 2018, Vol. 38
2018, Vol. 38文章信息
- 毛晋花, 邢亚娟, 闫国永, 王庆贵.
- MAO Jinhua, XING Yajuan, YAN Guoyong, WANG Qinggui.
- 陆生植物生物量分配对模拟氮沉降响应的Meta分析
- A meta-analysis of the response of terrestrial plant biomass allocation to simulated N deposition
- 生态学报. 2018, 38(9): 3183-3194
- Acta Ecologica Sinica. 2018, 38(9): 3183-3194
- http://dx.doi.org/10.5846/stxb201706281164
-
文章历史
- 收稿日期: 2017-06-28
- 网络出版日期: 2018-01-26

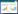




2. 黑龙江省林业科学研究所, 哈尔滨 150081
2. Institute of Forestry Science of Heilongjiang Province, Harbin 150081, China
自工业革命以来, 由于化石燃料的燃烧及化肥的使用, 大气氮沉降明显加重, 预计全球平均氮沉降量到21世纪中期将达到200 Tg N a-1, 约为现在氮沉降量的二倍左右[1], 氮沉降会对陆地生态系统产生严重影响[2-3], 有关氮沉降的模拟研究已经成为全球气候变化的热点领域。氮沉降增加会提高氮的可利用性, 进而改变土壤的养分状况, 影响植物各组织中光合作用产物的数量和分配[4-5]。有研究表明在养分贫瘠的生境中, 植物会分配更多的生物量给根部, 而当养分充足时地上生物量更大, 这符合功能平衡假说[6]。植物会通过改变生物量分配模式来适应外界环境的变化[7-9], 目前虽然有大量氮沉降背景下植物生物量分配的相关研究[10-11], 但氮沉降对生物量在植物各组织中具体分配的影响尚不清楚, 需要加强该领域研究。
根、茎、叶等器官在植物生活史中均发挥至关重要的作用, 它们共同调节植物的生存、生长和繁殖[12]。资源分配在调节植物生长、发育、繁殖和抗性等方面起着重要作用, 同时也是陆地生态系统碳、氮循环的重要影响因素。陆生植物生物量在地上和地下的分配策略是为适应不同环境所采取的重要措施, 植物根冠间生物量的分配模式是长期适应自然选择的结果, 根冠比在一定程度上可反应某种植物在陆地环境中生殖隔离的进化史[13-14], 定量了解植物的生物量分配模式对生态学研究具有重要意义。近年来有大量研究探索植物地上、地下生物量和根冠比对环境变化的响应[15-18], 但是不同研究所得出的结论不尽相同, 因此, 试图得出植物生物量分配的一般性规律, 有必要对全球范围内相关研究的试验数据进行整合分析。
本研究收集了国内外有关大气氮沉降对植物生物量分配影响的野外模拟试验观测数据, 通过Meta分析定量评估了氮沉降处理对植物生物量分配的影响, 并通过亚组分析阐明了不同生态系统类型、植物种类、氮肥形式、施氮水平及持续时间影响下的植物地上、地下生物量、根冠比和总生物量4个指标对外源氮输入的响应, 探索了氮沉降对植物生物量分配的影响规律, 为未来大气氮沉降加重情景下植物生物量分配的相关研究提供参考依据。
1 材料与方法 1.1 数据的收集及纳入标准通过对Web of Science和China National Knowledge Infrastructure(CNKI)等数据库的检索, 本文利用关键词“氮沉降、氮添加、氮肥和生物量分配等”共收集到63篇同行发表的论文[15, 19-80], 共562个观测值用于Meta分析。所收集的论文必须符合以下标准:(1)所选论文中必须包含至少一个用于分析植物生物量分配所选定的指标, 而且文章中对试验持续时间、施氮量、种类以及试验地的气候类型、生态系统类型均有明确说明; (2)为考虑施氮条件下气候条件和生态系统类型对生物量分配的影响, 仅仅野外模拟氮沉降的试验被考虑在内, 室内试验被排除; (3)各独立研究中试验区和对照区要处于同样的气候条件下; (4)各指标的均值、方差或标准差均在文章中有说明或可通过软件Engauge Digitizer (Free Software Foundation, Inc., Boston, MA, USA)从各研究的图表中进行提取或可通过计算得出; (5)对于交互作用的研究, 仅提取模拟氮沉降的实验组和对照组数据。为了避免发表偏倚, 本文通过Stata12(Stata Corp., College Station, TX, USA)的Egger直线回归法对各数据进行检验使数据来源更可靠。
在数据库中通过地上生物量(AGB)、地下生物量(BGB)、根冠比(R/S)、总生物量(TB)、叶生物量(LB)、茎生物量(SB)、细根生物量(FRB)、粗根生物量(CRB)、茎重比(SWR)、根重比(RWR)和叶重比(LWR)这11个指标(表 1)来说明氮沉降条件下植物生物量的分配情况, 并将地上、地下生物量、根冠比和总生物量分成了不同亚组用于进一步探讨不同因素对氮沉降引起生物量分配变化的相对影响。具体分组情况如下:氮处理水平分为高氮(HN)≥120 kg hm-2 a-1, 中氮(MN)60—120 kg hm-2 a-1和低氮(LN)≤60 kg hm-2 a-1; 氮添加形式分为硝酸铵(NH4NO3)、尿素(urea)和其他; 生态系统类型分为森林(forest)、草原(grassland)和其他; 植物种类分为草本(grass)、阔叶树(broadleaf)、针叶树(conifer)、灌木(shrub)和其他; 研究持续时间分为不超过1年、1—3年和3年及以上3个阶段。
| 缩写 Abbreviation |
英文全拼 English full name |
指标 Variables |
定义 Definition |
单位 Unit |
| AGB | Aboveground biomass | 地上生物量 | 植物叶和茎的干重 | g |
| BGB | Belowground biomass | 地下生物量 | 植物根的干重 | g |
| R/S | Root/shoot ratio | 根冠比 | 根干重/叶干重+茎干重 | g/g |
| TB | Total biomass | 总生物量 | 根、茎和叶的总干重 | g |
| LB | Leaf biomass | 叶生物量 | 叶干重 | g |
| SB | Stem biomass | 茎生物量 | 茎干重 | g |
| FRB | Fine root biomass | 细根生物量 | < =2 mm根的干重 | g |
| CRB | Coarse root biomass | 粗根生物量 | 2 mm以上根的干重 | g |
| SWR | Stem weight ratio | 茎重比 | 茎干重/根茎叶的总干重 | g/g |
| RWR | Root weight ratio | 根重比 | 根干重/根茎叶的总干重 | g/g |
| LWR | Leaf weight ratio | 叶重比 | 叶干重/根茎叶的总干重 | g/g |
Meta分析通过METAWIN 2.1(Sinauer Associates Inc., Sunderland, MA, USA)和Stata 12(Stata Corp., College Station, TX, USA)实现, 通过响应比(RR)的自然对数作为效应量(effect size)来衡量氮沉降对植物生物量分配的影响, 公式如下:
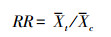
|
(1) |
式中, Xt和Xc分别表示试验组和对照组的均值。
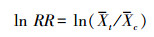
|
(2) |
方差为:
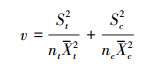
|
(3) |
式中, St和Sc分别代表试验组和对照组的标准差, nt和nc分别代表试验组和对照组的样本量。
权重w由公式(4)计算得出。由于某些研究中的观测值个数要大于1, 因此将权重调整为每个研究中的观测值总数, 通过总权重w′(公式(5))来估计总效应量(RR++, 公式(6、7))。
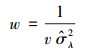
|
(4) |

|
(5) |

|
(6) |
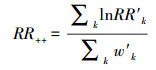
|
(7) |
式中, n是每项研究中观测值的总个数, w′k和lnRR′k是第k个观测值所对应的w′和lnRR′。
本研究采取随机效应模型进行Meta分析, 当某一指标平均效应量的95%置信区间和“0”没有交叉时, 表示氮沉降对该指标有显著影响, 即认为氮沉降对该指标的影响具有统计学意义, 当效应量大于0时表示氮沉降对指标产生正效应, 相反则表示产生负效应。
本研究将总异质性QT分为各亚组的组间异质性QB和组内异质性QW, 通过组间异质性(QB)来检验同一指标的效应量在不同亚组(生态系统类型、植物种类、施氮形式、施氮水平和持续时间)之间的差异, 具体计算过程见公式(8)[81]。
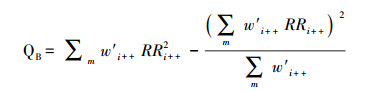
|
(8) |
式中, m代表某一分组的组数, i代表第i个亚组, RRi++代表第i个亚组的总效应量。
当某指标的95%置信区间和“0”没有交叉且显著性在0.05水平时, 则表明该效应量具有显著差异。上述计算过程通过METAWIN软件实现。
