 2019, Vol. 39
2019, Vol. 39文章信息
- 王家楠, 蒋勇军, 贺秋芳, 范佳鑫, 何瑞亮, 吴超
- WANG Jianan, JIANG Yongjun, HE Qiufang, FAN Jiaxin, HE Ruiliang, WU Chao
- 中梁山岩溶槽谷区荒草地土壤微生物群落对隧道建设的响应
- Response of soil microbial community in grassland to tunnel construction in the karst trough valley, Zhongliang Mountain, Chongqing
- 生态学报. 2019, 39(16): 6136-6145
- Acta Ecologica Sinica. 2019, 39(16): 6136-6145
- http://dx.doi.org/10.5846/stxb201811262560
-
文章历史
- 收稿日期: 2018-11-26
- 修订日期: 2019-05-30

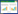




随着国民经济发展和各地交通设施完善, 大型隧道工程建设逐年增多, 给交通发展和经济增长带来新机遇的同时, 也给当地生态、水文和地质环境带来挑战。截至2017年底, 全国修建公路隧道共计16229处、总长度约15285.1 km, 其中大型隧道有902处、长度约4013.2 km[1]。但是, 在隧道开挖和运营过程中, 地下水经隧道漏失, 地下水位下降[2-3], 进而导致地表储水量减少乃至表层泉干涸, 此类环境影响普遍存在于隧道建设区[4]。首尔市隧道建设后, 地下水年排放量6300万m3, 地下水位从1996年的16.85 m下降到2003年的20.40 m[5];重庆市中梁山隧道建设造成29处地表塌陷、48个泉点干枯[6], 地表水受影响区面积约7.2 km2, 严重危及当地居民生活和生产用水[7]。与此同时, 地表、地下水的减少引起当地生态系统供水不足, 土壤性质和植物生长状况随之发生恶化。位于湖南省邵阳市与怀化市交界处的雪峰山隧道建设后, 受地下水漏失的影响, 隧道影响区上覆地表土壤含水率降低, 土壤成分(速效钾、硝态氮、有机质等)含量小于未受隧道影响区, 土壤质量下降[8];重庆中梁山隧道影响区树木年轮宽度持续减少[9], 优势植物个体(马尾松、杉木等)因水分缺失生长速度出现不同程度地衰退[10-11]。此外, 隧道建设对水循环的显著影响也将进一步改变土壤微生物群落结构及其功能。土壤微生物作为土壤生态系统的重要组成部分, 其变化对整个生态系统影响巨大[12], 但是与显著可见的植被变化相比, 土壤微生物群落的变化常不为人知。
土壤微生物在调节土壤元素循环、维持土壤肥力和保障植物养分有效性等方面发挥重要作用[13-14], 对土壤环境的变化较为敏感。土壤水分、营养元素等理化性质和环境因素发生改变时, 通常会导致土壤微生物群落组成和结构发生巨大的变化[15-16]。干旱和半干旱区的土壤微生物群落结构变化研究表明, 土壤含水率变化影响着拟杆菌门和芽单胞菌门的相对丰度[17], 土壤中放线菌门对土壤含水率的响应最快(土壤含水率越低, 放线菌门相对丰度越高), 疣微菌门则相反(土壤含水率越低, 疣微菌门相对丰度越低)[18]。同时土壤含水率的变化常与土壤pH具有协同效应, 土壤pH的升高与土壤微生物群落中Gamma-变形菌门、绿弯菌门和硝化螺旋菌门的相对丰度呈显著正相关, 而与Alpha-变形菌门、装甲菌门和纤维杆菌门呈负相关[19]。土壤含水率降低也可能引起土壤养分(有机质、硝态氮、总磷、有效磷、速效钾等)改变, 从而带来土壤微生物群落结构的变化[14], 如贫营养环境对土壤样品中酸杆菌门具有选择性作用[20]。已有研究表明, 土壤含水率和理化性质的改变将影响土壤微生物群落结构变化, 从而影响土壤物质循环, 扰动地上过程, 引起生态系统功能改变[21]。但是, 隧道建设引起地下水漏失带来的土壤环境变化, 以及土壤环境变化对隧道影响区上覆土壤微生物群落结构带来的影响尚未见详细报道。
由于岩溶区存在地上-地下双层水循环系统[22-23], 土壤厚度薄, 隧道建设后岩溶槽谷区地下水运移管道被打破, 地表水漏失严重, 进而可能会导致土壤理化性质及微生态系统发生改变, 打破当地生态环境平衡。荒草地是陆地生态系统对气候变化反馈的重要组成部分, 在调节土壤碳库的年际变化上发挥重要作用[24], 且荒草地受人为干扰较少, 土壤环境较为稳定, 因此了解荒草地土壤微生物群落对于解释和预测环境变化至关重要。本研究对中梁山龙凤槽谷(隧道影响区)和龙车槽谷(无隧道影响区)的典型荒草地进行采样, 通过对比土壤理化性质(土壤pH、有机质、碱解氮、速效钾、有效磷、全氮、全钾、全磷、容重、土壤含水率)分析隧道建设对土壤环境的影响, 通过16S rDNA高通量测序比较两个槽谷的土壤微生物群落组成差异揭示土壤微生物群落对隧道建设的响应模式, 为岩溶槽谷区土壤微生物多样性提供基础数据, 为减少隧道建设对生态环境的负面效应提供理论参考。
1 研究区概况龙凤、龙车岩溶槽谷区(图 1)位于重庆市北碚区中梁山观音峡背斜(29°39′—29°49′N, 106°22′—106°29′E), 发育于三叠纪嘉陵江组的岩溶角砾岩白云质灰岩中, 属于川东平行岭谷华蓥山帚状褶皱束南沿部分, 呈现出“一山三岭两槽”的“笔架”式地形。背斜核部地层为三叠系下统飞仙关组(Tlf), 向两翼逐渐过渡为三叠系下统嘉陵江组(T1j)、三叠系中统雷口坡组(T2l)和三叠系上统须家河组(T3xj)。槽谷区总面积38 km2, 受亚热带季风影响, 年均温18℃, 年均降雨量1200 mm, 土壤主要为岩溶区典型棕黄壤, 土层厚薄不均。

