 2018, Vol. 38
2018, Vol. 38文章信息
- 刘昊奇, 吕光辉.
- LIU Haoqi, LÜ Guanghui.
- 破碎景观建立保护区面积-数量模式界定
- Should several large or many small reserves be built in fragmented landscapes?
- 生态学报. 2018, 38(9): 3272-3280
- Acta Ecologica Sinica. 2018, 38(9): 3272-3280
- http://dx.doi.org/10.5846/stxb201704120640
-
文章历史
- 收稿日期: 2017-04-12
- 网络出版日期: 2018-01-04

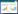




2. 新疆绿洲生态教育部重点实验室, 乌鲁木齐 830046;
3. 新疆大学干旱生态环境研究所, 乌鲁木齐 830046
2. Xinjiang Key Laboratory of Oasis Ecology, Ministry of Education, Urumqi 830046, China;
3. Institute of Arid Ecology and Environment, Xinjiang University, Urumqi 830046, China
由于动植物生存环境的改变, 很多物种的生存变得日益困难[1], 更严重的是大量珍稀野生动植物濒临灭绝, 如东北虎(Panthera tigris altaica)、大熊猫(Ailuropoda melanoleuca)和银杉(Cathaya argyrophylla)等, 因而保护濒危物种成为当前研究的热点。自然保护区作为保护濒危物种的一种有效工具[2-4], 引起了研究者的广泛关注。在设计自然保护区时, 有几个问题特别引人关注, 例如:应该将可利用的生境建设成少量大保护区还是大量小保护区?是否应在各保护区间建立廊道, 将孤立的保护区连接成网络[5-6]?第一个问题就是著名的FLOMS(few large or many small reserves, 少量大保护区还是大量小保护区)问题, 或称之为SLOSS(single large or several small reserves, 单一大保护区还是多个小保护区)问题, 本文针对此问题进行探究。
早在20世纪70年代中期, 研究者就提出了6条自然保护区的设计准则, 其中之一:当保护区面积固定时, 单一的大保护区模式更有助于物种生存[5, 7]。但随后这些结论受到了质疑, FLOMS问题随之成为研究的热点[8-11]。部分研究表明少量大保护区对物种生存有利[12-15]; 部分研究则认为大量小保护区更有利[10-11, 16-18]; 一些研究者认为中等程度的保护区数量最有利[19]; 还有研究者认为最优保护区数量随保护区环境承载量的增大而增加[20]。尽管已经有了大量研究结果, 但由于研究目标和方法的不同, 没有得出统一的结论, 综合所有的研究结果只能得到:最优保护区数量由物种自身特征和所处的环境决定, 例如:环境承载量、个体扩散和环境扰动[11, 19-20]。然而, 还有很多因子也可能对FLOMS问题有重要影响, 但之前的研究却未深入研究, 例如:生境丧失及其与个体扩散的共同作用。因而探讨这些因子可帮助研究者了解物种自身特征和所处的环境如何影响FLOMS问题, 并促进此问题的最终解决。
生境丧失不仅是物种灭绝的首要因素[21], 还造成现今很难找到一片未被破坏的景观来建设保护区, 因而生境丧失已成为影响保护区建设的重要因素。研究表明:生境丧失可通过作用于个体扩散、种群增长和环境扰动来影响FLOMS问题[20, 22-23]。个体扩散也可能对FLOMS问题有重要影响。个体扩散可通过“援救效应”帮助物种摆脱灭绝[24], 但个体扩散也可能带入疾病或寄生虫, 对物种造成负面影响[25]。关于个体扩散, 即使不考虑带入疾病和寄生虫, 也存在着截然相反的观点——大部分研究认为扩散对物种有利[26-27], 其他的则认为不利[28]。出现两个截然相反的结论的原因可能在于扩散过程中的个体死亡。若死亡率低, 扩散可通过“援救效应”帮助物种摆脱灭绝; 否则, 较高的死亡率会导致扩散带来的副面作用大于正面作用。
尽管已经有了如此多的研究, 但回顾之前的研究可发现几个缺点。其一, 尽管被破坏生境的空间分布方式是非随机的[29-30], 但之前的研究均假定其为随机的[20, 22-23]。其二, 个体扩散可能与生境丧失共同作用于物种, 从而影响FLOMS问题, 但涉及到该问题的研究很少。
1 模型模型描述根据Overview, Design concepts, Details protocol(ODD)[31-32]基于代理模型(agent-based models, ABM)的描述方式展开。
1.1 目的在一个已经遭受过生境丧失的景观上, 选取一片正方形的区域, 并调整此区域的面积以保证未被破坏的生境的面积为一个固定常数, 探讨生境丧失和个体扩散对FLOMS问题的影响。
1.2 载体和状态变量所选取的区域包含两种类型的生境:被破坏的生境和未被破坏的生境, 且所有未被破坏的生境将被建设成自然保护区。因此, 模型共有3种载体—被破坏的生境, 保护区和生存于保护区中的被保护物种。每个保护区对应一个环境最大可持续承载量和空间位置; 每个保护区都存在一个局部种群, 局部种群的某些个体可在保护区间扩散, 这些扩散的个体将各局部种群联系起来形成一个集合种群; 每个被破坏的生境也占据着一个空间位置。
1.3 过程综述一个离散时间和空间的模型被用于描述整个生态过程(图 1)。每一步:环境可持续承载量增长; 局部种群大小增长; 每个保护区都可能受到扰动的影响, 扰动率为μ; 局部种群中的某些个体可能扩散到相邻的保护区。如果在某一步, 被保护物种的个体总数少于a, 那么认为此物种的个体数量太少, 以至于已经生态灭绝, 并且终止整个过程。