2 结果与分析 2.1 氮沉降对生物量分配的影响总体而言, 外源施氮能有效促进植物生物量的累积, 植物总生物量在施氮条件下明显增加(平均效应量=0.266;95%CI: 0.155—0.378; 图 1)。在11个选定指标中, 地上生物量、地下生物量、根冠比、总生物量、叶生物量、茎生物量6个指标对外源施氮响应明显。外源施氮会显著提高植物地上生物量的分配(图 1), 而对地下生物量的促进作用要低于地上部分。这就导致植物的根冠比在施氮条件下明显降低(平均效应量=-0.055;95%CI:-0.106—-0.004;图 1), 说明植物地上部分对氮沉降更敏感。对于植物的地上部分, 茎生物量和叶生物量在施氮条件下都显著增加, 相反的, 施氮对植物地下部分的细根和粗根生物量虽然均有促进作用, 但并不显著。根重比在施氮条件下有降低趋势(平均效应量=-0.186;95%CI:-0.426—0.054;图 1), 茎重比(平均效应量=0.060;95%CI:-0.036—0.155;图 1)和叶重比(平均效应量=0.024;95%CI:-0.070—0.119;图 1)表现为增加趋势, 但并不显著。

|
| 图 1 外源氮输入对植物生物量分配各指标的影响 Fig. 1 Impacts of elevated nitrogen on biomass allocation variables of plant AGB:地上生物量, aboveground biomass; BGB:地下生物量, belowground biomass; R/S:根冠比, Root/shoot; TB:总生物量, total biomass; LB:叶生物量, leaf biomass; SB:茎生物量, stem biomass; FRB:细根生物量, fine root biomass; CRB:粗根生物量, coarse root biomass; SWR:茎重比, stem weight ratio; RWR:根重比, root weight ratio; LWR:叶重比, leaf weight ratio; 效应量=(平均值±95%置信区); 图右括号内的数值代表观测值个数 |
本研究将生态系统类型分为森林、草原和其他三类, 不同生态系统植物的根冠比对模拟氮沉降的响应差异极显著(表 2)。施氮会显著降低森林生态系统中植物的根冠比(图 2c)。然而在草原中, 氮输入对植物根冠比不存在显著影响。植物总生物量在不同生态系统中对模拟氮沉降的响应差异极显著, 总的来看, 氮沉降在各生态系统中均有利于植物生物量的累积, 其中草原(平均效应量=0.354;95%CI:0.161—0.547;图 2d)以及其他(平均效应量=0.553;95%CI:0.303—0.802;图 2c)均表现为显著增加。与根冠比和总生物量不同, 不同生态系统类型和氮沉降对植物地上及地下生物量分配影响的关联性很弱, 即生态系统类型对植物地上和地下生物量的影响差异不显著(表 2)。
| 变量 Variables |
地上生物量(AGB) | 地下生物量(BGB) | 根冠比(R/S) | 总生物量(TB) | |||||||
| QB | P | QB | P | QB | P | QB | P | ||||
| 生态系统类型Ecosystem type | 2.499 | 0.287 | 5.846 | 0.054 | 13.730 | 0.001** | 12.460 | 0.002** | |||
| 植物类型Vegetation type | 0.932 | 0.919 | 0.039 | 0.980 | 19.730 | < 0.001** | 18.924 | < 0.001** | |||
| 氮形式N-form | 2.692 | 0.260 | 0.232 | 0.630 | 6.380 | 0.041* | 0.591 | 0.744 | |||
| 氮水平N-addition level | 8.179 | 0.017* | 3.073 | 0.215 | 3.050 | 0.218 | 6.915 | 0.032* | |||
| 持续时间Duration | 11.683 | 0.003** | 20.952 | < 0.001** | 0.870 | 0.646 | 1.173 | 0.556 | |||
| QB:组间异质性, between-group heterogeneity; *代表在P<0.05处差异显著, **代表差异极显著 | |||||||||||

|
| 图 2 不同生态系统类型、植物类型、氮形式、施氮水平和施氮持续时间对植物生物量分配的影响 Fig. 2 Effect of elevated nitrogen on biomass allocation with different ecological type, vegetation type, N form, N level and duration a:地上生物量, aboveground biomass; b:地下生物量, belowground biomass; c:根冠比, Root/shoot; d:总生物量, total biomass; 图右括号内的数值代表观测值个数 |
本文将植物分为草本、灌木、针叶、阔叶和其他五类, 不同植物的根冠比对模拟氮沉降的响应差异极显著(表 2)。阔叶树种的根冠比在氮输入条件下显著降低(平均效应量=-0.155;95%CI:-0.223—-0.086;图 2c), 但针叶树种、灌木和草本植物的根冠比在氮输入下并无显著变化。不同植物的总生物量对氮沉降的响应差异极显著(表 2), 其中草本植物(平均效应量=0.371;95%CI:0.186—0.556;图 2d)和阔叶树种(平均效应量=0.265;95%CI:0.098—0.433;图 2d)的总生物量在氮沉降影响下均明显增加, 氮沉降对针叶树种总生物量影响不显著。不同植物类型的地上生物量和地下生物量差异不显著(表 2)。
2.4 不同氮肥形式下植物生物量分配对模拟氮沉降的响应现有的大量氮沉降模拟研究以硝酸铵(NH4NO3)和尿素(CO(NH2)2)为主, 本研究中将氮肥形式分为硝酸铵、尿素和其他3种。氮肥形式对植物根冠比的影响差异显著, 硝酸铵作用下植物根冠比明显降低(平均效应量=-0.102;95%CI:-0.162—-0.041;图 2c), 尿素和其他氮肥对植物根冠比的影响不明显。氮肥种类对植物地上、地下和总生物量影响的差异不显著(表 2)。
2.5 不同施氮水平下植物生物量分配对模拟氮沉降的响应本研究中包含3个施氮水平, 高氮(HN, ≥120 kg hm-2 a-1)、中氮(MN, 60—120 kg hm-2 a-1)和低氮(LN, ≤60 kg hm-2 a-1)。不同施氮水平对植物地上生物量和总生物量的影响差异显著, 地上生物量和总生物量在不同施氮水平下均明显增加, 其中中氮水平(60—120 kg hm -2 a-1)增加最多, 分别为(平均效应量=0.334;95%CI:0.225—0.443;图 2a)和(平均效应量=0.475;95%CI:0.270—0.680;图 2d)。高氮水平和低氮水平植物地上生物量增加相接近, 分别为(平均效应量=0.176;95%CI:0.058—0.294;图 2a)和(平均效应量=0.155;95%CI:0.085—0.225;图 2d)。高氮水平的总生物量增加量低于低氮水平, 分别为(平均效应量=0.163;95%CI:0.022—0.304;图 2d)和(平均效应量=0.230;95%CI:0.055—0.405;图 2d)。不同施氮水平对植物地下生物量和总生物量的影响差异不显著(表 2)。
2.6 不同施氮持续时间下植物生物量分配对模拟氮沉降的响应本研究将施氮持续时间分为3个阶段, 不同持续时间对植物地上和地下生物量的影响差异极显著, 施氮短于1年及1—3年会明显提高植物地上生物量分配, 当施氮3年及以上时这种促进作用变得不明显(图 2a)。施氮短于1年时, 植物地下生物量分配明显提高(平均效应量=0.337;95%CI:0.176—0.498;图 2b), 继续延长施氮时间, 植物地下生物量分配变化不再明显(图 2b)。植物根冠比在不同施氮持续时间普遍降低, 总生物量普遍增加, 但施氮持续时间对根冠比和总生物量的影响差异并不显著(表 2)。
3 讨论与结论 3.1 讨论Fu等[82]的Meta分析研究表明, 氮沉降对植物地上生物量有明显的促进作用, 与本研究结果一致, 植物地上生物量在氮沉降下明显增加可能是因为氮沉降增加了土壤有效氮, 土壤养分充足使植物分配更多生物量给地上组织造成的[83-84]。本研究发现, 茎生物量在氮输入条件下表现为明显增加趋势, 这和Lu等[85]的Meta分析结果一致, 可能原因是氮沉降使植物养分运输能力加强, 进而表现为茎部的显著增加。除上述结论外, 本研究还发现随施氮时间延长(本研究中为≥3年), 氮处理对植物地上生物量的促进作用逐渐消失, 由此可见氮沉降对植物生长的促进作用存在时间差异。因此, 如果大气氮沉降在未来得不到有效控制, 其对植物生长所产生的抑制作用会随时间推移而慢慢显现出来。施氮水平的高低也会显著影响植物地上生物量, 陆生植物地上生物量在中氮水平所受促进作用最大, 更高的施氮水平反而会减弱促进作用, 大量研究也发现了同样的结论[28, 86]。造成这一现象的可能原因是长期高浓度施氮会使土壤酸化[77-88]或养分失衡[89-90]进而抑制植物生长。由于Metawin只能用于分析单因素因子对效应值的影响, 而植物生物量分配对氮沉降的响应受多因子控制, 如氮沉降量、沉降时间等, 因此在进行植物生物量分配对氮沉降响应的亚组分析时, 可能存在一定的局限性。
