 2018, Vol. 38
2018, Vol. 38文章信息
- 胡刚, 黎洁, 覃盈盈, 胡宝清, 刘熊, 张忠华.
- HU Gang, LI Jie, QIN Yingying, HU Baoqing, LIU Xiong, ZHANG Zhonghua.
- 广西北仑河口红树植物种群结构与动态特征
- Population structure and dynamics of mangrove species in Beilun Estuary, Guangxi, southern China
- 生态学报. 2018, 38(9): 3022-3034
- Acta Ecologica Sinica. 2018, 38(9): 3022-3034
- http://dx.doi.org/10.5846/stxb201705170918
-
文章历史
- 收稿日期: 2017-05-17
- 修订日期: 2017-11-29





2. 广西师范学院广西西江流域生态环境与一体化发展协同创新中心, 南宁 530001;
3. 广西师范学院地理科学与规划学院, 南宁 530001;
4. 广西师范大学生命科学学院, 桂林 541004;
5. 防城港市海洋环境监测预报中心, 防城港 538001;
6. 广西北仑河口国家级自然保护区管理处, 防城港 538021
2. The Collaborative Innovation Center of the Ecological Environment and Integration Development in the Xijiang River Basin, Guangxi Teachers Education University, Nanning 530001, China;
3. School of Geography and Planning, Guangxi Teachers Education University, Nanning 530001, China;
4. College of Life Sciences, Guangxi Normal University, Guilin 541004, China;
5. Marine Environmental Monitoring and Forecasting Center of Fangchenggang City, Fangchenggang 538001, China;
6. National Nature Reserve of Beilun Estuary, Fangchenggang 538021, China
种群结构与动态一直是生态学研究的核心内容[1-3]。种群是群落的基本组成部分, 种群结构的变化对群落特征产生直接影响, 能客观地表征群落的发展和演变趋势4]。植物种群的生物学和生态学特性是植物对环境条件长期适应和选择的结果, 种群大小或数量在时空尺度上的动态变化反映了种群的配置和发展趋势, 体现了种群和环境间的交互性关系及其在群落中的地位和作用[5-7]。因此, 解析植物种群的结构与动态对阐明群落的演替路线以及生态系统的结构和功能具有重要的意义。
红树林是生长于热带和亚热带海岸潮间带、受海水周期性浸淹的木本植物群落, 兼具有陆地生态和海洋生态特性, 是海岸重要生态关键区[8]。作为海岸潮间带特殊的植被生态系统, 红树林在防风消浪、固岸护堤、维护生物多样性和沿海生态安全等方面发挥着重要作用。我国红树林主要分布于广东、广西、海南、福建和台湾等省区, 其中广西的红树林主要位于北部湾海岸带的珍珠湾、钦州湾和英罗湾等地, 以白骨壤(Avicennia marina)、桐花树(Aegiceras corniculatum)、秋茄(Kandelia obovata)、红海榄(Rhizophora stylosa)和木榄(Bruguiera gymnorrhiza)等植物为主[9-11]。国内外学者和政府部门一直高度关注广西红树林的保护、恢复与合理利用[11-12]。然而, 伴随海岸带人为干扰和全球变化因素的加剧, 广西红树林生态系统遭受诸多负面因素的制约, 如生境丧失和破碎化、过度利用、环境污染和外来物种入侵等, 部分地区红树林呈现逐步衰退甚至消失殆尽的局面[13-15]。广西红树林面积由1960/1976年的9062.5hm2降至2010年的7054.3hm2, 近50年红树林面积减少了22.16%, 平均每年减少0.53%[14]。近些年, 通过人工红树林造林措施的积极实施, 广西部分滩涂的红树林面积呈现一定程度的稳中有增态势[16]。然而, 现存的天然红树林面积仍然面临逐年减少的威胁, 其科学研究和保育工作还亟待加强。
我国学者对广西红树林开展过较多生态学研究工作。例如, 林鹏等[9, 17-18]较早关注广西红树林, 在红树林的植物种类组成、群落类型、生物量与生产力特征和演替规律等方面开展过奠基性基础研究工作, 为科学认识广西红树林做出了重要贡献。范航清等[11, 19-22]在红树林分布与结构、生态恢复与重建、病虫害的防控等方面持续进行过大量富有成效的研究工作。此外, 梁士楚[23-25]、何斌源等[26]、李春干等[14]分别在红树植物种群和群落结构、生理生态以及遥感面积监测等方面开展过积极探索工作。上述学者主要侧重红树林恢复与重建措施以及大尺度时空分布演变等方面研究, 而对种群和群落层面的基础研究多见于十年前的早期文献报道。近些年, 广西红树林面临气候变化和人为干扰等多重复杂的环境胁迫, 亟需在变化环境下持续开展红树林结构和动态的长期生态学研究, 如红树植物的种群结构和分布格局的动态监测, 将有助于及时掌握红树林植物种群和群落结构现状, 预测其发展趋势, 为科学保护和管理红树林提供理论和实践支持。
广西北仑河口国家级保护区位于我国大陆海岸线最西南端的沿海地带, 该保护区分布有我国唯一的连片面积最大的边境红树林生态系统, 为研究天然红树林生态系统结构与功能提供珍贵样本。近年来, 对北仑河口红树林开展的生态学方面研究相对较少, 仅在红树植物群落组成[25]以及红树林生态恢复和重建[19, 22]等方面开展过一定研究工作, 而针对红树植物的种群结构和动态方面, 至今鲜有报道。植物种群的年龄结构、生命表、存活曲线、格局强度和时间序列预测等分析手段是进行种群结构与动态分析的重要方法, 借助这些方法可以分析种群的生存状态, 揭示种群和群落的发展趋势[3, 7, 27]。本研究采用上述分析方法对北仑河口保护区秋茄、桐花树、白骨壤和木榄4种优势红树植物的种群结构和动态进行分析和预测, 旨在阐明北仑河口红树植物种群与群落的结构特征和动态变化规律以及潜在的驱动机制, 为广西红树林生态系统的保护与恢复提供基础资料和科学依据。
