸形成4根红色的种皮维管束。种子外壁呈粉红色, 种皮外的粘液物质呈乳白色。胚呈绿色(图 1D), 约占总体积的4/5, 较小的绿色胚((0.35±0.06)mm)镶嵌于胚乳中。种子千粒重为(19.18±1.44)g, 种子长度为(8.56±0.13)mm, 宽为(2.81±0.10)mm;种子初始含水量为53.01%±1.64%。
2.5 果实内的种子萌发在实验期间, 小红花生境的昼夜温度为17—32℃;3个萌发箱的温度分别是20℃、25℃和30/20℃。无论在小红花寄生生境中的小样块内, 还是在光照培养箱内, 小红花寄生的种子在果实中均未萌发。由此可见, 在完整的果实中, 种子萌发是受抑制的。
2.6 温度和光照对种子萌发的影响在5个恒温(15、20、25、30℃和35℃)和3个变温(30/20℃、25/15℃和20/10℃)下, 小红花寄生种子的萌发率显著不同(F =117.012, df = 7, P < 0.001)(图 3)。在周期性光照下, 20℃、25℃、30℃、30/20℃和25/15℃下的种子萌发率均≥96%。此外, 在上述8个温度下, 小红花寄生种子的平均萌发时间(MGT)也有显著差异(F=1.219E4, df=7, P < 0.001), 其中以25℃下的平均萌发时间最短(3.19 d)。另外4个恒温(15、20、30℃和35℃)的MGT分别是13.14、6.60、5.46 d和9.82 d, 其他3个变温(30/20℃、25/15℃和20/10℃)的MGT分别是8.67 d、11.49 d和12.35 d。由此可见, 在已试验的8个温度中, 最适宜的萌发温度是25℃, 其次是30℃、30/20℃和20℃;低温或高温都不同程度地抑制小红花种子萌发。

|
| 图 3 小红花寄生种子在不同温度下的萌发率(%) Fig. 3 Germination percentages of Scurrula parasitica var. graciliflora seeds which were incubated at different temperatures 不同小写字母表示种子萌发率在0.05水平上存在显著差异 |
在黑暗中, 25℃和30/20℃下的种子萌发率分别是92%±3.27%和91%±5.03%。在25℃下, 黑暗与光照的种子萌发率之间没有显著差异(F=9.000, df=1, P = 0.241);在30/20℃下, 黑暗与光照的种子萌发率之间也没有显著性差异(F=12.789, df =1, P=0.122)。由此可见, 光照条件对小红花种子萌发没有明显的影响。
2.7 种子的脱水耐性脱水过程中, 随着脱水时间的延长, 小红花寄生种子的含水量从初始时的53.06%逐渐下降到9.17%。干燥脱水对种子的萌发率有显著的影响(F=402.518, df = 5, P < 0.001)(图 4)。未脱水种子的萌发率可以达到97%;当脱水24 h, 含水量下降至22.27%时, 种子萌发率为36%;进一步脱水至9.17%时, 种子已完全失去活力。

|
| 图 4 脱水时间对小红花寄生种子含水量及萌发率的影响 Fig. 4 Effects of desiccation time on seed water content (%) and germination percentage of Scurrula parasitica var. graciliflora 不同字母表示种子萌发率在0.05水平上存在显著差异 |
人工接种于桂花和桃树不同位点的种子萌发率是有显著差异的(F=42.640, df = 5, P < 0.001), 其中桂花幼树茎顶叶腋的种子萌发率最高(90.6%), 而桃树叶片上的种子萌发率最低(47.2%)(表 1)。与寄主成功建立寄生关系的小苗均位于桂花幼树的茎顶叶腋, 其寄生率为6.25%(表 1)。
| 桂花Osmanthus fragrans | 桃树Prunus persica | ||||||
| 茎干 Stem |
茎顶叶腋 Axil close to the stem tip |
叶片 Leaf |
茎干 Stem |
茎顶叶腋 Axil close to the stem tip |
叶片 Leaf |
||
| 接种数量 Inoculation quality/n |
80 | 80 | 80 | 80 | 80 | 80 | |
| 种子萌发率 Seed germination percentage/%±SD |