根冠比在研究陆地生态系统中碳、氮分配和储存时起着重要的指示作用, 预测植物在应对全球气候变化情景时是如何进行地上、地下生物量分配具有重要意义。大量有关氮沉降模拟试验的Meta分析结果表明外源施氮会显著降低植物的根冠比[85, 91-92], 与本研究的结果一致。植物地上生物量对氮沉降的敏感性要显著高于地下[93], 根冠比降低可能和地上生物量对氮沉降响应更敏感有关。本研究发现生态系统类型会显著影响植物的地上、地下生物量分配对氮沉降的响应, 与Wang等[94]的结果一致。相比于森林生态系统, 草地生态系统中植物根冠比在氮沉降下减小并不明显, 可能原因是在氮输入时植物的光合作用会显著加强, 植物对水分和养分的需求也会随之增强, 进而使更多的有机物和能源分配给根系用以维持土壤和根系之间的水分运输和养分获取, 而这种现象草地比森林更明显[95-96], 本研究中草地植物的地下生物量在外源氮输入时有增加趋势, 也进一步验证了上述解释。植物种类的差异会显著影响植物根冠比对模拟氮沉降的响应[64, 97], 本研究证实了上述结论, 除阔叶树外, 其他植物根冠比对氮沉降的响应与未分类前存在差异, 可能原因是样本量太小。与本研究不同的是Fu等[82]的Meta分析发现氮沉降对根冠比的影响不显著, 可能是因为Fu等的Meta分析只研究了高山植物根冠比在氮沉降下的变化。
植物地下生物量是全球陆地生态系统碳储量的重要组成部分, 在全球生态系统碳循环中起重要作用。本研究中植物细根和粗根生物量在模拟氮沉降条件下并没有显著变化(图 1), 但总的地下生物量却显著增加, 与Liu等[98]的Meta分析结果一致, 这表明当前植物生长仍处于氮限制状态。细根生物量在氮输入时并不会增加, 但细根周转率和呼吸却会显著提高, 植物主要靠细根吸收养分, 细根周转率提高会促进植物对养分的吸收[91], 此外, 根系呼吸和根系中的氮含量成正比[99], 因此, 即使细根生物量没有显著增加, 但植物在氮沉降下依然可以加强对土壤有效氮的吸收。氮沉降对植物地下生物量的促进作用只在短时间内表现, 当沉降持续时间过长, 促进作用消失, 可能是长期持续性供氮使土壤处于氮饱和状态, 进而限制了地下部分的生长。
目前有关氮输入对植物生物量累积作用较为一致的结论是:氮输入能有效促进氮缺乏生态系统植物生物量累积, 但在氮饱和生态系统中却有相反结论[100-101]。本研究发现尽管植物生物量累积对氮输入的响应较一致, 但生态系统类型、植物类型和施氮水平等不同均会显著影响植物生物量累积对模拟氮沉降的响应。目前所进行的氮沉降模拟研究在不同陆地生态系统均有开展, 草原约覆盖了陆地表面的25%, 通常处于氮限制状态[102-103], 本研究中氮沉降对草原生物量的促进作用要明显高于森林, Xia等[104]的Meta分析结果也表明氮输入对草本生物量的促进作用要显著大于木本。其原因可能是氮沉降会缓解草原的氮限制状态, 使草本植物得以旺盛生长; 而对于森林, 尽管温带森林处于氮限制状态, 但热带森林一般处于氮饱和状态[105](本研究中森林的试验样地以热带、亚热带居多), 这也就很好的诠释了为何草原中植物生物量受氮沉降的促进作用要明显高于森林, 这一变化对未来群落演替和植被动态具有重要意义。植物生物量累积在中氮水平(本研究为60—120 kg hm -2 a-1)达到最大, 这和地上生物量的变化一致, 也进一步说明植物生物量在模拟氮沉降研究中的累积主要是由地上生物量增加引起的。
3.2 结论(1) 氮沉降会对植物的地上生物量产生显著影响。具体表现为:地上生物量在外源氮输入时显著增加, 叶生物量和茎生物量在氮输入时均表现出明显增加趋势, 这可能和氮输入条件下植物加强光合作用和养分运输有关, 植物地上部分所受的促进作用表明植物冠层结构有可能在氮沉降下扩大。
(2) 植物地下生物量在外源氮输入时所受促进作用不及地上部分, 氮输入对细根和粗根生物量均表现出促进作用, 但并不显著。植物地上和地下生物量对氮输入响应的差异导致根冠比在氮输入时显著降低, 这说明相比于根部, 植物的地上部分对氮输入更加敏感, 即地上部分更有可能从短期氮沉降中获利。
(3) 生态系统类型和植物种类会显著影响植物根冠比和总生物量对氮输入的响应。氮沉降对草地(草本)的促进作用要优于森林(木本), 这说明氮沉降有可能会对未来的植被动态及群落演替产生影响。此外, 根冠比对氮沉降的响应会显著受到施氮形式影响, 相比于尿素, 硝酸铵对根冠比的作用更明显, 这说明在氮沉降对植物生物量影响的模拟试验中, 施氮形式有可能是造成各独立研究结果差异的一个重要原因。
(4) 短期内氮沉降将有利于植物生物量累积, 随着时间延长促进作用逐渐消失。这暗示着未来氮沉降背景下植物生长可能受限, 这为提前应对氮沉降对植物生长可能产生的消极影响起到了很好的警示作用, 也可为未来植被和生态系统管理提供理论指导和科学依据。
| [1] | Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle:recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889–892. DOI:10.1126/science.1136674 |
| [2] | Liu X J, Duan L, Mo J M, Du E Z, Shen J L, Lu X K, Zhang Y, Zhou X B, He C, Zhang F S. Nitrogen deposition and its ecological impact in China:an overview. Environmental pollution, 2011, 159(10): 2251–2264. DOI:10.1016/j.envpol.2010.08.002 |
| [3] | De Schrijver A, De Frenne P, Ampoorter E, Van Nevel L, Demey A, Wuyts K, Verheyen K. Cumulative nitrogen input drives species loss in terrestrial ecosystems. Global Ecology and Biogeography, 2011, 20(6): 803–816. DOI:10.1111/j.1466-8238.2011.00652.x |
| [4] | Yang Y, Guo J Y, Wang G X, Yang L D, Yang Y. Effects of drought and nitrogen addition on photosynthetic characteristics and resource allocation of Abies fabri seedlings in eastern Tibetan Plateau. New Forests, 2012, 43(4): 505–518. DOI:10.1007/s11056-011-9295-3 |
| [5] | 李明月, 王健, 王振兴, 吴晓燕, 黄儒珠, 朱锦懋. 模拟氮沉降条件下木荷幼苗光合特性、生物量与C、N、P分配格局. 生态学报, 2013, 33(5): 1569–1577. |
| [6] | Yang Y H, Fang J Y, Ma W H, Guo D L, Mohammat A. Large-scale pattern of biomass partitioning across China's grasslands. Global Ecology and Biogeography, 2010, 19(2): 268–277. DOI:10.1111/j.1466-8238.2009.00502.x |
| [7] | Callaway J C, Sullivan G, Zedler J B. Species-rich plantings increase biomass and nitrogen accumulation in a wetland restoration experiment. Ecological Applications, 2003, 13(6): 1626–1639. DOI:10.1890/02-5144 |
| [8] | 祁瑜, 黄永梅, 王艳, 赵杰, 张景慧. 施氮对几种草地植物生物量及其分配的影响. 生态学报, 2011, 31(18): 5121–5129. |
| [9] | Kiær L P, Weisbach A N, Weiner J. Root and shoot competition:a meta-analysis. Journal of Ecology, 2013, 101(5): 1298–1312. DOI:10.1111/jec.2013.101.issue-5 |
| [10] | 涂丽华, 胡庭兴, 张健, 李仁洪, 何远洋, 田祥宇, 肖银龙, 景建飞. 模拟氮沉降对华西雨屏区慈竹林土壤活性有机碳库和根生物量的影响. 生态学报, 2010, 30(9): 2286–2294. |
| [11] | Dong J L, Li X, Duan Z Q. Biomass allocation and organs growth of cucumber (Cucumis sativus L.) under elevated CO2 and different N supply. Archives of Agronomy and Soil Science, 2016, 62(2): 277–288. DOI:10.1080/03650340.2015.1045497 |