|
| 图 1 重庆中梁山地质简图和采样点分布图 Fig. 1 Geological profile and sampling sites distribution at Zhongliang Mountain, Chongqing |
1999—2018年轻轨和重庆内、外环高速建设期间, 三条大型隧道穿越观音峡背斜连通重庆北部区县:轨道交通6号线中梁山隧道(2010年10月开工—2013年8月贯通)长度为4554 m;G75渝武高速北碚隧道(1999年6月开工—2001年12月贯通)左洞长4025.5 m、右洞长4045 m;G5001绕城高速施家梁隧道(2006年4月开工—2008年6月贯通)左洞长4303 m、右洞长4267.5 m。三条隧道通过雷口坡组、嘉陵江组、飞仙关组灰岩时地下水涌出, 并从隧道口排出地表, 影响岩溶含水层。其中, 龙凤槽谷区总面积约11.7 km2, 形成一个相对独立的水文地质单元, 三条大型隧道从其下穿过, 地下水文流场受隧道建设影响, 出现泉眼断流、水井水位逐渐降低甚至完全消失等现象;而南部另一个相对独立的水文地质单元即龙车槽谷, 总面积约26.8 km2, 作为对照区, 不受隧道建设的影响。
2 研究方法 2.1 样品采集本研究于2017年11月在龙凤槽谷区选取6个隧道影响区典型荒草地样地(T1—T6), 在龙车槽谷区对照选取6个无隧道影响区典型荒草地样地(U7—U12)(图 1)。使用梅花5点法分两层(0—20 cm、20—40 cm)采集土壤样品, 其中隧道影响区和无隧道影响区土壤样品各11个(T1、U7缺失20—40 cm深度的土壤样品)。各样点采集土样三份, 剔除根系和石块等杂质, 一份储藏在冰箱(-20℃)中用作土壤DNA的提取;一份测试土壤容重;一份土样用来测试土壤含水率(带回实验室后24小时内测定), 余下的土样放置在阴凉干燥的地方自然风干后测定土壤理化性质。
2.2 室内实验(1) 土壤理化性质测定
土壤容重测定采用环刀法, 土壤含水率测定采用烘干法, 测定在西南大学地理科学学院完成。余下土样风干后, 混合均匀研磨土样后过0.25 mm和2 mm筛。土壤pH采用电极法测试;土壤有机质采用重铬酸钾氧化-外加热法;土壤碱解氮采用碱解扩散法;土壤速效钾采用醋酸铵浸提-火焰光度法测定;土壤有效磷采用0.5 mol/L的NaHCO3-浸提钼锑抗比色法测定;土壤全氮采用半微量凯氏法测定;土壤全钾采用氢氧化钠熔融-火焰光度法测定;土壤全磷采用氢氧化钠熔融-钼锑抗比色法测定, 以上测定在西南大学资源环境学院完成。
(2) 土壤微生物样品16S rDNA高通量测序
另一份土壤样品自然解冻后称取0.5 g, 使用FastDNA® SPIN Kit for Soil试剂盒(MP Biomedicals, Santa Ana, CA, USA)提取DNA。测序使用illuminaMiseq平台, 利用细菌通用引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)测定V3—V4区域。PCR正式试验采用TransGen AP221-02:TransStartFastpfu DNA Polymerase, 20 μL反应体系。PCR扩增条件为:95℃ 3 min;95℃ 30 s, 55℃ 30 s, 72℃ 45 s, 27个循环;72℃ 10 min。全部样本按照正式实验条件进行, 将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测, 使用AxyPrepDNA凝胶回收试剂盒(AXYGE公司)切胶回收PCR产物, Tris_HCl洗脱;2%琼脂糖电泳检测。参照电泳初步定量结果, 将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统(Promega公司)进行检测定量, 之后按照每个样本的测序量要求, 进行相应比例的混合。将相似性97%以上的DNA序列聚类(cluster)归为许多小组, 一个小组为一个OTU, 对97%相似水平下的OTU进行生物信息统计分析。土壤总DNA提取在西南大学地理科学学院岩溶环境重庆市重点实验室完成, 16S rDNA高通量测序送样至上海美吉生物医药科技有限公司完成。
2.3 数据分析通过统计分析软件SPSS 24.0进行t检验, 对比分析隧道影响区、无隧道影响区土壤理化性质差异。土壤微生物群落Alpha多样性选用Sobs指数和香农维纳指数(Shannon Wiener)进行估算, Sobs是用来估算样品中所含OTU数目的指数, 在生态学中常用来统计物种总数;香农维纳指数反映样品中微生物多样性, Shannon值越大, 说明群落多样性越高。土壤微生物群落Beta多样性选用PCoA分析(Principal co-ordinates analysis)即主坐标分析, 用来研究样本群落组成的相似性或差异性。Wilcoxon检验用于分析两组样品中微生物群落结构差异。通过VIF分析(Variance inflation factor, 方差膨胀因子)进行环境因子筛选, 多次过滤掉VIF大于10、保留自相关作用较小的环境因子后进行RDA/CCA分析, 进而检测环境因子、样本、菌群三者间的关系或者两两之间的关系。数据分析在上海美吉公司Majorbio高通量数据分析云平台完成, 利用Adobe Illustrator CS6进行绘制作图。