|
| 图 1 过程, 次序安排和伪代码 Fig. 1 Processes, scheduling and pseudo-code |
因为可被建设成保护区的生境的面积是恒定常数, 所以所有保护区的总面积是恒定常数。尽管在现实中, 各保护区的面积、形状和环境可持续承载量以及各局部种群的大小不会完全相同, 忽略这些差异, 还假定各被破坏的生境的面积和形状与保护区相同。
依据被保护物种的灭绝概率决定建立大量小保护区还是少量大保护区, 最优保护区数量为被保护物种的灭绝概率达到最低时对应的保护区数量, 此时还可得到单个保护区的面积, 因为所有保护区的总面积是固定的常数, 据此可以决定建设大量小保护区还是少量大保护区。将未被破坏的生境划分成N2个面积相等的保护区(N2的取值范围从1变化到很多), 对于每个N2各进行1000次独立重复模拟实验, 并且从实验结果得到物种的灭绝概率。物种的灭绝概率定义为:在模拟过程中发生物种灭绝的那些模拟的次数与总模拟次数(1000)的比值。所以每个N2都对应一个物种灭绝概率, 据此可得知物种灭绝概率达到最低时N2的值, 这个值就是最优保护区数量。
在每一次模拟中, 从初始时刻到模拟终止时刻之间, 被保护物种个体数量的变动以及所对应的时间点都被记录。每个保护区受到扰动影响的概率都为μ, 所以扰动是一个随机发生的事件, 也是模型所包含的随机因素。
1.5 初始化使用元胞自动机模拟本文的模型, 元胞自动机的每个格子对应于一个保护区或被破坏的生境。为了保证保护区数量从1变化到很多, 设定元胞自动机有N1×N1(N1=2, 3, …, 31)个格子, 所以保护区的数量为N2=N1×N1×(1-D)(D为被破坏生境的数量与格子数量的比值。因为假定各保护区和各被破坏生境的面积都相同, 所以D也为所有被破坏生境的总面积与所选择区域的面积的比值, 即生境丧失比例), 被破坏生境的数量为N1×N1×D。因为每个保护区在初始时刻的环境可持续承载量都相同, 所以单个保护区在初始时刻的环境可持续承载量为KTOT/N2, KTOT为所有保护区在初始时刻的环境可持续承载量。因为生存于每个保护区上的局部种群的大小在初始时刻也相同, 所以假定初始时刻局部种群的大小为所在保护区环境承载量的一半——KTOT/(N2×2)。被破坏生境的位置由D和qE/E(从一个被破坏生境的邻居中随机挑一个, 其也为被破坏生境的概率, 也称之为聚集度[33])决定, 保护区占据剩余的位置。
1.6 子模型 1.6.1 自然保护区模型保护区共有N2=N1×N1×(1-D)(N1=2, 3, …, 31)个, 被破坏的生境共有N1×N1×D个。被破坏生境的位置由D和qE/E决定, 保护区占据剩余的位置。若被破坏生境的空间分布方式是随机的, 则在任一被破坏生境的邻居中随机选一个, 其也为被破坏生境的概率(qE/E)为D, 即D=qE/E; 若D>qE/E或D < qE/E, 则被破坏生境的空间分布方式是非随机的。从图 2可以看出聚集度qE/E越高, 被破坏生境的空间分布越集中, 保护区的分布也越集中。