86.3±3.6a | 90.6±2.4a | 66.9±3.9b | 70.6±4.6b | 75±2.7b | 47.2±8.0c | |
| 幼苗寄生率 Seedling parasitism rate/%±SD |
0 | 6.25±3.91 | 0 | 0 | 0 | 0 | |
| 同一行中不同小写字母表示在0.05水平上存在显著性差异 | |||||||
观察与实验结果表明, 半寄生的小红花寄生在自然条件下能大量的开花与结实, 且自花授粉的座果率达80%以上。成熟的种子在适宜的条件下能快速萌发。这表明小红花寄生的种子是非休眠的。但是, 萌发的种子却难以与寄主成功地建立寄生关系, 这极可能是小红花寄生小苗罕见的主要原因。
自花授粉的水平取决于花药与柱头之间的时空分离程度, 雌雄功能时空分离对植物传粉和交配格局有着直接的影响[18]。大多数被子植物产生完全花(既有雄蕊又有雌蕊), 并且其中一部分是自交亲和的, 完全自交的植物很少[27]。相比异交而言, 在种群密度低或缺少传粉者时, 自交可以提高植物的结实率(繁殖保障效应)和赋予自动选择优势[28]。尽管异交广泛存在于多种分布广泛的桑寄生植物中, 但是, 自交也是常见的交配模式[19]。小红花寄生的野生种群在4—6月开花, 持续约75 d;开花高峰期为4月18日—5月29日;单花寿命约7—8 d。小红花寄生的花是完全花, 在5种传粉类型中, 自然授粉的小红花寄生(对照组)有较高的坐果率(82.5%), 花开之后的异花授粉实验组(MEOBA)的座果率仅有33.8%。而且, 花蕾期套袋的实验组(MBB)在隔绝了异花传粉之后有着非常高的座果率(83.1%)。但是, 无论套袋与否, 去雄的花都不能结实。由此可见, 小红花寄生的繁育系统是兼性的, 以自花授粉为主, 而异花授粉在有效传粉后亦能发生, 但其座果率显著低于自交的座果率。在花冠筒弯曲而未裂开之前花药已裂开, 且花粉已落置于充分发育的粘性柱头上;花开之后, 柱头可授性显著降低, 而且柱头上的粘液物质消失。花药与和柱头的紧贴现象以及花粉活力和柱头可授性的变化趋势充分说明小红花寄生不存在雌雄异熟现象(即雌雄时间上的分离), 这为花内传粉与授精(自交)奠定了重要基础。类似地, 在没有传粉者服务时, 分泌花蜜的Ligaria cuneifoli和Tristerix corymbosus都是自交的[29]。同样, 墨西哥槲寄生(Psittacanthus calyculatus)在蜂鸟授粉受阻时能部分自交(44%的座果率), 而且去雄的花均不能产生果实[30]。
Darwin认为繁殖保障是自交进化的最重要因子[31]。根据自有花粉和外来花粉到达柱头的时间序列, 自交可分为3种不同的类型:前自交、竞自交和后自交;前自交是指自体授粉发生在异体授粉之前, 后自交是指自体授粉发生在异体授粉之后;竞自交是指自体授粉和异体授粉同时发生的情形[18, 32]。在开花期间, 小红花寄生的花没有分泌花蜜, 而且花筒长时间处于闭合状态;花开72 h后, 花粉和柱头呈紫黑色;而且, 花的香味极其淡雅。显然, 这样的花部特征不利于吸引昆虫或其他动物访花。可能正是这个原因, 在花期, 我们未能观察到真正的传粉昆虫或其他动物来访花。此外, 花开之后, 尽管花粉的活力能保持较长时间(约48 h), 但是, 72 h后柱头几乎全部失活。这暗示着异体授粉只能在开花之后较短时间内发生。开花之后, 如果传粉时间延后, 那么花粉在活性低的或失活的柱头上就不能有效授精。这极可能是花开之后异花授粉实验组(MEOBB)座果率低(33.8%)的原因。综上所述, 小红花寄生的交配模式属于前自交类型, 在一定条件下可能存在异花授粉现象。值得注意的是, 花开之后的异花授粉的有效性还受天气与传粉媒介等因素的影响。如果花期的天气或其他环境因子不利于异交传粉时, 那么小红花寄生就高度依赖于前自交的“闭花受精”, 以产生延续种族的种子(繁殖保障)。之前, Lord曾做了一个假设, 即闭花受精的植物一般能产生自由传粉的两性花, 但整个植株并不一定完全自交[33]。综合而言, 小红花寄生的交配模式是对上述假设的有力支持。