3 结果与分析 3.1 荒草地土壤理化性质分析如表 1所示, 隧道影响区荒草地土壤pH、容重和有效磷大于无隧道影响区, 两组样品中土壤pH存在显著性差异(P < 0.05)。而无隧道影响区土壤含水率、碱解氮、全氮、有机质、全磷、全钾和速效钾大于隧道影响区, 两组样品之间土壤含水率(P < 0.05)、全钾(P < 0.05)、速效钾(P=0.001)、全磷(P < 0.01)存在显著差异。
| 统计项目 Statistical item | 隧道影响区 Tunneling affected valley | 无隧道影响区 Tunneling unaffected valley | 显著性水平 P value |
| 土壤含水率Soil water content/% | 24.22±3.33 | 30.72±6.61 | 0.015 |
| pH | 7.90±0.29 | 7.49±0.64 | 0.026 |
| 有机质Organic matter/(g/kg) | 19.00±7.82 | 20.61±10.09 | 0.644 |
| 容重Bulk density/(g/cm3) | 1.33±0.08 | 1.25±0.06 | 0.090 |
| 全氮Total nitrogen/(mg/kg) | 1416.04±406.50 | 1496.27±489.10 | 0.576 |
| 碱解氮Alkali-hydrolyzable nitrogen/(mg/kg) | 153.02±63.99 | 157.98±101.53 | 0.875 |
| 全磷Total phosphorus/(mg/kg) | 477.51±163.34 | 581.51±211.70 | 0.007 |
| 有效磷Available phosphorus/(mg/kg) | 12.87±12.42 | 9.25±7.56 | 0.120 |
| 全钾Total potassium/(g/kg) | 29.12±3.15 | 32.53±3.50 | 0.013 |
| 速效钾Available potassium/(mg/kg) | 112.45±17.82 | 151.09±44.13 | 0.001 |
对样本的16S rDNA测序数据进行随机抽平, 消除由于样本间总序列数不等而造成的群落结构差异(图 2)。采用抽样序列数与它们所代表的OTU数目构建稀疏曲线, 可以看出稀疏曲线均趋于平缓, 表明测序数据合理。两组样品的文库覆盖度均大于98%, 表明本次测序结果获得的序列能够代表不同采样点土壤中的绝大多数微生物, 可以反映样本中微生物的真实情况。微生物群落多样性指数结果表明, 隧道影响区物种丰度Sobs指数为1610, 多样性Shannon指数为6.35;无隧道影响区物种丰度Sobs指数为1241, 多样性Shannon指数为5.74, 由此隧道影响区土壤微生物群落的物种丰度和多样性高于无隧道影响区。

|
| 图 2 重庆中梁山土壤微生物群落多样性指数 Fig. 2 Soil microbial community diversity index of Zhongliang Mountain, Chongqing |
22个样品的测序数据聚类后共得到属于43个门类、167个纲类、318个目类、491个科类、685个属类、731个种类和8566条OTU。通过对比两组样品中相对丰度大于0.5%的物种(图 3), 发现门水平上土壤微生物群落的组成及优势类群大致相同, 但其相对丰度在两组样品间却存在显著差异。变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、拟杆菌门(Bacteriodetes)、硝化螺旋菌门(Nitrospirae)、芽单胞菌门(Gemmatimonadetes)7个主要门在总OTU数中的占比超过90%。其中在隧道影响区土壤样品中变形菌门(Proteobacteria)相对丰度最多, 所占比例为26.56%, 其次为酸杆菌门(Acidobacteria), 所占比例为25.46%;无隧道影响区土壤样品中变形菌门(Proteobacteria)相对丰度最多, 所占比例为27.51%, 酸杆菌门(Acidobacteria)次之, 所占比例为20.77%。

|
| 图 3 中梁山荒草地样品微生物群落组成 Fig. 3 Microbial community composition of grassland samples at Zhongliang Mountain |
土壤样品所获得的序列约50%能进一步分类到属水平上, 其中17个属相对丰度超过1%(图 3)。对17个属进行归类, 依次为:norank_f__Syntrophobacteraceae、Kaistobacter、norank_o__MND1、norank_f__Rhodospirillaceae、unclassified_f__Oxalobacteraceae属于变形菌门(Proteobacteria), norank_o__iii1-15、norank_o__RB41、unclassified_c__Acidobacteria-6属于酸杆菌门(Acidobacteria), norank_c__S085、norank_f__A4b、norank_f__[Kouleothrixaceae]属于绿弯菌门(Chloroflexi), norank_c__Gemmatimonadetes、norank_c__Gemm-1属于芽单胞菌门(Gemmatimonadetes), norank_f__0319-6A21属于硝化螺旋菌门(Nitrospirae), norank_f__Gaiellaceae属于放线菌门(Actinobacteria), norank_f__Chitinophagaceae属于拟杆菌门(Bacteriodetes), norank_p__WPS-2属于WPS-2。norank_o__iii1-15、norank_o__RB41、norank_f__A4b、norank_f__0319-6A21为相对丰度最多的属, 在隧道影响区和无隧道影响区样品平均相对丰度所占比例为3%以上。