1 研究区概况北仑河口国家级自然保护区(21°31′00″—21°37′30″N, 108°00′30″—108°16′30″E)地处广西防城港市和东兴市海域, 东南临北部湾, 西南与越南毗邻, 由东到西跨越珍珠湾、江平三岛和北仑河口, 岸线长105 km, 滩涂面积53 km2, 现有红树林面积约1274 hm2, 其中1081 hm2红树林分布于珍珠湾内[22]。该地区的气候属于南亚热带海洋季风气候, 年平均气温22.3℃, 极端最高温37.8℃, 极端最低温2.8℃, 年平均降水量2220.5 mm, 年均蒸发量为1400 mm;海域的潮汐类型为正规全日潮, 多年平均潮差2.22 m(以黄海基准面起算), 最大潮差5.64 m, 多年平均潮位0.34 m, 海水年平均温度23.5℃, 盐度23.1‰[22]。保护区现有红树植物16种, 其中真红树11种, 伴生植物5种, 除常见红树植物秋茄、桐花树、白骨壤和木榄外, 还包括有红海榄、海漆(Excoecaria agallocha)、榄李(Lumnitzera racemosa)、老鼠簕(Acanthus ilicifolius)、小花老鼠簕(Acanthus ebracteatus)等。
2 研究方法 2.1 样地设置与调查对广西北仑河口保护区红树林进行全面考察的基础上, 于2015年4—10月选取珍珠湾天然红树林分布相对集中区域, 依据红树植物的分布规律在高、中、低潮带随机建立20 m×20 m的固定样地12个(每个潮带3—5样地, 每个样地间隔50 m以上), 将每个样地划分为4个10 m×10 m的样方调查单元, 并使用PVC管每间隔5 m进行样方标记, 对样方内所有基径(板状根和支柱根上方1 cm处)≥1 cm的秋茄、桐花树和白骨壤个体以及胸径≥1 cm的木榄个体进行编号登记, 并记录其名称、高度、胸径、基径、相对坐标等指标, 对于基径或胸径<1 cm的幼苗只记录种名和高度, 同时记录每个样地地理坐标、滩涂类型、干扰程度和土壤性质等环境指标。
2.2 数据分析 2.2.1 高度级和大小级结构以1.0 m为一个高度级将秋茄、木榄、白骨壤和桐花树的高度结构划分为5级(Ⅰ级0.0—1.0 m, Ⅱ级1.0—2.0 m, Ⅲ级2.0—3.0 m, Ⅳ级3.0—4.0 m, Ⅴ级4.0—5.0 m), 分级统计株数。
虽然植物的径级或大小级不能直接表征种群的龄级特征, 但在相似的生境条件下, 龄级和大小级对环境的反应规律具有一定的一致性。因而, 在种群年龄难以准确获取的情况下, 多数学者采用径级或大小级代替龄级分析种群的结构和动态特征[28-29]。北仑河口4种红树植物的生长习性差异明显, 其中木榄为有独立主干的小乔木, 而秋茄、桐花树和白骨壤个体呈多萌枝的低矮灌木状。因此, 为合理地划分大小级, 以胸径划分木榄种群的大小级结构, 而其余3种以基径大小进行划分。由此, 本文对4种红树植物大小级的界定方法为:将基径<1 cm的幼苗, 按其高度划分为2级, Ⅰ级:0<高度<0.5 m、Ⅱ级:高度≥0.5 m, 对于基径≥1 cm的秋茄、桐花树和白骨壤个体, 基径每增加3 cm划分为1级, 合计划分为8级, 对于胸径≥1 cm的木榄个体, 胸径每增加3 cm划分为1级, 同样可划分为8级。
2.2.2 静态生命表与存活曲线参考魏新增等[27]对静态生命表的编制方法, 分别编制4种红树植物的种群静态生命表, 其参数包括:χ为大小级, 在静态生命表中替代年龄等级;Nχ为在χ大小级内出现的个体数;Lχ为标准化的存活数, Lχ=(Nχ+Nχ+1)/2;lgLχ为Lχ取以10为底的对数;Dχ为从χ到χ+1大小级的死亡数, Dχ=Nχ-Nχ+1;Qχ为从χ到χ+1大小级的死亡率, Qχ=Dχ/Nχ;T为从χ大小级起超过χ大小级的存活个体总数, T=∑Lχ;Eχ为生命期望, 表示第χ大小级的个体在未来所能存活的平均年数, Eχ=Tχ/Nχ。以χ为横坐标、lgLχ为纵坐标, 绘制4种红树植物种群的存活曲线。
2.2.3 时间序列分析时间序列分析在病虫害、树木生长等方面具有较高的预测准确性, 在植物种群动态预测方面也得到广泛采用并具有较高的预测能力和可靠性[30-32]。本文采用时间序列分析中的一次移动平均法对4种红树植物种群年龄结构进行预测, 其计测方法如下[31]:
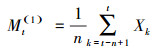
|
式中, n为需要预测的时间(本文为龄级时间), (1)表示1次移动平均, t代表龄级, Xk为k龄级内的个体数量, Mt(1)表示经过未来n个龄级时间后t龄级的种群大小。本文对经过2、4、6、8龄级时间后的种群各龄级的个体数量进行预测。
2.2.4 空间分布格局综合运用平均拥挤度(m*)、丛生指标(I)、聚块性指数(m*/m)、Cassie指标(CA)、扩散系数(C)、负二项参数(K)和格林指数(GI)7种聚集强度指数分析4种红树植物不同大小级种群的空间分布格局。其中, 平均拥挤度m*表示个体在样地中的邻体数, 表征个体的拥挤程度, 其数值越大表示聚集程度越高;丛生指标I<0时表示种群为均匀分布, C=0为随机分布, C> 0为聚集分布;聚块性指数m*/m<1时判定为逼近均匀分布, m*/m=1时为随机分布, m*/m> 1时为逼近聚集分布;Cassie指标CA<0时为均匀分布, CA=0为随机分布, CA> 0为聚集分布;扩散系数C<1时种群趋于均匀分布, C=1趋于随机分布, C> 1趋于聚集分布;负二项参数K<0时趋于均匀分布, K > 0时趋于聚集分布, 当K→+∞(一般为8以上)则逼近随机分布;格林指数GI值介于0—1之间, GI趋于0时表示逼近随机分布, 趋于1时为逼近最大聚集分布。7种格局强度指数的计算结果可进行对比分析, 以确保种群分布格局结果的准确性和可靠性。上述聚集强度指数的具体计测方法可参见文献[2]。
3 结果和分析 3.1 种群的高度结构调查表明, 12个样地仅出现秋茄、桐花树、白骨壤和木榄4种红树植物, 这4种红树植物也是北仑河口保护区红树林的优势种。高度结构分析表明(图 1), 4种红树植物的高度均在5.0m以下。其中, 白骨壤高度级Ⅰ级最少(仅2株), Ⅲ级最多(324株), 分别占个体总数的0.34%和54.65%。木榄Ⅰ级个体数最多(1911株), 占个体总数的64.60%, 其余Ⅱ、Ⅲ、Ⅳ、Ⅴ级株数分别为506株、377株、155株和9株, 分别占个体总数的17.11%、12.75%、5.24%和0.30%。秋茄高度级Ⅰ级(1291株)和Ⅲ级(673株)个体数较多, 占总数的54.96%和28.65%。桐花树高度级主要集中在Ⅰ级(835株)和Ⅱ级(1183株), 占比37.66%和53.36%。秋茄和桐花树种群都没有株高大于4.0 m的植株。上述结果表明, 4种红树植物高度结构中, 木榄种群呈现典型倒J形结构, 秋茄种群呈双峰型, 说明其幼苗和幼树较为丰富, 种群更新良好;桐花树种群更新受到一定阻力, 成年植株相对较少, 但能维持一定幼苗数, 种群相对稳定;白骨壤种群个体数相对较少且没有幼苗, 种群更新不良, 呈现较为明显的衰退趋势。