| [12] | Ackerly D D, Dudley S A, Sultan S E, Schmitt J, Coleman J S, Linder C R, Sandquist D R, Geber M A, Evans A S, Dawson T E, Lechowicz M J. The evolution of plant ecophysiological traits:recent advances and future directions:new research addresses natural selection, genetic constraints, and the adaptive evolution of plant ecophysiological traits. BioScience, 2000, 50(11): 979–995. DOI:10.1641/0006-3568(2000)050[0979:TEOPET]2.0.CO;2 |
| [13] | Fortunel C, Fine P V A, Baraloto C. Leaf, stem and root tissue strategies across 758 Neotropical tree species. Functional Ecology, 2012, 26(5): 1153–1161. DOI:10.1111/j.1365-2435.2012.02020.x |
| [14] | Roa-Fuentes L L, Campo J, Parra-Tabla V. Plant biomass allocation across a precipitation gradient:an approach to seasonally dry tropical forest at Yucatán, Mexico. Ecosystems, 2012, 15(8): 1234–1244. DOI:10.1007/s10021-012-9578-3 |
| [15] | White K P, Langley J A, Cahoon D R, Megonigal J P. C3 and C4 biomass allocation responses to elevated CO2 and nitrogen:contrasting resource capture strategies. Estuaries and Coasts, 2012, 35(4): 1028–1035. DOI:10.1007/s12237-012-9500-4 |
| [16] | Poorter H, Niklas K J, Reich P B, Oleksyn J, Poot P, Mommer L. Biomass allocation to leaves, stems and roots:meta-analyses of interspecific variation and environmental control. New Phytologist, 2012, 193(1): 30–50. DOI:10.1111/j.1469-8137.2011.03952.x |
| [17] | Sun J, Cheng G W, Li W P. Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau. Biogeosciences, 2013, 10(3): 1707–1715. DOI:10.5194/bg-10-1707-2013 |
| [18] | Xie J B, Tang L S, Wang Z Y, Xu G Q, Li Y. Distinguishing the biomass allocation variance resulting from ontogenetic drift or acclimation to soil texture. PLoS One, 2012, 7(7): e41502. DOI:10.1371/journal.pone.0041502 |
| [19] | Zong N, Shi P L, Chai X, Jiang J, Zhang X Z, Song M H. Responses of ecosystem respiration to nitrogen enrichment and clipping mediated by soil acidification in an alpine meadow. Pedobiologia, 2017, 60: 1–10. DOI:10.1016/j.pedobi.2016.11.001 |
| [20] | Wang D, He H L, Gao Q, Zhao C Z, Zhao W Q, Yin C Y, Chen X L, Ma Z L, Li D D, Sun D D, Cheng X Y, Liu Q. Effects of short-term N addition on plant biomass allocation and C and N pools of the Sibiraea angustata scrub ecosystem. European Journal of Soil Science, 2017, 68(2): 212–220. DOI:10.1111/ejss.2017.68.issue-2 |
| [21] | Liu Y, Li P, Wang G L, Liu G B, Li Z B. Above-and below-ground biomass distribution and morphological characteristics respond to nitrogen addition in Pinus tabuliformis. New Zealand Journal of Forestry Science, 2016, 46(1): 25. DOI:10.1186/s40490-016-0083-x |
| [22] | Li J Y, Guo Q X, Zhang J X, Korpelainen H, Li C Y. Effects of nitrogen and phosphorus supply on growth and physiological traits of two Larix species. Environmental and Experimental Botany, 2016, 130: 206–215. DOI:10.1016/j.envexpbot.2016.06.006 |
| [23] | Huangfu C H, Li H Y, Chen X W, Liu H M, Wang H, Yang D L. Response of an invasive plant, Flaveria bidentis, to nitrogen addition:a test of form-preference uptake. Biological Invasions, 2016, 18(11): 3365–3380. DOI:10.1007/s10530-016-1231-1 |
| [24] | Chen Guo H X, Xu B, Wu Y, Shi F, Wu C, Wu N. Allometric partitioning theory versus optimal partitioning theory:the adjustment of biomass allocation and internal CN balance to shading and nitrogen addition in Fritillaria unibracteata (Liliaceae). Polish Journal of Ecology, 2016, 64(2): 189–199. DOI:10.3161/15052249PJE2016.64.2.004 |
| [25] | Meyer-Grünefeld M, Calvo L, Marcos E, Oheimb G V, Haerdele W. Impacts of drought and nitrogen addition on Calluna heathlands differ with plant life-history stage. Journal of Ecology, 2015, 103(5): 1141–1152. DOI:10.1111/1365-2745.12446 |
| [26] | Mei X, Zhang X. Effects of N and P additions to water column on growth of Vallisneria natans. Journal of Aquatic Plant Management, 2015, 53: 36–43. |
| [27] | Li K H, Liu X J, Song L, Gong Y M, Lu C F, Yue P, Tian C Y, Zhang F S. Response of alpine grassland to elevated nitrogen deposition and water supply in China. Oecologia, 2015, 177(1): 65–72. DOI:10.1007/s00442-014-3122-4 |
| [28] | Wang G L, Liu F. Carbon allocation of Chinese pine seedlings along a nitrogen addition gradient. Forest Ecology and Management, 2014, 334: 114–121. DOI:10.1016/j.foreco.2014.09.004 |
| [29] | Palmroth S, Bach L H, Nordin A, Palmqvist K. Nitrogen-addition effects on leaf traits and photosynthetic carbon gain of boreal forest understory shrubs. Oecologia, 2014, 175(2): 457–470. DOI:10.1007/s00442-014-2923-9 |