Alpha多样性(物种丰度Sobs指数、多样性Shannon指数)反映样品中物种的总丰富度以及分布均匀性[25];Beta群落多样性则反映生境间的物种差异, 体现了生态因子对生境中物种多样性的影响[26]。PCoA分析表明, 隧道影响区和无隧道影响区土壤样品Beta多样性在属水平存在十分显著的差异(图 4), 两组样品分离度较大, 明显划分为不同的群落。ANOSIM分析表明, 研究区两组样品群落多样性存在显著差异(P=0.001, 图 4)。隧道影响区土壤样品的组间差异大于组内差异, 说明隧道建设对土壤性质的改变, 可能引起了土壤中细菌群落结构的变化。

|
| 图 4 荒草地样品Beta多样性分析和样本分组分析 Fig. 4 Beta diversity analysis and ANOSIM analysis of grassland samples |
根据Wilcoxon检验组间差异显著性, 发现两组样品微生物群落中, 属水平上三种表现出丰度差异显著的物种为酸杆菌门(Acidobacteria)中norank_o__iii1-15(P < 0.01)、芽单胞菌门(Gemmatimonadetes)中norank_c__Gemmatimonadetes(P < 0.001)和变形菌门(Proteobacteria)中的norank_o__MND1(P < 0.05), 其土壤中的相对丰度表现为隧道影响区显著高于无隧道影响区(图 5)。其中norank_o__iii1-15、norank_o__MND1和norank_c__Gemmatimonadetes在隧道影响区相对丰度分别为:7.78%、3.10%和2.26%, 在无隧道影响区相对丰度分别为:4.00%、1.35%和0.38%。

|
| 图 5 土壤微生物群落物种差异分析 Fig. 5 Analysis of species differences in soil microbial communities on genus level |
VIF方差膨胀因子筛选发现, pH、土壤含水率、全氮、全钾、容重和有效磷为不存在多重共线性的环境因子(0 < VIF < 10)。环境因子与样品微生物群落结构数据RDA/CCA分析表明(图 6), 隧道影响区荒草地土壤微生物群落与pH(P=0.001)呈显著的正相关关系, 无隧道影响区荒草地土壤微生物群落与土壤含水率(P=0.006)呈显著的正相关关系, 全氮(P=0.001)则与大部分样品中土壤微生物群落呈负相关关系。隧道影响区荒草地土壤微生物群落与有效磷、容重呈正相关关系, 无隧道影响区荒草地土壤微生物群落与全钾呈正相关关系, 但均无显著性差异。隧道影响区样品丰度较高的代表性物种依次为norank_o__iii1-15、norank_o__RB41和norank_f__A4b, 在菌群与环境因子之间关系上显示出norank_o__iii1-15与pH、有效磷、容重呈正相关关系, 与全氮、土壤含水率呈负相关关系;norank_o__RB41与全氮、有效磷、容重呈正相关关系, 与土壤含水率、全钾呈负相关关系;norank_f__A4b与pH、有效磷、容重呈正相关关系, 与全氮呈负相关关系。

|
| 图 6 土壤微生物群落与环境因子RDA分析 Fig. 6 RDA analysis results showing the relationship between environmental factors and the soil microbial community |
中梁山隧道开挖后, 地下水以基岩裂隙水和构造裂隙水的形式涌出, 深层溶隙水从渗流转为明流从而加大了深层地下水的排泄量[6], 地下水平衡被打破, 引起隧道影响地区土壤含水率下降。中梁山岩溶槽谷区基岩造壤能力低, 土壤持水性能差, 且当地岩溶十分发育形成地上地下的双层排水系统[27], 地表水极易向地下流失。当地下水位下降时, 地表水和土壤水作为水循环的重要环节, 流经表层岩溶带、岩溶裂隙/孔隙向深部基岩补给[6], 造成地表径流和表层泉干涸。中梁山岩溶槽谷区三条隧道贯通后, 深层地下水排泄漏失, 岩溶裂隙发育形成的地下排水管网放大了隧道建设引起的地下水漏失, 引起地下水位下降和上覆土壤水分下降, 从而导致隧道影响区土壤含水率(均值)显著降低(表 1, 隧道影响区24% < 无隧道影响区31%)。此外, 隧道影响区土壤容重较大, 表明土壤紧实, 反映了土壤的水分、空气、热量状况差于无隧道影响区土壤。