|
| 图 1 北仑河口4种红树植物种群的高度结构 Fig. 1 Population height structure of four mangrove plants in Beilun Estuary |
由大小级结构分析表明(图 2), 白骨壤、木榄和桐花树种群个体数量变化均存在一个峰值, 木榄Ⅱ级的个体数最多(1336株), 占个体总数的42.90%, 白骨壤和桐花树种群都是Ⅳ级的个体数最多(198株和791株), 分别占个体总数的33.33%和35.55%;而秋茄个体数的变化存在两个峰值, 分别是Ⅰ级(733株)和Ⅳ级(462株), 但是Ⅰ级的个体数要明显多于Ⅳ级的个体数。大小级结构比较完整的是木榄和秋茄种群, 而白骨壤和桐花树种群都存在不同的大小级缺失。与高度级结构相比, 大小级结构进一步表明, 北仑河口白骨壤种群属于典型衰退型种群, 桐花树种群有逐渐衰退趋势, 而木榄和秋茄种群属于增长型种群。

|
| 图 2 北仑河口4种红树植物种群的大小级结构 Fig. 2 Population size class structure of four mangrove plants in Beilun Estuary |
由静态生命表可看出(表 1), 白骨壤的Ⅰ、Ⅱ、Ⅲ级个体数均小于其后面一级, 导致出现死亡率为负值, 但从第Ⅳ级开始, 白骨壤的死亡率随着大小级的增加呈上升趋势, 在第Ⅵ级时出现一个高峰, 在第Ⅷ级时达到最大值, 这与存活数一栏第Ⅷ级白骨壤数量极少相符合。白骨壤第Ⅱ级生命期望最高, 随着龄级的增加生命期望值呈递减趋势。木榄的Ⅰ级个体数要小于其后一级, 秋茄的Ⅲ级个体数小于其后一级, 导致木榄Ⅰ级的死亡率和秋茄的Ⅲ级均出现负值, 但木榄从第Ⅱ级开始, 秋茄从第Ⅳ级开始, 死亡率也是随着龄级的增大而增加。桐花树的Ⅰ、Ⅱ、Ⅲ级个体数均小于其后面一级, 也导致出现死亡率为负值, 但与白骨壤不同的是, 桐花树的死亡率在第Ⅳ、Ⅴ、Ⅵ大小级相同均为0.9, 3个龄级的生命期望值也相近。种群静态生命表表明, 4种红树植物种群不同龄级的存活量差别较大, 存活量一般随龄级的增加逐渐减少。白骨壤和桐花树第Ⅰ、Ⅱ和Ⅲ级个体数均小于其后面一级, 导致死亡数和死亡率均出现较高的负值, 而木榄和秋茄幼年阶段的死亡率均较低。4种红树植物的生命期望往往在第Ⅰ、Ⅱ或Ⅲ级较高, 随后逐步递减。
| 种群 Population |
大小级 Size class χ |
存活数 Survival number Nχ |
标准化存活数 Standardized number of survivors Lχ |
标准化存活数对数 Logarithmic Lχ lgLχ |
死亡数 Standardized number of mortality Dχ |
死亡率 Mortality Qχ |
存活总数 Total number of survivors T |
生命期望 Life expectation Eχ |
| 白骨壤 A. marina |
Ⅰ | 0 | 1 | 0.0 | -2 | — | 594 | — |
| Ⅱ | 2 | 85 | 1.9 | -166 | -83.0 | 594 | 297.0 | |
| Ⅲ | 168 | 183 | 2.3 | -30 | -0.2 | 592 | 3.5 | |
| Ⅳ | 198 | 163.5 | 2.2 | 69 | 0.3 | 424 | 2.1 | |
| Ⅴ | 129 | 99.5 | 2.0 | 59 | 0.5 | 226 | 1.8 | |
| Ⅵ | 70 | 44.5 | 1.6 | 51 | 0.7 | 97 | 1.4 | |
| Ⅶ | 19 | 13.5 | 1.1 | 11 | 0.6 | 27 | 1.4 | |
| Ⅷ | 8 | 458.5 | 2.7 | -901 | -112.6 | 8 | 1.0 | |
| 木榄 B. gymnorrhiza |
Ⅰ | 909 | 1122.5 | 3.1 | -427 | -0.5 | 3114 | 3.4 |
| Ⅱ | 1336 | 935 | 3.0 | 802 | 0.6 | 2205 | 1.7 | |
| Ⅲ | 534 | 386.5 | 2.6 | 295 | 0.6 | 869 | 1.6 | |
| Ⅳ | 239 | 149.5 | 2.2 | 179 | 0.7 | 335 | 1.4 | |
| Ⅴ | 60 | 42.5 | 1.6 | 35 | 0.6 | 96 | 1.6 | |
| Ⅵ | 25 | 17.5 | 1.2 | 15 | 0.6 | 36 | 1.4 | |
| Ⅶ | 10 | 5.5 | 0.7 | 9 | 0.9 | 11 | 1.1 | |
| Ⅷ | 1 | 367 | 2.6 | -732 | -732.0 | 1 | 1.0 | |
| 秋茄 K. obovata |
Ⅰ | 733 | 663 | 2.8 | 140 | 0.2 | 2437 | 3.3 |
| Ⅱ | 593 | 395.5 | 2.6 | 395 | 0.7 | 1704 | 2.9 | |
| Ⅲ | 198 | 330 | 2.5 | -264 | -1.3 | 1111 | 5.6 | |
| Ⅳ | 462 | 367 | 2.6 | 190 | 0.4 | 913 | 2.0 | |
| Ⅴ | 272 | 188.5 | 2.3 | 167 | 0.6 | 451 | 1.7 | |
| Ⅵ | 105 | 71 | 1.9 | 68 | 0.6 | 179 | 1.7 | |
| Ⅶ | 37 | 37 | 1.6 | 0 | 0.0 | 74 | 2.0 | |
| Ⅷ | 37 | 164 | 2.2 | -254 | -6.9 | 37 | 1.0 | |
| 桐花树 A. corniculatum |
Ⅰ | 291 | 341.5 | 2.5 | -101 | -0.3 | 2226 | 7.6 |
| Ⅱ | 392 | 514.5 | 2.7 | -245 | -0.6 | 1935 | 4.9 | |