| [30] | Wang G L, Fahey T J, Xue S, Liu F. Root morphology and architecture respond to N addition in Pinus tabuliformis, west China. Oecologia, 2013, 171(2): 583–590. DOI:10.1007/s00442-012-2441-6 |
| [31] | Siegenthaler A, Buttler A, Grosvernier P, Gobat J M, Nilsson M B, Mitchell E A D. Factors modulating cottongrass seedling growth stimulation to enhanced nitrogen and carbon dioxide:compensatory tradeoffs in leaf dynamics and allocation to meet potassium-limited growth. Oecologia, 2013, 171(2): 557–570. DOI:10.1007/s00442-012-2415-8 |
| [32] | Tian D, Pan Q, Simmons M, Chao L H, Du B, Bai Y, Wang H, Han X. Hierarchical reproductive allocation and allometry within a perennial bunchgrass after 11 years of nutrient addition. PLoS One, 2012, 7(9): e42833. DOI:10.1371/journal.pone.0042833 |
| [33] | Santiago L S, Wright S J, Harms K E, Yaviit J B, Korine C, Garcia M N, Turner B L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. Journal of Ecology, 2012, 100(2): 309–316. DOI:10.1111/jec.2012.100.issue-2 |
| [34] | LESKOVŠEK R, Datta A, Knezevic S Z, Simončič A. Common ragweed (Ambrosia artemisiifolia) dry matter allocation and partitioning under different nitrogen and density levels. Weed Biology and Management, 2012, 12(2): 98–108. DOI:10.1111/wbm.2012.12.issue-2 |
| [35] | Zhou X B, Zhang Y M, Ji X H, Downing A, Serpe M. Combined effects of nitrogen deposition and water stress on growth and physiological responses of two annual desert plants in northwestern China. Environmental and Experimental Botany, 2011, 74: 1–8. DOI:10.1016/j.envexpbot.2010.12.005 |
| [36] | Singh V. Effect of nitrogen addition on the architecture and biomass allocation of two invasive plant species (Ageratum conyzoides L. and Parthenium hysterophorus L.). Proceedings of The National Academy of Sciences India Section B-biological Sciences, 2010, 81(3): 348–358. |
| [37] | Pan Q M, Bai Y F, Wu J G, Han X G. Hierarchical plant responses and diversity loss after nitrogen addition:testing three functionally-based hypotheses in the Inner Mongolia grassland. PLoS One, 2011, 6(5): e20078. DOI:10.1371/journal.pone.0020078 |
| [38] | Zhang L, Yang Y X, Zhan X Y, Zhang C J, Zhou S X, Wu D X. Responses of a dominant temperate grassland plant (Leymus chinensis) to elevated CO2 and N addition in China, 2010, 39: 251-259. |
| [39] | Iversen C M, Bridgham S D, Kellogg L E. Scaling plant nitrogen use and uptake efficiencies in response to nutrient addition in peatlands. Ecology, 2010, 91(3): 693–707. DOI:10.1890/09-0064.1 |
| [40] | Zhao C Z, Liu Q. Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization. Canadian Journal of Forest Research, 2008, 39(1): 1–11. |
| [41] | Laclau J P, Almeida J C R, Gonçalves J L M, Saint-André, Ventura M, Ranger J, Moreira R M, Nouvellon Y. Influence of nitrogen and potassinm fertilization on leaf lifespan and allocation of above-ground growth in Eucalyptus plantations. Tree Physiology, 2009, 29(1): 111–124. |
| [42] | Zhao W, Chen S P, Lin G H. Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecology, 2008, 196(1): 85–99. DOI:10.1007/s11258-007-9336-3 |
| [43] | Mo J M, Li D J, Gundersen P. Seedling growth response of two tropical tree species to nitrogen deposition in southern China. European Journal of Forest Research, 2008, 127(4): 275–283. DOI:10.1007/s10342-008-0203-0 |
| [44] | Cai Z Q, Poorter L, Han Q, Bongers F. Effects of light and nutrients on seedlings of tropical Bauhinia lianas and trees. Tree physiology, 2008, 28(8): 1277–1285. DOI:10.1093/treephys/28.8.1277 |
| [45] | Manninen O H, Stark S, Kytöviita M M, Tolvanen A. Individual and combined effects of disturbance and N addition on understorey vegetation in a subarctic mountain birch forest. Journal of Vegetation Science, 2011, 22(2): 262–272. DOI:10.1111/jvs.2011.22.issue-2 |
| [46] | Lamb E G, Shore B H, Cahill J F. Water and nitrogen addition differentially impact plant competition in a native rough fescue grassland. Plant Ecology, 2007, 192(1): 21–33. DOI:10.1007/s11258-006-9222-4 |
| [47] | Wu M Y, Niu S L, Wan S Q. Contrasting effects of clipping and nutrient addition on reproductive traits of Heteropappus altaicus at the individual and population levels. Ecological research, 2010, 25(4): 867–874. DOI:10.1007/s11284-010-0718-y |