土壤含水率是影响土壤微生物的关键因素[28-30], 因而地下水减少对土壤微生物的潜在影响被列为隧道建设的主要环境效应之一, 影响着土壤微生物多样性和群落结构。隧道影响区荒草地土壤水分状况年际变化幅度大, 湿度条件弹性大, 生态环境较为波动, 可能会使潜在的机会性微生物种群快速生长和适应, 从而缓和微生物群落的种内竞争, 促进生态系统中土壤微生物物种的共存[31]。因此, 与土壤含水率较高的无隧道影响区相比, 土壤微生物在隧道影响区低含水率条件下表现出更高的丰度和多样性。同时, 在隧道影响区代表性微生物类群中, norank_o__iii1-15(属于酸杆菌门)对干旱的环境条件具有很强的抵抗力[32], 更能适应隧道区土壤含水率较低的环境成为优势物种;norank_c__Gemmatimonadetes(属于芽单胞菌门)为干旱土壤中的优势菌群[17, 33], 在隧道影响区土壤中含量更多。因此, 以norank_o__iii1-15(属于酸杆菌门)和norank_c__Gemmatimonadetes(属于芽单胞菌门)为特征物种的隧道影响区土壤微生物群落, 其群落结构变化受下降的土壤含水率控制, 微生物菌群与土壤含水率呈显著负相关关系(P < 0.05)。
4.2 隧道影响区土壤微生物优势菌群对土壤pH和养分变化的响应土壤水分是土壤中营养元素运移和吸收的载体, 其波动变化将导致土壤pH、有效养分等因子的协同变化。中梁山隧道建设引起的地下水排泄和土壤含水率下降, 造成了受隧道影响地区上覆土壤pH上升、土壤有效养分下降等一系列土壤环境的变化。相较于无隧道影响区, 隧道影响区土壤含水率下降, 土壤中水解酸的溶出减少, H+的释放速率减缓[34], 使隧道影响区土壤pH高于无隧道影响区(P < 0.05)。同时, 地下水位的下降也引起溶解在土壤水中的养分随之下降, 加上大气降雨的淋溶作用, 土壤中的养分被淋溶到土壤下层甚至到地下水, 从而造成土壤中有机质、碱解氮、速效钾等组分的减少;同时土壤含水率下降, 土壤水分运移路径缩短, 随土壤水分运移的营养物质减少, 土壤微环境中的营养物质补充减少, 由此隧道影响区成为贫营养微环境。
受隧道影响区土壤中, pH上升、有效养分降低给个体微生物和整个群落带来生理压力, 显著改变生态系统的功能, 引起土壤中微生物群落结构的改变。在隧道影响区代表性微生物类群中, norank_c__Gemmatimonadetes的相对丰度与土壤pH呈显著正相关, 即相对丰度随土壤pH值的增加而增加[35];norank_o__RB41和norank_o__iii1-15为酸杆菌门中第4和第6亚门细菌, 其对pH的适应范围为pH < 10, 更适应较高pH的隧道影响区土壤[19, 36];norank_o__MND1属于Beta-变形菌, 丰度与pH呈正相关[37]。因此隧道影响区荒草地土壤中的优势微生物菌群更适应高pH的土壤环境。芽单胞菌门相对丰度与土壤全氮、土壤全磷呈显著负相关, 即相对丰度随土壤全氮、土壤全磷的增加而降低[35];酸杆菌门是土壤中最常见微生物门类之一, 在土壤生态过程中起到重要的作用, 具有寡营养、难培养等特点[20];norank_o__MND1属于Beta-变形菌, 其降解能力可变, 与营养物质呈负相关, 适于贫营养环境[38]。隧道影响区土壤养分较缺乏的环境条件对微生物群落的环境压力, 造成适应贫营养环境微生物类群在隧道影响区荒草地土壤中更占优势。
5 结论(1) 中梁山岩溶槽谷荒草地土壤样品中, 酸杆菌门的norank_o__iii1-15和norank_o__RB41、绿弯菌门的norank_f__A4b、硝化螺旋菌门norank_f__0319-6A21为相对丰度最多的细菌属, 平均相对丰度占全部OTU的3%以上。
(2) 隧道建设后地下水漏失严重, 土壤含水率减少, 导致土壤pH增加, 土壤有效养分下降。隧道影响区荒草地土壤微生物群落与pH呈正相关关系, 而与土壤含水率、土壤有效养分呈负相关关系。能适应低含水率、高pH和贫营养环境的norank_o__iii1-15、norank_c__Gemmatimonadetes和norank_o__MND1成为隧道影响区荒草地土壤微生物群落的优势菌群。
| [1] |
陈建勋, 罗彦斌, 万利, 刘同展. 超大跨度公路隧道研究现状与面临的挑战. 筑路机械与施工机械化, 2018, 35(6): 34-44. |
| [2] |
Vincenzi V, Gargini A, Goldscheider N. Using tracer tests and hydrological observations to evaluate effects of tunnel drainage on groundwater and surface waters in the Northern Apennines (Italy). Hydrogeology Journal, 2009, 17(1): 135-150. DOI:10.1007/s10040-008-0371-5 |
| [3] |
Yoo C. Interaction between tunneling and groundwater-numerical investigation using three dimensional stress-pore pressure coupled analysis. Journal of Geotechnical and Geoenvironmental Engineering, 2005, 131(2): 240-250. DOI:10.1061/(ASCE)1090-0241(2005)131:2(240) |
| [4] |