| Ⅲ | 637 | 714 | 2.9 | -154 | -0.2 | 1543 | 2.4 | |
| Ⅳ | 791 | 446 | 2.6 | 690 | 0.9 | 906 | 1.1 | |
| Ⅴ | 101 | 56.5 | 1.8 | 89 | 0.9 | 115 | 1.1 | |
| Ⅵ | 12 | 6.5 | 0.8 | 11 | 0.9 | 14 | 1.2 | |
| Ⅶ | 1 | 1 | 0.0 | 0 | 0.0 | 2 | 2.0 | |
| Ⅷ | 1 | 0.5 | -0.3 | 1 | 1.0 | 1 | 1.0 |
存活曲线一般分成3种类型, DeeveyⅠ型(凸型)表示种群中大多数个体均能达到其平均生理寿命, 在达到平均寿命时几乎同时死亡, DeeveyⅡ型(对角线型)表示种群中的各龄级个体具有相同的死亡率, DeeveyⅢ型(凹型)表示种群中的幼体死亡率较高, 以后龄级个体的死亡率低而稳定[33]。从图 3可看出, 白骨壤种群的存活曲线属于DeeveyⅠ型, 幼苗(Ⅰ级)存活率低, Ⅱ级开始存活率有所升高, Ⅲ级存活率最高, 之后存活率开始下降, 至Ⅷ级死亡率达到最大值。桐花树幼苗存活率偏低, 存活率最大值出现在Ⅲ级, 其存活曲线介于DeeveyⅠ型和DeeveyⅡ型之间。木榄和秋茄种群的存活曲线属DeeveyⅡ型, 木榄和秋茄的存活率的最大值均出现在幼苗阶段, 但是秋茄种群在Ⅳ级出现一个峰值, 而木榄种群的存活率是逐渐减小的。上述结果表明, 白骨壤种群个体均能达到其平均生理寿命, 在达到平均寿命后几乎同时死亡, 桐花树幼龄期死亡率偏高, 中龄期后死亡率逐渐接近, 而秋茄和木榄种群中的各龄级个体具有相对一致的死亡率。

|
| 图 3 北仑河口4种红树植物种群的存活曲线 Fig. 3 Survival curves of four mangrove plants in Beilun Estuary |
时间序列分析结果表明(表 2), 在经历2、4、6、8龄级之后, 白骨壤各龄级种群出现数量降低或先增加后降低的趋势, 例如, 第Ⅵ龄级由现在的70株增加到4龄级后的141株, 6龄级又降低至95株, 总的趋势是若干龄级后种群多度逐步降低;除Ⅱ龄级种群数量在2龄级后有所下降外, 木榄各龄级种群数量均呈现快速的增长, 如Ⅷ龄级种群仅1株, 8龄级后转变为389株;秋茄各龄级种群数量也均出现增长趋势, 但增长速度相对较缓;桐花树不同龄级种群出现不同的发展趋势, 其中Ⅱ、Ⅲ、Ⅳ龄级种群数量逐步降低, Ⅴ、Ⅶ、Ⅷ龄级种群逐步增加。由时间序列分析可看出, 白骨壤种群未来种群数量将明显降低, 种群逐步衰退;桐花树种群幼年个体数降低而成年个体数趋增, 在未来能维持一定的种群更新能力, 种群数量会表现为稳中有降;木榄和秋茄幼年个体较多, 各龄级种群补充良好, 因此未来种群数量将趋于增加。
| 种群 Population |
龄级 Age class |
原始数据 Primary data |
两龄级后的个体数 Survival number after two age class M2(1) |
四龄级后的个体数 Survival number after four age class M4(1) |
六龄级后的个体数 Survival number after six age class M6(1) |
八龄级后的个体数 Survival number after eight age class M8(1) |
| 白骨壤 A. marina |
Ⅰ | 0 | ||||
| Ⅱ | 2 | 1 | ||||
| Ⅲ | 168 | 85 | ||||
| Ⅳ | 198 | 183 | 92 | |||
| Ⅴ | 129 | 164 | 124 | |||
| Ⅵ | 70 | 100 | 141 | 95 | ||
| Ⅶ | 19 | 45 | 104 | 98 | ||
| Ⅷ | 8 | 14 | 57 | 99 | 74 | |
| 木榄 B. gymnorrhiza |
Ⅰ | 909 | ||||
| Ⅱ | 1336 | 1123 | ||||
| Ⅲ | 534 | 935 | ||||
| Ⅳ | 239 | 387 | 755 | |||
| Ⅴ | 60 | 150 | 542 | |||
| Ⅵ | 25 | 43 | 215 | 517 | ||
| Ⅶ | 10 | 18 | 84 | 367 | ||
| Ⅷ | 1 | 6 | 24 | 145 | 389 | |
| 秋茄 K. obovata |
Ⅰ | 733 | ||||
| Ⅱ | 593 | 663 | ||||
| Ⅲ | 198 | 396 | ||||
| Ⅳ | 462 | 330 | 497 | |||
| Ⅴ | 272 | 367 | 381 | |||
| Ⅵ | 105 | 189 | 259 | 394 | ||
| Ⅶ | 37 | 71 | 219 | 278 | ||
| Ⅷ | 37 | 37 | 113 | 185 | 305 | |
| 桐花树 A. corniculatum |
Ⅰ | 291 | ||||
| Ⅱ | 392 | 342 | ||||
| Ⅲ | 637 | 515 | ||||
| Ⅳ | 791 | 714 | 528 | |||
| Ⅴ | 101 | 446 | 480 | |||
| Ⅵ | 12 | 57 | 385 | 371 | ||
| Ⅶ | 1 | 7 | 226 | 322 | ||
| Ⅷ | 0 | 1 | 29 | 257 | 278 |
综合7种格局强度指数的计测结果表明(表 3), 除木榄Ⅷ级和桐花树Ⅶ级种群属随机分布外, 4种红树植物不同龄级种群均表现出较高的聚集强度, 不同龄级的聚集程度有所差异, 但总体表现为聚集强度随种群龄级的增加而逐渐降低。上述结果表明, 北仑河口红树林4种红树植物种群呈现明显的空间聚集分布, 这种聚集特征可能与其繁殖特性以及生境的异质性有关。
| 种群 Population |
大小级 Size class (χ) |
平均拥挤度 Mean crowding (m*) |
丛生指标 Clumping index (I) |
聚块性指数 Patchiness index (m*/m) |
Cassie指标 Cassie index (CA) |
扩散系数 Diffusing index (C) |
负二项参数 Negative bionmial parameter (K) |
格林指数 Green index (GI) |
结果 Result |
| 白骨壤 A. marina |