| [48] | Holub P, T ma I. The effect of enhanced nitrogen on aboveground biomass allocation and nutrient resorption in the fern Athyrium distentifolium. Plant ecology, 2010, 207(2): 373–380. DOI:10.1007/s11258-009-9681-5 |
| [49] | Falk K, Friedrich U, von Oheimb G V, Mischke K, Merkle K, Meyer H, Härdtle W. Molinia caerulea responses to N and P fertilisation in a dry heathland ecosystem (NW-Germany). Plant ecology, 2010, 209(1): 47–56. DOI:10.1007/s11258-010-9720-2 |
| [50] | Macek P, Rejmánková E. Response of emergent macrophytes to experimental nutrient and salinity additions. Functional Ecology, 2007, 21(3): 478–488. DOI:10.1111/fec.2007.21.issue-3 |
| [51] | Tanis S R, McCullough D G, Cregg B M. Effects of paclobutrazol and fertilizer on the physiology, growth and biomass allocation of three Fraxinus species. Urban Forestry & Urban Greening, 2015, 14(3): 590–598. |
| [52] | Zhou X B, Zhang Y M, Niklas K J. Sensitivity of growth and biomass allocation patterns to increasing nitrogen:a comparison between ephemerals and annuals in the Gurbantunggut Desert, north-western China. Annals of botany, 2014, 113(3): 501–511. DOI:10.1093/aob/mct275 |
| [53] | Yan J, Xu C Y, Wei H X. Response of carbon and nitrogen allocation in two hybrid poplar clones to soil nitrogen in Northern China. J Food Agric Environ, 2013, 11(2): 1050–1054. |
| [54] | Bonomelli C, Artacho P. Nitrogen application to non-bearing 'Bing' sweet cherry trees on Gisela R6 rootstock:Effects on accumulation and partitioning of biomass and nitrogen. Scientia Horticulturae, 2013, 162: 293–304. DOI:10.1016/j.scienta.2013.08.024 |
| [55] | Blanke V, Bassin S, Volk M, Fuhrer J. Nitrogen deposition effects on subalpine grassland:the role of nutrient limitations and changes in mycorrhizal abundance. Acta Oecologica, 2012, 45: 57–65. DOI:10.1016/j.actao.2012.09.002 |
| [56] | Samuelson L J, Farris M G, Stokes T A, Coleman M D. Fertilization but not irrigation influences hydraulic traits in plantation-grown loblolly pine. Forest ecology and management, 2008, 255(8/9): 3331–3339. |
| [57] | Zhao G Y, Liu J S, Wang Y, Dou J X, Dong X Y. Effects of elevated CO2 concentration and nitrogen supply on biomass and active carbon of freshwater marsh after two growing seasons in Sanjiang Plain, Northeast China. Journal of Environmental Sciences, 2009, 21(10): 1393–1399. DOI:10.1016/S1001-0742(08)62431-6 |
| [58] | Cusack D F, Silver W L, Torn M S, McDowell W H. Effects of nitrogen additions on above-and belowground carbon dynamics in two tropical forests. Biogeochemistry, 2011, 104(1/3): 203–225. |
| [59] | Deng Q, Zhou G, Liu J, Liu S, Duan H, Zhang D. Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences, 2010, 7(1): 315–328. DOI:10.5194/bg-7-315-2010 |
| [60] | Ochoa-Hueso R, Pérez-Corona M E, Manrique E. Impacts of simulated N deposition on plants and mycorrhizae from Spanish semiarid Mediterranean shrublands. Ecosystems, 2013, 16(5): 838–851. DOI:10.1007/s10021-013-9655-2 |
| [61] | Liu Q, Yin H J, Chen J S, Zhao C Z, Cheng X Y, Wei Y Y, Lin B. Belowground responses of Picea asperata seedlings to warming and nitrogen fertilization in the eastern Tibetan Plateau. Ecological Research, 2011, 26(3): 637–648. DOI:10.1007/s11284-011-0824-5 |
| [62] | 王晓荣, 潘磊, 庞宏东, 郑京津, 王瑞文. 模拟氮沉降对亚热带栎属树种幼苗生长、生物量累积及光合特性的影响. 中南林业科技大学学报, 2016, 36(1): 78–85. |
| [63] | 刘双娥, 李义勇, 方熊, 黄文娟, 龙凤玲, 刘菊秀. 不同氮添加量和添加方式对南亚热带4个主要树种幼苗生长的影响. 植物生态学报, 2015, 39(10): 950–961. DOI:10.17521/cjpe.2015.0092 |
| [64] | 赵亮, 周国逸, 张德强, 段洪浪, 刘菊秀. CO2浓度升高和氮沉降对南亚热带主要乡土树种及群落生物量的影响. 应用生态学报, 2011, 22(8): 1947–1954. |
| [65] | 赵光影, 刘景双, 王洋. CO2浓度升高与氮添加对三江平原湿地小叶章生长的影响. 应用生态学报, 2011, 22(6): 1653–1658. |
| [66] | 周晓兵, 张元明, 王莎莎, 张丙昌. 模拟氮沉降和干旱对准噶尔盆地两种一年生荒漠植物生长和光合生理的影响. 植物生态学报, 2010, 34(12): 1394–1403. |
| [67] | 辛月, 尚博, 陈兴玲, 冯兆忠. 氮沉降对臭氧胁迫下青杨光合特性和生物量的影响. 环境科学, 2016, 37(9): 3642–3649. |
| [68] | Adamek M, Corre M D, Hölscher D. Responses of fine roots to experimental nitrogen addition in a tropical lower montane rain forest, Panama. Journal of Tropical Ecology, 2011, 27(1): 73–81. DOI:10.1017/S0266467410000507 |