|
| 图 2 两种保护区模式 Fig. 2 Two types of nature reserve models a—e:少量大保护区模式; f—j:大量小保护区模式; D=0.5;如1.4部分所述, 每一幅图中保护区(灰色格子)的总面积都相同且为一固定常数, 每幅图中所有保护区环境承载量的总和也相同。图中灰色格子为保护区, 白色格子为被破坏的生境, 黑线为各保护区或各被破坏生境的分界线 |
因为所有保护区的总面积为固定常数, 每个保护区或被破坏生境的面积都相同, 所以若保护区的总面积为S, 单个保护区或被破坏生境的面积都为S/N2, 且单个保护区的面积与保护区数量N2成反比。图 2为两种类型的保护区模式:少量大保护区模式和大量小保护区模式。
1.6.2 物种模型T时刻, 保护区I上的局部种群为NI(T), KI(T)为保护区I的环境可持续承载量。初始时刻, 所有保护区的环境可持续承载量为KTOT, K为单个保护区的环境可持续承载量。因为在初始时刻, 每个保护区的环境可持续承载量都相同, 所以K=KTOT/N2, N2为保护区数量。
种群增长:T时刻, 保护区I上的局部种群为[34]
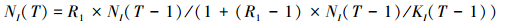
|
(1) |
R(R1≥1)为无竞争情况下, 局部种群的增长率; 而R1/(1+(R1-1)×NI(T-1)/KI(T-1))-1为局部种群的实际增长率。从方程(1)可得到:局部种群越接近环境承载量, 实际增长率越低; 当局部种群达到环境承载量时, 实际增长率为0;若其超过环境承载量, 种群会逐步减小。
环境可持续承载量增长:T时刻,保护区I的环境承载量为[34]
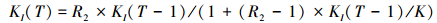
|
(2) |
R2(R2>1)为环境承载量的增长率。R2/(1+(R2-1)×KI(T-1)/K)-1为环境承载量的实际增长率, 其变动方式与局部种群实际增长率的变动方式类似, 但KI(T)≤K。
扰动:每个保护区受到扰动影响的概率都为μ, 扰动导致环境承载量降低和局部种群减小。T时刻, 如果保护区I受到扰动影响, 环境承载量和局部种群变化如下:

|
(3) |
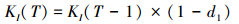
|
(4) |
d1(0 < d1 < 1)用来衡量扰动对局部种群和承载量的影响。
扩散:假设任一保护区中的个体只能扩散到其周围的保护区中, 且不能穿过被破坏的生境。任一保护区中的个体只能扩散到其周围八个邻居(摩尔邻居关系), 且其邻居必须也是保护区(图 3)。MIJ(T)为T时刻从保护区I扩散到保护区J中的个体。
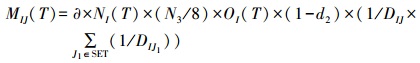
|
(5) |
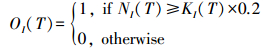
|
(6) |