Ⅰ | — | — | — | — | — | — | — | — |
| Ⅱ | 1.04 | 1.00 | 25.00 | 24.00 | 2.00 | 0.04 | 0.01 | A | |
| Ⅲ | 19.89 | 16.39 | 5.68 | 4.68 | 17.39 | 0.21 | 0.17 | A | |
| Ⅳ | 16.93 | 12.81 | 4.11 | 3.11 | 13.81 | 0.32 | 0.13 | A | |
| Ⅴ | 10.64 | 7.95 | 3.96 | 2.96 | 8.95 | 0.34 | 0.08 | A | |
| Ⅵ | 4.92 | 3.46 | 3.37 | 2.37 | 4.46 | 0.42 | 0.03 | A | |
| Ⅶ | 1.95 | 1.55 | 4.92 | 3.92 | 2.55 | 0.26 | 0.02 | A | |
| Ⅷ | 1.04 | 0.87 | 6.23 | 5.23 | 1.87 | 0.19 | 0.01 | A | |
| 木榄 B. gymnorrhiza |
Ⅰ | 52.94 | 34.00 | 2.80 | 1.80 | 35.00 | 0.56 | 0.34 | A |
| Ⅱ | 78.28 | 50.45 | 2.81 | 1.81 | 51.45 | 0.55 | 0.51 | A | |
| Ⅲ | 32.73 | 21.60 | 2.94 | 1.94 | 22.60 | 0.52 | 0.22 | A | |
| Ⅳ | 20.07 | 15.09 | 4.03 | 3.03 | 16.09 | 0.33 | 0.15 | A | |
| Ⅴ | 4.42 | 3.17 | 3.54 | 2.54 | 4.17 | 0.39 | 0.03 | A | |
| Ⅵ | 1.15 | 0.63 | 2.22 | 1.22 | 1.63 | 0.82 | 0.01 | A | |
| Ⅶ | 0.43 | 0.22 | 2.04 | 1.04 | 1.22 | 0.96 | 0.00 | A | |
| Ⅷ | 0.02 | 0.00 | 1.00 | 0.00 | 1.00 | ∞ | 0.00 | R | |
| 秋茄 K. obovata |
Ⅰ | 51.09 | 35.82 | 3.35 | 2.35 | 36.82 | 0.43 | 0.36 | A |
| Ⅱ | 36.12 | 23.77 | 2.92 | 1.92 | 24.77 | 0.52 | 0.24 | A | |
| Ⅲ | 14.70 | 10.57 | 3.56 | 2.56 | 11.57 | 0.39 | 0.11 | A | |
| Ⅳ | 19.72 | 10.09 | 2.05 | 1.05 | 11.09 | 0.95 | 0.10 | A | |
| Ⅴ | 11.65 | 5.99 | 2.06 | 1.06 | 6.99 | 0.95 | 0.06 | A | |
| Ⅵ | 6.61 | 4.42 | 3.02 | 2.02 | 5.42 | 0.49 | 0.04 | A | |
| Ⅶ | 2.93 | 2.16 | 3.80 | 2.80 | 3.16 | 0.36 | 0.02 | A | |
| Ⅷ | 4.20 | 3.43 | 5.45 | 4.45 | 4.43 | 0.22 | 0.03 | A | |
| 桐花树 A. corniculatum |
Ⅰ | 27.26 | 21.20 | 4.50 | 3.50 | 22.20 | 0.29 | 0.21 | A |
| Ⅱ | 31.03 | 22.86 | 3.80 | 2.80 | 23.86 | 0.36 | 0.23 | A | |
| Ⅲ | 49.96 | 36.69 | 3.76 | 2.76 | 37.69 | 0.36 | 0.37 | A | |
| Ⅳ | 71.57 | 55.09 | 4.34 | 3.34 | 56.09 | 0.30 | 0.56 | A | |
| Ⅴ | 6.91 | 4.81 | 3.29 | 2.29 | 5.81 | 0.44 | 0.05 | A | |
| Ⅵ | 3.76 | 3.51 | 15.04 | 14.04 | 4.51 | 0.07 | 0.04 | A | |
| Ⅶ | 0.02 | 0.00 | 1.00 | 0.00 | 1.00 | ∞ | 0.00 | R | |
| Ⅷ | — | — | — | — | — | — | — | — | |
| 注:A:聚集分布Aggregated distribution;R:随机分布Random distribution | |||||||||
种群结构受到生物(如种内和种间竞争、病虫害和人为干扰等)与非生物(如土壤理化性质、光照等)诸多因素的共同影响, 分析种群结构有助于推断种群的动态过程及其潜在的生态学机制, 是揭示种群生存现状和更新策略的重要途径之一[34-36]。本研究基于高度和大小级结构、静态生命表、存活曲线以及时间序列预测等分析方法均较为一致地表明, 北仑河口4种红树植物中, 白骨壤种群属于明显的衰退型种群, 桐花树种群稳中有降, 而秋茄和木榄属于典型的增长型种群。有学者认为, 白骨壤种群主要分布于低潮带, 是红树群落演替的最前期先锋树种, 桐花树种群分布于低潮带和中潮带, 一般在红树林演替的中、前期阶段出现, 秋茄多分布于中潮带, 属于演替的中、后期常见种, 而木榄种群分布于高潮带, 属于典型的演替后期常见种[10, 37-38]。因此, 本研究中4种红树植物的种群更新动态结果很好地印证了上述观点, 表明红树植物种群动态与红树植物群落所处的演替阶段息息相关。先锋植物白骨壤种群处于衰退阶段, 桐花树种群相对稳定, 但由于幼苗增补不多而出现衰退征兆, 而相应地演替中后期的秋茄和木榄种群出现不同程度的增长, 表明本次调查范围内的红树林群落正处于演替的中期阶段。
红树植物种群的更新状况与其所处演替阶段的生境条件密切相关。白骨壤作为北仑河口红树林的先锋植物, 其隐胎生繁殖特性和独特的树形和广展的根系结构使其能较快占据新生裸地, 适应性强, 内、中、外滩均有分布, 形成郁闭的潮间带先锋群落[37]。先锋群落形成后, 消弱了风浪的侵蚀, 加速了海滩淤泥的累积, 滩泥化的土壤开始累积有机质, 盐度增加, 为适应泥质生境的其他红树植物定居创设了立地条件, 而此时土壤盐度和光照环境的改变致使先锋植物幼苗难以成功定居, 更新受阻[25, 37]。因此, 木榄、秋茄等因显胎生繁殖特性以及适应淤泥生境而逐步取得优势。范航清等[11]认为北仑河口红树林的演替规律为:老鼠簕、卤蕨和桐花树为优势种的群丛→白骨壤、桐花树为优势种的群丛→秋茄为优势种的群丛→木榄为优势种的群丛→海漆、黄瑾和银叶树为优势种的群丛。因此, 本研究结果有力地支持了这一观点, 即北仑河口保护区现有的白骨壤和白骨壤+桐花树群丛将逐步演替为以木榄为优势种的群丛, 如木榄+秋茄群丛、木榄+秋茄-桐花树群丛或木榄群丛。