Zhao Y, Li P F, Tian S M. Prevention and treatment technologies of railway tunnel water inrush and mud gushing in China. Journal of Rock Mechanics and Geotechnical Engineering, 2013, 5(6): 468-477. DOI:10.1016/j.jrmge.2013.07.009 |
| [5] |
Chae G T, Yun S T, Choi B Y, Yu S Y, Jo H Y, Mayer B, Kim Y J, Lee J Y. Hydrochemistry of urban groundwater, Seoul, Korea:the impact of subway tunnels on groundwater quality. Journal of Contaminant Hydrology, 2008, 101(1/4): 42-52. |
| [6] |
罗鉴银, 傅瓦利. 岩溶地区开凿隧道对地下水循环系统的破坏——以重庆市中梁山为例. 西南农业大学学报:自然科学版, 2005, 27(4): 432-435. |
| [7] |
Cheng P, Zhao L H, Luo Z B, Li L, Li Q, Deng X, Peng W Q. Analytical solution for the limiting drainage of a mountain tunnel based on area-well theory. Tunnelling and Underground Space Technology, 2019, 84: 22-30. DOI:10.1016/j.tust.2018.10.014 |
| [8] |
杨长健.雪峰山隧道隧址区地下水土壤环境影响评价研究[D].长沙: 中南大学, 2007. http://cdmd.cnki.com.cn/Article/CDMD-10533-2007171318.htm
|
| [9] |
Zheng W, Wang X L, Tang Y, Liu H, Wang M, Zhang L J. Use of tree rings as indicator for groundwater level drawdown caused by tunnel excavation in Zhongliang Mountains, Chongqing, Southwest China. Environmental Earth Sciences, 2017, 76(15): 522. DOI:10.1007/s12665-017-6859-3 |
| [10] |
王芳其, 郑炜, 徐华, 陈剑楠. 岩溶山区隧道地下水漏失对植物生长的影响分析及对策. 隧道建设(中英文), 2018, 38(6): 915-923. |
| [11] |
郑炜, 杨保, 徐宗永, 肖羚. 隧道建设活动对地表植被群落动态的影响——以武昆高速公路象鼻岭隧道为例. 公路交通技术, 2012(5): 145-149. DOI:10.3969/j.issn.1009-6477.2012.05.033 |
| [12] |
Wang X L, Yu S Q, Zhou L X, Fu S L. Soil microbial characteristics and the influencing factors in subtropical forests. Acta Ecologica Sinica, 2016, 36(1): 8-15. DOI:10.1016/j.chnaes.2015.12.004 |
| [13] |
Xu X F, Thornton P E, Post W M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecology and Biogeography, 2013, 22(6): 737-749. DOI:10.1111/geb.12029 |
| [14] |
Jiao S, Chen W M, Wang J L, Du N N, Li Q P, Wei G H. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome, 2018, 6(1): 146. DOI:10.1186/s40168-018-0526-0 |
| [15] |
Žifčáková L, Větrovský T, Howe A, Baldrian P. Microbial activity in forest soil reflects the changes in ecosystem properties between summer and winter. Environmental Microbiology, 2016, 18(1): 288-301. DOI:10.1111/1462-2920.13026 |
| [16] |
Fang X M, Yu D P, Zhou W M, Zhou L, Dai L M. The effects of forest type on soil microbial activity in Changbai Mountain, Northeast China. Annals of Forest Science, 2016, 73(2): 473-482. DOI:10.1007/s13595-016-0540-y |
| [17] |