种子萌发与寄生关系的建立是寄生植物进化的重要环节[5, 15]。许多寄生植物的种子在成熟之后是休眠的, 需要在特殊的环境因子的刺激下才能萌发;萌发后种子的胚根要寄生到寄主植物上还必须通过诱导形成吸器来完成[16, 34]。例如, 根寄生植物独脚金(Striga asiatica)的种子在成熟后处于休眠状态, 寄主根系分泌的独脚金内酯能诱导种子萌发[35]。当然, 有的寄生植物的种子成熟后是非休眠的, 在适宜的条件下能够迅速萌发, 如五蕊寄生(Dendrophthoe pentandra)、栗寄生(Kortalsella japonica)等10种桑寄生植物的种子是非休眠的, 能在室温(18—32℃)下萌发[14]。与此类似, 小红花寄生的成熟种子在25、30、30/20℃和25/15℃下的萌发率均在96%以上, 但是在15℃下的萌发率为57%, 在35℃下萌发率为62%。由此可知, 非休眠的小红花寄生的种子萌发有明显的温度依赖性。当萌发温度为25℃和30/20℃时, 光照与黑暗下的萌发率都在90%以上。可见, 光照不是小红花寄生种子萌发的必要条件。
果实是种子的初始载体, 果皮和果肉的质地及其含有的化学物质直接影响果实及其种子的传播与萌发[36]。小红花寄生的果实是浆果, 成熟时呈红黄色或黄色, 果肉较多, 含水量也较高, 而且其乳白色的果肉极具粘性。此类特征为鸟取食小红花寄生果实并为之传播提供了可能性。不过, 在野外生境, 尽管偶尔见到了红胸啄花鸟(Dicaeum ignipectus)和白头鹎(Pycnonotus sinensis)停留枝头取食小红花寄生果实, 但未能收集到鸟食用果肉后留下的种子。因此, 尚不能证实鸟取食会对小红花寄生种子萌发产生何种影响。
在“果实萌发”实验中, 小红花寄生的种子在完整的果实中均不萌发。一旦去除果皮, 无论带果肉的种子, 还是不带果肉的种子, 都能在适宜的条件下萌发。这与肖来云和普正和以及罗亚皇和张玲对五蕊寄生(Dendrophthoe pentandra))种子萌发的研究结果是一致的[37-38]。
当去除果皮和果肉后, 小红花寄生的种子(即真正的“种子”)在干燥脱水过程中, 由于水分的丧失而表现出不同的萌发率。换而言之, 种子的含水量对种子萌发率有直接而显著的影响。当温度适宜时, 未脱水种子几乎能完全萌发;严重脱水至22.27%时, 仅有36%的萌发率。种子对干燥脱水的敏感程度是划分种子贮藏行为的首要依据(另一个依据是种子对贮藏温度的敏感性)[39]。迄今, 根据种子对干燥脱水的敏感程度, 种子被分为正常性种子、顽拗性种子和中间性种子三种基本类型;正常性种子对脱水不敏感, 其含水量降低至5%时仍能正常萌发;顽拗性种子对干燥脱水十分敏感, 在较高的含水量(20%—30%)时已严重劣变, 萌发率显著下降;中间性种子对干燥脱水的耐性介于二者之间, 一般在7%—12%的含水量时仍能正常萌发[40, 41]。综上可知, 小红花寄生的种子对干燥脱水是敏感的, 极可能是顽拗性种子。虽然环境中的自然干燥对种子萌发而言是一个限制因子, 但小红花种子在成熟时具有较高的含水量, 在雨热条件适宜的夏季能够快速萌发, 从而可以避免土壤微生物长时间的侵染而劣变, 进而提高其适合度。从进化生态学的角度看, 小红花寄生种子的非休眠性和脱水敏感性是适应水热的季节性变化格局的。
事实上, 去除果皮后, 带果肉的种子就失去了一道保护屏障。小红花寄生的果肉是高度粘性的, 有助于种子粘附于茎、树枝和叶片等媒介上。当食果鸟取食其果实后, 随粪便排出的是“没有粘性果肉”的种子。因此, 这样的种子又失去了粘性果肉的保护作用。当鸟粪自然变干后, 这种裸露的种子就暴露于空气中而逐渐干燥失水。所以, 小红花寄生的种子就难以萌发和形成吸器, 从而就不能与寄主建立寄生关系。
在人工建立寄生关系的研究中, 我们发现小红花种子可以在桂花和桃树的不同部位萌发, 但是仅在桂花幼树的茎顶叶腋处有6.25%的寄生率。尽管小红花寄生是广寄生的, 有多种寄主[21], 但是, 幼苗与潜在的寄主能否建立寄生关系在很大程度上受寄主部位的生理状态及解剖结构等因素影响。尽管寄生植物的种子萌发形成的幼苗产生了吸器, 但如果吸器不能与茎(枝条)和根的维管组织相联结, 那么就不能成功地建立寄生关系[3]。也就是说, 幼苗能否正常长成植株, 一个关键因素就是树体的抗寄生性, 这种抗寄生性与寄生部位的生理状态、器官解剖结构及硬度、化学物质的信号作用等因子密切相关[42]。小红花寄生的幼苗能在桂花幼树的茎顶叶腋处实现寄生关系的建立, 这说明桂花的幼树茎顶叶腋的抗寄生力较低, 也暗示了寄生植物对寄主(和寄主部位)表现出强烈的选择性。
| [1] |
Kuijt J. The Biology of Parasitic Flowering Plants. Los Angeles: University of California Press, 1969.