除了生境条件外, 种内和种间竞争、虫害等生物因素也可能促进白红树植物种群出现不同程度的衰退。白骨壤先锋单优群落形成后, 植株的竞争力增强, 对空间、矿质营养和光照资源的需求相对增大, 导致种群内部为争夺光照、空间和矿质营养而发生激烈竞争, 种内竞争产生自疏作用。此外, 与桐花树、秋茄的种间竞争也会导致白骨壤个体死亡, 繁殖能力下降, 从而使存活率和更新能力逐渐降低。此外, 近十余年广西红树林几乎每年都不同程度地遭受病虫害危害, 导致红树植物生长受限, 严重情况下出现红树林大面积死亡[21]。因此, 虫害也是调控红树林植物种群动态的重要因素。例如, 广州小斑螟(Oligochroa cantoonella)专一性取食白骨壤的叶片, 对桐花树、秋茄和木榄却危害极少, 受害严重的白骨壤植株有90%以上的叶片干枯, 枝条或整株逐渐枯死[20]。另外, 白骨壤和桐花树是我国红树林的主体, 危害这2种植物的害虫达10余种, 因而白骨壤和桐花树遭受的虫害规模要远大于其他红树植物[38]。综上所述, 北仑河口白骨壤和桐花树种群出现不同程度的退化可能与单优群落种内和种间竞争作用加剧以及红树林虫害的发生有关, 但该推论还有待进一步验证。
4.2 红树植物的种群空间分布格局种群空间分布格局是种群对环境长期适应和选择的结果, 聚集分布与生境异质性、种子的扩散限制以及种内和种间竞争作用等有密切关系, 是陆地森林生态系统中最为普遍的植物种群空间分布格局类型[39-40]。本研究中, 4种红树植物不同发育阶段种群普遍表现为聚集分布, 聚集强度随种群龄级的增加而逐渐下降。已有研究表明, 澳门凼仔等地的桐花树、白骨壤和老鼠簕[41]、广东深圳福田的秋茄、桐花树和白骨壤种群[42]、福建漳江口的秋茄、桐花树和白骨壤[43], 这些植物种群均呈现空间聚集分布。此外, 海南东寨港的角果木(Ceriops tagal)种群在中潮带和内潮带表现为聚集分布[44], 广东湛江高桥的红海榄种群在中潮带或混交林中趋于聚集分布[45]。综上所述, 笔者认为, 受潮间带生境异质性(如, 盐度、微地形等)和植物自身生物和生态习性(如, 胎生现象等)的影响, 局域尺度上的聚集分布应该是我国海岸带红树植物种群空间分布格局的主要类型。
一般认为, 在较小尺度上, 种群空间分布格局可能由于不同程度的种内或种间竞争、种子扩散限制等生物因素所致, 而在大尺度上空间分布格局则可能是由物种分布的异质性以及土壤水分、养分与地形等非生物环境的异质性造成的[35-36, 39]。本研究中, 4种红树植物种群趋于聚集分布受其自身的生物生态学特性及所处的环境的双重影响。虽然白骨壤和桐花树为隐胎生植物, 轻且易于漂流的胎生苗有利于种群形成随机分布, 但无性系繁殖特性以及样地地形的异质性也有助于聚集格局的形成[46]。例如, 梁士楚[23]认为白骨壤种群的聚集分布与取样地点凸凹不平的微地形有关, 凹陷的地形会截留更多的胎生苗, 从而形成聚集分布。木榄和秋茄是显胎生红树植物, 其胎生幼苗成熟脱离母树后, 在其胚轴重力的作用下, 垂直下落后插入底泥中并迅速定植[47], 因而这种以母树为中心的胎生苗扩散限制特性大大提高了种群的空间聚集性。此外, 聚集强度随种群龄级的增加而逐渐下降是林木生长中普遍存在的现象[35-36]。随着4种红树林植物种群年龄的增大, 它们对资源的需求逐步增加, 种内或种间相互作用增强而造成密度制约性的自疏或它疏作用出现, 导致种群的个体数量递减, 从而降低了种群分布的聚集程度。
现有的关于红树林植物种群空间分布格局方面研究多是基于小尺度样地(如, 10 m×10 m或20 m×20 m样地)分析所得结果[41-45], 而种群空间分布格局分析结果的准确性与取样尺度和取样地点选取有密切关系, 不同的取样尺度和生境类型往往会产生不同的格局类型[39]。因此, 针对红树林种群空间分布格局方面研究要特别注重取样尺度和生境类型选取带来的影响。在红树林集中分布区建立大尺度的红树林固定样地, 如, 沿潮间带建立100 m×300 m样地(3hm2)或更大面积样地, 可以涵盖多种潮间带类型和异质生境, 并且可以基于点格局[48-49]、小波分析[50]等方法探讨时空尺度与格局的关系, 这将可以更为合理地解析红树植物种群结构并推断格局形成的生态学机制。因此, 笔者建议今后的红树林生态学研究中应尝试建立大尺度固定样地, 并进行包括动植物、微生物与环境的长期生态监测研究。
4.3 对红树林保护与抚育的建议近些年, 城市化、工业污染、生物入侵和病虫害等多重因素致使广西红树林多处于亚健康状态[11, 21]。作为弥补和改善天然红树林逐步退化的现状, 近些年人工红树林的营造面积不断扩大, 然而天然红树林及其长期形成的生态系统结构和功能往往是人工林系统无法比拟的[22, 51-52]。因此, 亟待保护和抚育物种多样性、生产力和生态效益相对较高的天然红树林。对现有天然红树林建立红树资源信息库和病虫害预警预报防控体系, 树立良好的信息搜集和反馈过程, 及时发现和处理各种导致天然红树林出现退化的生态问题。对由于干扰而处于衰退的种类, 采取人工抚育措施, 做好幼林初期的抚育和保护, 提高幼林的成活率和成林覆盖面积。同时加强红树人工林的种植和培育, 多层次、多角度提高红树自身抚育更新, 有效提高红树群落的健康生长。
退化或受损生态系统的恢复目标是其结构与功能恢复到接近干扰前的水平[3]。得益于人工红树林造林措施的实施, 近些年广西红树林分布面积保持相对稳定且出现一定幅度的增长趋势[15-16], 但红树林的人工恢复与重建需要以现有的天然红树林种群和群落结构特征为参照系, 研制出近自然的人工红树林林分结构和功能的优化配置和调控技术[53]。不同红树植物具有不同的生物学和生态学特性并适应于不同的生境类型。因此, 红树林生态恢复时需要遵循适地适树的原则, 分析恢复区的环境特征和生态限制因素, 在不同滩涂地带选取合适的造林树种和配置措施, 以乡土树种为主, 采取多树种混交和乔、灌结合的方式构建红树植物群落, 从而增强红树林空间结构复杂性和群落稳定性。