Bastida F, Torres I F, Andrés-Abellán M, Baldrian P, López-Mondéjar R, Větrovský T, Richnow H H, Starke R, Ondoño S, García C, López-Serrano F R, Jehmlich N. Differential sensitivity of total and active soil microbial communities to drought and forest management. Global Change Biology, 2017, 23(10): 4185-4203. DOI:10.1111/gcb.13790 |
| [18] |
Barnard R L, Osborne C A, Firestone M K. Responses of soil bacterial and fungal communities to extreme desiccation and rewetting. The ISME Journal, 2013, 7(11): 2229-2241. DOI:10.1038/ismej.2013.104 |
| [19] |
Liu J J, Sui Y Y, Yu Z H, Shi Y, Chu H Y, Jin J, Liu X B, Wang G H. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biology and Biochemistry, 2014, 70: 113-122. DOI:10.1016/j.soilbio.2013.12.014 |
| [20] |
Lee S H, Ka J O, Cho J C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiology Letters, 2008, 285(2): 263-269. DOI:10.1111/j.1574-6968.2008.01232.x |
| [21] |
Bell C, McIntyre N, Cox S, Tissue D, Zak J. Soil microbial responses to temporal variations of moisture and temperature in a Chihuahuan desert grassland. Microbial Ecology, 2008, 56(1): 153-167. |
| [22] |
Bakalowicz M. Karst groundwater:a challenge for new resources. Hydrogeology Journal, 2005, 13(1): 148-160. DOI:10.1007/s10040-004-0402-9 |
| [23] |
Gu D X, Zhang Z F, Mallik A, Zhou A P, Mo L, He C X, Huang Y Q. Seasonal water use strategy of Cyclobalanopsis glauca in a karst area of southern China. Environmental Earth Sciences, 2015, 74(2): 1007-1014. DOI:10.1007/s12665-014-3817-1 |
| [24] |
Ochoa-Hueso R, Collins S L, Delgado-Baquerizo M, Hamonts K, Pockman W T, Sinsabaugh R L, Smith M D, Knapp A K, Power S A. Drought consistently alters the composition of soil fungal and bacterial communities in grasslands from two continents. Global Change Biology, 2018, 24(7): 2818-2827. DOI:10.1111/gcb.14113 |
| [25] |
Engelhardt I C, Welty A, Blazewicz S J, Bru D, Rouard N, Breuil M C, Gessler A, Galiano L, Miranda J C, Spor A, Barnard R L. Depth matters:effects of precipitation regime on soil microbial activity upon rewetting of a plant-soil system. The ISME Journal, 2018, 12(4): 1061-1071. DOI:10.1038/s41396-018-0079-z |
| [26] |
Hartmann M, Frey B, Mayer J, Mäder P, Widmer F. Distinct soil microbial diversity under long-term organic and conventional farming. The ISME Journal, 2015, 9(5): 1177-1194. DOI:10.1038/ismej.2014.210 |
| [27] |
张川, 陈洪松, 张伟, 聂云鹏, 叶莹莹, 王克林. 喀斯特坡面表层土壤含水量、容重和饱和导水率的空间变异特征. 应用生态学报, 2014, 25(6): 1585-1591. |