|
| [2] |
Parasitic plant genera & species.[2017-06-10]. http://www.parasiticplants.siu.edu/ParPlantNumbers.pdf.
|
| [3] |
Riopel JL, Timko MP. Haustorial initiation and differentiation//Press MC, Graves JD eds. Parasitic Plants. London: Chapman and Hall, 1995: 39-79.
|
| [4] |
Davis CC, Xi Z. Horizontal gene transfer in parasitic plants. Current Opinion in Plant Biology, 2015, 26: 14-19. DOI:10.1016/j.pbi.2015.05.008 |
| [5] |
宋文坚, 金宗来, 曹栋栋, 唐桂香, 周伟军. 寄生植物种子萌发特异性及其与寄主的识别机制. 应用生态学报, 2006, 17(2): 335-339. DOI:10.3321/j.issn:1001-9332.2006.02.035 |
| [6] |
Těšitel J. Functional biology of parasitic plants:a review. Plant Ecology and Evolution, 2016, 149(1): 5-20. |
| [7] |
Bell TL, Adams MA. Attack on all fronts:functional relationships between aerial and root parasitic plants and their woody hosts and consequences for ecosystems. Tree Physiology, 2011, 31: 3-15. DOI:10.1093/treephys/tpq108 |
| [8] |
李钧敏, 董鸣. 植物寄生对生态系统结构和功能的影响. 生态学报, 2011, 31(4): 1174-1184. |
| [9] |
Donohue K, Dorn L, Griffith C, Kim E, Aguilera A, Polisetty CR, Schmitt J. The evolutionary ecology of seed germination of Arabidopsis thaliana:variable natural selection on germination timing. Evolution, 2005, 59(4): 758-770. DOI:10.1111/evo.2005.59.issue-4 |
| [10] |
Herrera CM, Pellmyr O. Plant animal interactions:an evolutionary approach. Oxford: Blackwell Publishing, 2002.
|
| [11] |
黄双全, 郭友好. 传粉生物学的研究进展. 科学通报, 2000, 45(3): 225-237. DOI:10.3321/j.issn:0023-074X.2000.03.001 |
| [12] |
Matsumura C, Washitani I. Effects of population size and pollinator limitation on seed-set of Primula sieboldii populations in a fragmented landscape. Ecological Research, 2000, 15(3): 307-322. DOI:10.1046/j.1440-1703.2000.00350.x |
| [13] |
Traveset A, Gulias J, Riera N, Mus M. Transition probabilities from pollination to establishment in a rare dioecious shrub species (Rhamnus ludovici-salvatoris) in two habitats. Journal of Ecology, 2003, 91: 427-437. DOI:10.1046/j.1365-2745.2003.00780.x |
| [14] |
肖来云, 普正和. 云南西双版纳桑寄生植物传播与鸟的关系研究. 生态学报, 1994, 14(2): 128-135. DOI:10.3321/j.issn:1000-0933.1994.02.011 |
| [15] |
Boone LS, Fate G, Chang M, Lynn DG. Seed germination.//Press MC, Graves JD eds. Parasitic plants. London: Chapman & Hall, 1995: 14-38.
|
| [16] |
Yoneyama K, Awad AA, Xie X, Yoneyama K, Takeuchi Y. Strigolactones as germination stimulants for root parasitic plants. Plant & Cell Physiology, 2010, 51(7): 1095-1103. |
| [17] |
Hill JP, Edwards W. Dispersal of desiccation-sensitive seeds is not coincident with high rainfall in a seasonal tropical forest in Australia. Biotropica, 2010, 42(3): 271-275. DOI:10.1111/j.1744-7429.2009.00623.x |
| [18] |
张大勇. 植物生活史进化与繁殖生态学. 北京:科学出版社, 2004, 101-127. |
| [19] |
Molau U. Reproductive ecology and biology//Press MC, Graves JD, eds. Parasitic Plants. London: Chapman & Hall, 1995: 141-176.
|
| [20] |
Díaz Infante S, Lara C, Arizmendi MDC, Eguiarte LE, Ornelas JF. Reproductiove ecology and isolation of Psittacanthus calyculatus and P. auriculatus mistletoes (Loranthaceae). PeerJ, 2016.