广西早期人工红树林造林以乡土灌木种桐花树和白骨壤为主, 近些年以小乔木种秋茄和外来树种无瓣海桑(Sonneratia apetala)为主[11]。对于早期营造的处于退化阶段的白骨壤和桐花树等先锋植物为主的低矮灌丛, 不能采用间伐、补植等改造措施继续栽植先锋红树植物, 而是参照红树林自然演替的规律, 引种相对高大的演替中、后期乔木树种, 如木榄和红海榄等, 使之形成符合演替规律的乔灌两层混交的林分结构, 从而提高群落的生物多样性、群落稳定性和生态效益。对于现有的红树林纯林(如无瓣海桑林、白骨壤林), 可参考天然红树林的物种配置模式, 采取间伐修枝和补植套种等措施增加林分中的乡土红树植物, 形成如无瓣海桑林+红海榄、白骨壤+秋茄等混交群落, 从而提升红树林的植物丰富度和林分的混交度, 达到降低病虫害危害程度和增强生态效益的目的。
| [1] | Harper J L. Population Biology of Plant. London:Oxford University Press, 1977. |
| [2] | 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. |
| [3] | 董鸣. 生态学透视——种群生态学. 北京: 科学出版社, 2016. |
| [4] | 刘贵峰, 臧润国, 丁易, 郭仲军, 张炜银, 成克武, 白志强, 张新平, 巴哈尔古丽·阿尤甫. 天山云杉种群结构的研究. 林业科学研究, 2011, 24(5): 659–662. |
| [5] | Galal T M. Size structure and dynamics of some woody perennials along elevation gradient in Wadi Gimal, Red Sea coast of Egypt. Flora——Morphology, Distribution, Functional Ecology of Plants, 2011, 206(7): 638–645. DOI:10.1016/j.flora.2010.11.010 |
| [6] | 卢杰, 郭其强, 郑维列, 徐阿生. 藏东南高山松种群结构及动态特征. 林业科学, 2013, 49(8): 154–160. DOI:10.11707/j.1001-7488.20130822 |
| [7] | Li W, Zhang G F. Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae). Flora——Morphology, Distribution, Functional Ecology of Plants, 2015, 212: 10–18. DOI:10.1016/j.flora.2015.02.002 |
| [8] | 廖宝文, 李玫, 陈玉军, 管伟. 中国红树林恢复与重建技术. 北京: 科学出版社, 2010. |
| [9] | 林鹏, 胡继添. 广西的红树林. 广西植物, 1983, 3(2): 95–102. |
| [10] | 梁士楚. 广西红树植物群落特征的初步研究. 广西科学, 2000, 7(3): 210–216. |
| [11] | 范航清, 黎广钊, 周浩郎. 广西北部湾典型海洋生态系统——现状与挑战. 北京: 科学出版社, 2015. |
| [12] | Li S S, Meng X W, Ge Z M, Zhang L Q. Vulnerability assessment of the coastal mangrove ecosystems in Guangxi, China, to sea-level rise. Regional Environmental Change, 2015, 15(2): 265–275. DOI:10.1007/s10113-014-0639-3 |
| [13] | 张忠华, 胡刚, 梁士楚. 广西红树林资源与保护. 海洋环境科学, 2007, 26(3): 275–279, 282. |
| [14] | 李春干, 代华兵. 1960-2010年广西红树林空间分布演变机制. 生态学报, 2015, 35(18): 5992–6006. |
| [15] | 但新球, 廖宝文, 吴照柏, 吴后建, 鲍达明, 但维宇, 刘世好. 中国红树林湿地资源、保护现状和主要威胁. 生态环境学报, 2016, 25(7): 1237–1243. |
| [16] | 张威, 陈正华, 王纪坤. 广西北部湾海岸带红树林变化的遥感监测. 广西大学学报:自然科学版, 2015, 40(6): 1570–1576. |
| [17] | 林鹏, 胡继添. 广西的红树植物群落. 植物学报, 1983, 25(1): 95–97. |
| [18] | 林鹏, 尹毅, 卢昌义. 广西红海榄群落的生物量和生产力. 厦门大学学报:自然科学版, 1992, 31(2): 199–202. |
| [19] | 范航清, 何斌源. 北仑河口的红树林及其生态恢复原则. 广西科学, 2001, 8(3): 210–214. |
| [20] | 范航清, 邱广龙. 中国北部湾白骨壤红树林的虫害与研究对策. 广西植物, 2004, 24(6): 558–562. |
| [21] | 范航清, 刘文爱, 曹庆天. 广西红树林害虫生物生态特性与综合防治技术研究. 北京: 科学出版社, 2012. |
| [22] | 范航清, 王欣, 何斌源, 潘良浩. 人工生境创立与红树林重建. 北京: 中国林业出版社, 2014. |
| [23] | 梁士楚. 广西北海海岸沙生白骨壤种群分布格局研究. 广西科学, 2001, 8(1): 57–60, 69. |
| [24] | 梁士楚. 红树植物木榄种群的分形特征. 广州: 中山大学, 2002. |
| [25] | 梁士楚, 刘镜法, 梁铭忠. 北仑河口国家级自然保护区红树植物群落研究. 广西师范大学学报:自然科学版, 2004, 22(2): 70–76. |
| [26] | 何斌源, 赖廷和, 王文卿, 陈剑锋, 邱广龙. 梯度淹水胁迫下全日潮海区秋茄幼苗的生长和生理反应. 海洋通报, 2007, 26(2): 42–49. |
| [27] | 魏新增, 黄汉东, 江明喜, 杨敬元. 神农架地区河岸带中领春木种群数量特征与空间分布格局. 植物生态学报, 2008, 32(4): 825–837. |
| [28] | Wu J X, Zhang X M, Deng C Z, Liu G J. Structure and dynamic of Populus euphratica population along Tarim River. Pakistan Journal of Botany, 2012, 44(5): 1651–1656. |