| [69] | Bradford M A, Fierer N, Jackson R B, Maddox T R, Reynolds J F. Nonlinear root-derived carbon sequestration across a gradient of nitrogen and phosphorous deposition in experimental mesocosms. Global Change Biology, 2008, 14(5): 1113–1124. DOI:10.1111/j.1365-2486.2008.01564.x |
| [70] | Burton A J, Jarvey J C, Jarvi M P, Zak D R, Pregitzer K S. Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests. Global Change Biology, 2012, 18(1): 258–266. DOI:10.1111/j.1365-2486.2011.02527.x |
| [71] | Cheng X L, Luo Y Q, Su B, Verburg P S J, Hui D F, Obrist D, Arnone J A, Johnson D W, David Evans R. Responses of net ecosystem CO2 exchange to nitrogen fertilization in experimentally manipulated grassland ecosystems. Agricultural and Forest Meteorology, 2009, 149(11): 1956–1963. DOI:10.1016/j.agrformet.2009.07.001 |
| [72] | Hasselquist N J, Metcalfe D B, Högberg P. Contrasting effects of low and high nitrogen additions on soil CO2 flux components and ectomycorrhizal fungal sporocarp production in a boreal forest. Global Change Biology, 2012, 18(12): 3596–3605. DOI:10.1111/gcb.12001 |
| [73] | Hungate B A, Hart S C, Selmants P C, Boyle S I, Gehring C A. Soil responses to management, increased precipitation, and added nitrogen in ponderosa pine forests. Ecological Applications, 2007, 17(5): 1352–1365. DOI:10.1890/06-1187.1 |
| [74] | Jiang X Y, Cao L X, Zhang R D. Changes of labile and recalcitrant carbon pools under nitrogen addition in a city lawn soil. Journal of Soils and Sediments, 2014, 14(3): 515–524. DOI:10.1007/s11368-013-0822-z |
| [75] | Noguchi K, Nagakura J, Konôpka B, Sakata T, Kaneko S, Takahashi M. Fine-root dynamics in sugi (Cryptomeria japonica) under manipulated soil nitrogen conditions. Plant and Soil, 2013, 364(1/2): 159–169. |
| [76] | Tu L H, Hu T X, Zhang J, Li R H, Dai H Z, Luo S H. Short-term simulated nitrogen deposition increases carbon sequestration in a Pleioblastus amarus plantation. Plant and Soil, 2011, 340(1/2): 383–396. |
| [77] | Tu L H, Hu T X, Zhang J, Li X W, Hu H L, Liu L, Xiao Y L. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biology and Biochemistry, 2013, 58: 255–264. DOI:10.1016/j.soilbio.2012.12.005 |
| [78] | Wang C G, Han S J, Zhou Y M, Yan C F, Cheng X B, Zheng X B, Li M H. Responses of fine roots and soil N availability to short-term nitrogen fertilization in a broad-leaved Korean pine mixed forest in northeastern China. PLoS One, 2012, 7(3): e31042. DOI:10.1371/journal.pone.0031042 |
| [79] | Zhu F F, Yoh M, Gilliam F S, Lu X K, Mo J M. Nutrient limitation in three lowland tropical forests in southern China receiving high nitrogen deposition:insights from fine root responses to nutrient additions. PLoS One, 2013, 8(12): e82661. DOI:10.1371/journal.pone.0082661 |
| [80] | Gao Q, Hasselquist N J, Palmroth S, Zheng Z M, You W H. Short-term response of soil respiration to nitrogen fertilization in a subtropical evergreen forest. Soil Biology and Biochemistry, 2014, 76: 297–300. DOI:10.1016/j.soilbio.2014.04.020 |
| [81] | 彭少麟, 唐小焱. Meta分析及其在生态学上的应用. 生态学杂志, 1998, 17(5): 74–79. |
| [82] | Fu G, Shen Z X. Response of alpine plants to nitrogen addition on the Tibetan Plateau:a meta-analysis. Journal of Plant Growth Regulation, 2016, 35(4): 974–979. DOI:10.1007/s00344-016-9595-0 |
| [83] | McConnaughay K D M, Coleman J S. Biomass allocation in plants:ontogeny or optimality? A test along three resource gradients. Ecology, 1999, 80(8): 2581–2593. DOI:10.1890/0012-9658(1999)080[2581:BAIPOO]2.0.CO;2 |
| [84] | Reef R, Slot M, Motro U, Motro M, Motro Y, Adame M F, Garcia M, Aranda J, Lovelock C E, Winter K. The effects of CO2 and nutrient fertilisation on the growth and temperature response of the mangrove Avicennia germinans. Photosynthesis Research, 2016, 129(2): 159–170. DOI:10.1007/s11120-016-0278-2 |
| [85] | Lu M, Zhou X H, Luo Y Q, Yang Y H, Fang C M, Chen J K, Li B. Minor stimulation of soil carbon storage by nitrogen addition:a meta-analysis. Agriculture, Ecosystems & Environment, 2011, 140(1/2): 234–244. |
| [86] | Sillen W M A, Dieleman W I J. Effects of elevated CO2 and N fertilization on plant and soil carbon pools of managed grasslands:a meta-analysis. Biogeosciences, 2012, 9(6): 2247–2258. DOI:10.5194/bg-9-2247-2012 |