|
| [21] |
史焱, 马列, 卫莹芳, 王梦月. 小红花寄生的性状、显微和TLC鉴定. 中药材, 2001, 24(2): 93-95. DOI:10.3321/j.issn:1001-4454.2001.02.007 |
| [22] |
中国植物志编辑委员会. 中国植物志(第二十四卷). 北京: 科学出版社, 1988: 115.
|
| [23] |
叶要妹, 张俊卫, 齐迎春, 陈天花, 包满珠. 百日草柱头可授性和花粉活力的研究. 中国农业科学, 2007, 40(10): 2376-2381. DOI:10.3321/j.issn:0578-1752.2007.10.036 |
| [24] |
Dafni A, Maués MM. A rapid and simple procedure to determine stigma receptivity. Sexual Plant Reproduction, 1998, 11: 177-180. DOI:10.1007/s004970050138 |
| [25] |
ISTA. International rules for seed testing. Seed Science and Technology, 1999, 27: 25-30, 35-39. |
| [26] |
Bewley JD, Bradford KJ, Hilhorst HWM, Nonogaki H. Seeds: Physiology of Development, Germination and Dormancy. London: Springer, 2013: 136-140.
|
| [27] |
Barrett SCH. The evolution, maintenance, and the loss of self-incompatibility systems//Lovett Doust J, Lovett Doust L, eds. Plant reproductive ecology: patterns and strategies. Oxford: Oxford University Press, 1988: 98-124.
|
| [28] |
Holsinger KE. Pollination biology and the evolution of mating systems in flowering plants. Evolutionary Biology, 1996, 29: 107-149. |
| [29] |
Rivera G L, Galetto L, Bernardello L. Neactar secrtion patterns, removal effects, and breeding system of Ligaria cuneifoli (Loranthaceae). Canadian Journal of Botany, 1996, 74(12): 1996-2001. DOI:10.1139/b96-239 |
| [30] |
Azpeitia F, Lara C. Reproductive biology and pollination of the parasitic plant Psittacanthus calyculatus (Loranthaceae) in central México. Journal of the Torrey Botanical Society, 2006, 133(3): 429-438. DOI:10.3159/1095-5674(2006)133[429:RBAPOT]2.0.CO;2 |
| [31] |
Darwin C. The effects of cross and self fertilization in the vegetable kingdom. London: John Murray, 1876.
|
| [32] |
Lloyd DG. Some reproductive factors affecting the selection of self-fertilization in plants. American Naturalist, 1979, 113(1): 67-79. DOI:10.1086/283365 |
| [33] |
Lord EM. Cleistogamy:a tool for the study of floral morphogenesis, function and evolution. Botanical Review, 1981, 47(4): 421-429. DOI:10.1007/BF02860538 |
| [34] |
盛晋华, 张雄杰, 刘宏义, 李莉. 寄生植物概述. 生物学通报, 2006, 41(3): 9-13. DOI:10.3969/j.issn.0006-3193.2006.03.004 |
| [35] |
Xiong GS, Li JY, Smith SM. Evolution of Strigolactone perception by seeds of patasitic plants:reinventing the wheel. Molecular Plant, 2016, 9: 493-495. DOI:10.1016/j.molp.2016.01.010 |
| [36] |
Jordano P. Fruits and frugivory//Fenner M, ed. Seeds: The Ecology of Regeneration in Plant Communities. (2nd ed). Wallingford: CAB International, 2000: 125-162.
|
| [37] |
肖来云, 普正和. 西双版纳桑寄生植物的繁殖. 云南植物研究, 1989, 11(2): 175-180. |
| [38] |
罗亚皇, 张玲. 五蕊寄生种子萌发特性. 植物分类与资源学报, 2013, 35(1): 73-80. |
| [39] |
Roberts EH. Predicting the storage life of seeds. Seed Science and Technology, 1973, 1: 499-514. |
| [40] |
唐安军, 龙春林, 刀志灵. 种子顽拗性的形成机理及其保存技术. 西北植物学报, 2004, 24(11): 2170-2176. DOI:10.3321/j.issn:1000-4025.2004.11.037 |
| [41] |
Walters C, Berjak P, Pammenter N, Kenedy K, Raven P. Preservation of recalcitrant seeds. Science, 2013, 339(6122): 915-916. DOI:10.1126/science.1230935 |
| [42] |
胡飞, 孔垂华. 寄生植物对寄主的选择和影响. 应用生态学报, 2004, 15(5): 905-908. DOI:10.3321/j.issn:1001-9332.2004.05.038 |