| [29] | 张亚芳, 李登武, 王梅, 刘盼. 黄土高原不同地区杜松种群结构与动态. 林业科学, 2015, 51(2): 1–10. |
| [30] | 张文辉, 王延平, 康永祥, 刘祥君. 濒危植物太白红杉种群年龄结构及其时间序列预测分析. 生物多样性, 2004, 12(3): 361–369. |
| [31] | 肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252–257. DOI:10.17521/cjpe.2004.0037 |
| [32] | 王立龙, 王亮, 张丽芳, 刘玉洋, 徐世健. 不同生境下濒危植物裸果木种群结构及动态特征. 植物生态学报, 2015, 39(10): 980–989. DOI:10.17521/cjpe.2015.0095 |
| [33] | 牛翠娟, 娄安如, 孙儒泳, 李庆芬. 基础生态学(第二版). 北京: 高等教育出版社, 2007. |
| [34] | Velázquez E, Kazmierczak M, Wiegand T. Spatial patterns of sapling mortality in a moist tropical forest:consistency with total density-dependent effects. Oikos, 2015, 125(6): 872–882. |
| [35] | 张忠华, 胡刚, 倪健. 茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征. 生态学报, 2015, 35(24): 8221–8230. |
| [36] | 王慧杰, 常顺利, 张毓涛, 谢锦, 何平, 宋成程, 孙雪娇. 天山雪岭云杉森林群落的密度制约效应. 生物多样性, 2016, 24(3): 252–261. DOI:10.17520/biods.2015282 |
| [37] | 缪绅裕, 陈桂珠, 李海生. 红树林植物桐花树和白骨壤及其湿地系统. 广州: 中山大学出版社, 2007. |
| [38] | 王友绍. 红树林生态系统评价与修复技术. 北京: 科学出版社, 2013. |
| [39] | Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414–1418. DOI:10.1126/science.288.5470.1414 |
| [40] | Zhang Z H, Hu G, Zhu J D, Ni J. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China. Journal of Plant Ecology, 2013, 6(2): 131–140. DOI:10.1093/jpe/rts027 |
| [41] | 梁华. 澳门红树林植物组成及种群分布格局的研究. 生态科学, 1998, 17(1): 25–31. |
| [42] | 李明顺, 蓝崇钰, 陈桂珠, 李世华, 陈相如, 刘治平, 谭凤仪, 黄玉山. 深圳福田红树林的群落学研究——Ⅱ.多样性与种群格局. 生态科学, 1994(1): 82–86. |
| [43] | 吴筠. 福建漳江口红树林数量特征及其退化机制研究. 福州: 福建农林大学, 2008. |
| [44] | 张育霞, 吕晓波, 杨小波, 李东海, 李英英, 罗文启, 熊梦辉, 邢莎莎, 朱静, 林泽钦, 岑举人, 郭照彬. 海南东寨港不同潮间带角果木种群结构及分布格局. 热带生物学报, 2015, 6(4): 388–396. |
| [45] | 周元满, 黄剑坚, 韩维栋, 杨淑贞. 天然红海榄种群分布格局研究. 广西植物, 2013, 33(4): 496–501. |
| [46] | 李小飞. 红树植物繁殖体密度与潮间带分布格局的关系. 厦门: 厦门大学, 2010. |
| [47] | 姚贻强, 李桂荣, 梁士楚. 广西防城港红树植物木榄种群结构的研究. 海洋环境科学, 2009, 28(3): 301–304. |
| [48] | 张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344–349. |
| [49] | Wiegand T, Moloney K A. Handbook of Spatial-Point-Pattern Analysis in Ecology. Boca Raton: CRC Press, 2014. |
| [50] | Dale M R T, Mah M. The use of wavelets for spatial pattern analysis in ecology. Journal of Vegetation Science, 1998, 9(6): 805–814. DOI:10.2307/3237046 |
| [51] | Ferreira A C, Ganade G, de Attayde J L. Restoration versus natural regeneration in a Neotropical mangrove:effects on plant biomass and crab communities. Ocean & Coastal Management, 2015, 110: 38–45. |
| [52] | Nam V N, Sasmito S D, Murdiyarso D, Purbopuspito J, Mackenzie R A. Carbon stocks in artificially and naturally regenerated mangrove ecosystems in the Mekong Delta. Wetlands Ecology and Management, 2016, 24(2): 231–244. DOI:10.1007/s11273-015-9479-2 |
| [53] | Chen L Z, Wang W Q, Zhang Y H, Lin G H. Recent progresses in mangrove conservation, restoration and research in China. Journal of Plant Ecology, 2009, 2(2): 45–54. DOI:10.1093/jpe/rtp009 |