| [28] |
Xue L, Ren H D, Li S, Leng X H, Yao X H. Soil bacterial community structure and co-occurrence pattern during vegetation restoration in karst rocky desertification area. Frontiers in Microbiology, 2017, 8: 2377. DOI:10.3389/fmicb.2017.02377 |
| [29] |
Evans S E, Wallenstein M D. Climate change alters ecological strategies of soil bacteria. Ecology Letters, 2014, 17(2): 155-164. DOI:10.1111/ele.12206 |
| [30] |
Cregger M A, Schadt C W, McDowell N G, Pockman W T, Classen A T. Response of the soil microbial community to changes in precipitation in a semiarid ecosystem. Applied and Environmental Microbiology, 2012, 78(24): 8587-8594. DOI:10.1128/AEM.02050-12 |
| [31] |
Bell C W, Tissue D T, Loik M E, Wallenstein M D, Acosta-Martinez V, Erickson R A, Zak J C. Soil microbial and nutrient responses to 7 years of seasonally altered precipitation in a Chihuahuan Desert grassland. Global Change Biology, 2014, 20(5): 1657-1673. DOI:10.1111/gcb.12418 |
| [32] |
Ren C J, Chen J, Lu X J, Doughty R, Zhao F Z, Zhong Z K, Han X H, Yang G H, Feng Y Z, Ren G X. Responses of soil total microbial biomass and community compositions to rainfall reductions. Soil Biology and Biochemistry, 2018, 116: 4-10. DOI:10.1016/j.soilbio.2017.09.028 |
| [33] |
DeBruyn J M, Nixon L T, Fawaz M N, Johnson A M, Radosevich M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Applied and Environmental Microbiology, 2011, 77(17): 6295-6300. DOI:10.1128/AEM.05005-11 |
| [34] |
袁巧霞, 朱端卫, 武雅娟. 温度、水分和施氮量对温室土壤pH及电导率的耦合作用. 应用生态学报, 2009, 20(5): 1112-1117. |
| [35] |
李飞.高寒湿地和草甸退化与恢复对土壤养分、土壤微生物及有机化合物的影响[D].兰州: 兰州大学, 2018. http://cdmd.cnki.com.cn/Article/CDMD-10730-1018979822.htm
|
| [36] |
Xiong J B, Liu Y Q, Lin X G, Zhang H Y, Zeng J, Hou J Z, Yang Y P, Yao T D, Knight R, Chu H Y. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environmental Microbiology, 2012, 14(9): 2457-2466. DOI:10.1111/j.1462-2920.2012.02799.x |
| [37] |
Kim J M, Roh A S, Choi S C, Kim E J, Choi M T, Ahn B K, Kim S K, Lee Y H, Joa J H, Kang S S, Lee S A, Ahn J H, Song J, Weon H Y. Soil pH and electrical conductivity are key edaphic factors shaping bacterial communities of greenhouse soils in Korea. Journal of Microbiology, 2016, 54(12): 838-845. DOI:10.1007/s12275-016-6526-5 |
| [38] |
肖汉玉, 王鹏, 刘君政, 李传琼, 李燕, 赵君. 基于高通量测序的乐安江冬季细菌群落特征分析. 微生物学报, 2019, 59(1): 181-194. |