| [87] | Van Breemen N, Burrough P A, Velthorst E J, Van Dobben H F, De Wit T, Ridder T B, Reijnders H F R. Soil acidification from atmospheric ammonium sulphate in forest canopy throughfall. Nature, 1982, 299(5883): 548–550. DOI:10.1038/299548a0 |
| [88] | Schulze E D. Air pollution and forest decline in a spruce (Picea abies) forest. Science, 1989, 244(4906): 776–783. DOI:10.1126/science.244.4906.776 |
| [89] | Linder S. Foliar analysis for detecting and correcting nutrient imbalances in Norway spruce. Ecological Bulletins, 1995(44): 178–190. |
| [90] | Gress S E, Nichols T D, Northcraft C C, Peterjohn W T. Nutrient limitation in soils exhibiting differing nitrogen availabilities:what lies beyond nitrogen saturation?. Ecology, 2007, 88(1): 119–130. DOI:10.1890/0012-9658(2007)88[119:NLISED]2.0.CO;2 |
| [91] | Li W B, Jin C J, Guan D X, Wang Q K, Wang A Z, Yuan F H, Wu J B. The effects of simulated nitrogen deposition on plant root traits:a meta-analysis. Soil Biology and Biochemistry, 2015, 82: 112–118. DOI:10.1016/j.soilbio.2015.01.001 |
| [92] | Li F R, Dudley T L, Chen B M, Chang X Y, Liang L Y, Peng S L. Responses of tree and insect herbivores to elevated nitrogen inputs:a meta-analysis. Acta Oecologica, 2016, 77: 160–167. DOI:10.1016/j.actao.2016.10.008 |
| [93] | Vogt K A, Grier C C, Vogt D J. Production, turnover, and nutrient dynamics of above-and belowground detritus of world forests. Advances in Ecological Research, 1986, 15: 303–377. DOI:10.1016/S0065-2504(08)60122-1 |
| [94] | Wang L M, Li L H, Chen X, Tian X, Wang X K, Luo G P. Biomass allocation patterns across China's terrestrial biomes. PLoS One, 2014, 9(4): e93566. DOI:10.1371/journal.pone.0093566 |
| [95] | Wang W, Guo J X, Oikawa T. Contribution of root to soil respiration and carbon balance in disturbed and undisturbed grassland communities, Northeast China. Journal of Biosciences, 2007, 32(2): 375–384. DOI:10.1007/s12038-007-0036-x |
| [96] | Schindlbacher A, Zechmeister-Boltenstern S, Jandl R. Carbon losses due to soil warming:do autotrophic and heterotrophic soil respiration respond equally?. Global Change Biology, 2009, 15(4): 901–913. DOI:10.1111/gcb.2009.15.issue-4 |
| [97] | Liu J X, Zhou G Y, Xu Z H, Duan H L, Li Y L, Zhang D Q. Photosynthesis acclimation, leaf nitrogen concentration, and growth of four tree species over 3 years in response to elevated carbon dioxide and nitrogen treatment in subtropical China. Journal of Soils and Sediments, 2011, 11(7): 1155–1164. DOI:10.1007/s11368-011-0398-4 |
| [98] | Liu L L, Greaver T L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecology Letters, 2010, 13(7): 819–828. DOI:10.1111/ele.2010.13.issue-7 |
| [99] | Zhang C P, Niu D C, Hall S J, Wen H Y, Li X D, Fu H, Wan C G, Elser J J. Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biology and Biochemistry, 2014, 75: 113–123. DOI:10.1016/j.soilbio.2014.04.013 |
| [100] | Nordin A, Strengbom J, Witzell J, Näsholm T, Ericson L. Nitrogen deposition and the biodiversity of boreal forests:implications for the nitrogen critical load. AMBIO:A Journal of the Human Environment, 2005, 34(1): 20–24. DOI:10.1579/0044-7447-34.1.20 |
| [101] | Galloway J N, Aber J D, Erisman J W, Seitzinger S P, Howarth R W, Cowling E B, Cosby B J. The nitrogen cascade. BioScience, 2003, 53(4): 341–356. DOI:10.1641/0006-3568(2003)053[0341:TNC]2.0.CO;2 |
| [102] | Elser J J, Bracken M E S, Cleland E E, Gruner D S, Harpole W S, Hillebrand H, Ngai J T, Seabloom E W, Shurin J B, Smith J E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 2007, 10(12): 1135–1142. DOI:10.1111/ele.2007.10.issue-12 |
| [103] | Brookshire E N J, Gerber S, Menge D N L, Hedin L O. Large losses of inorganic nitrogen from tropical rainforests suggest a lack of nitrogen limitation. Ecology Letters, 2012, 15(1): 9–16. DOI:10.1111/ele.2011.15.issue-1 |
| [104] | Xia J Y, Wan S Q. Global response patterns of terrestrial plant species to nitrogen addition. New Phytologist, 2008, 179(2): 428–439. DOI:10.1111/nph.2008.179.issue-2 |
| [105] | Matson P A, McDowell W H, Townsend A R, Vitousek P M. The globalization of N deposition: ecosystem consequences in tropical environments//Townsend A R, ed. New Perspectives on Nitrogen Cycling in the Temperate and Tropical Americas. Dordrecht: Springer, 1999: 67-83. |