∂为扩散率, d2为扩散死亡率, DIJ为保护区I的中心到保护区J的中心之间距离, N3为保护区I周围八个邻居中保护区的数量, SET为保护区I的八个邻居中保护区的集合(图 3, SET包含保护区A周围的5个保护区(绿色方格)), 所以SET中共有N3个元素。方程(5)说明:T时刻, 离开保护区I进行扩散的个体的数量为∂×NI(T)×(N3/8)×OI(T), 所以保护区I周围邻居中的保护区的数量(N3)越高, 选择扩散的个体越多; 保护区J的中心到I的中心距离(DIJ)越短, 1/DIJ越大, 因而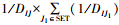
图 4展示随机或非随机的生境丧失对FLOMS问题的影响。表明:随机的生境丧失(D=qE/E)下, 生境丧失比例为0.2时, 最优保护区数量超过170;若生境丧失比例上升, 最优保护区数量将大幅减少, 当生境丧失比例上升到0.9时, 最优保护区数量已不足20。所以随机的生境丧失下, 生境丧失比例越高, 少量大保护区模式的优势越明显(保护区数量与各保护区的面积成反比)。

|
| 图 4 生境丧失比例和被破坏生境的聚集程度对最优保护区数量的影响 Fig. 4 The effect of proportion of lost habitats and the degree of clustering of lost habitats on the optimal reserve number 参数:R1=R2=1.013, kTOT=2000, μ=0.01, ∂=0.1, d1=0.5, d2=0, a=50, T=1100;聚集度: D=0.5 |
图 4b表明:若生境丧失比例恒定, 当聚集度为0.1时, 最优保护区数量不足20;聚集度提高, 最优保护区数量将大幅增加, 当聚集度提高到0.9时, 最优保护区数量已超过120。所以即使生境丧失比例保持不变, 被破坏生境的空间分布方式也会有重要影响-被破坏生境的空间聚集程度越高, 大量小保护区模式的优势越明显。
2.2 扩散(扩散率和扩散死亡率)对FLOMS问题的影响通过图 5, 探究生境丧失比例(D)恒定, 但被破坏生境的空间分布(用qE/E衡量)变化时, 扩散率和扩散死亡率对FLOMS问题的影响。扩散率为0时, 最优保护区数量非常少(不足20);当扩散率增加时, 最优保护区数量也增加。被破坏生境的聚集程度较高时(聚集度不小于0.7), 最优保护区数量的增幅更加明显; 即使被破坏生境的聚集程度非常低时(聚集度为0.2), 最优保护区数量也会增加, 但增幅较小。所以增加扩散率可导致从少量大保护区更有利于物种到大量小保护区的转变, 且被破坏生境的聚集程度越高, 转变的程度越高。

|
| 图 5 个体扩散率和个体的扩散死亡率对最优保护区数量的影响 Fig. 5 The optimal number of reserves changes with diffusion rate and diffusion mortality rate 参数:扩散率:D=0.5, R1=R2=1.012, kTOT=2000, μ=0.016, ∂=0.2, d1=0.5, a=50, T=1200;扩散死亡率:D=0.5, R1=R2=1.013, kTOT=2000, μ=0.01, d1=0.5, d2=0, a=50, T=1200 |
图 5还表明:当扩散死亡率较高时(0.9), 最优保护区数量很少; 若扩散死亡率降低, 最优保护区数量增加, 且被破坏生境的聚集程度较高时(聚集度不小于0.7), 最优保护区的增幅更加明显。所以降低扩散死亡可导致从少量大保护区更有利于物种到大量小保护区的转变, 且被破坏生境的聚集程度越高, 转变的程度越高。
3 实例作为“活化石”和“中国国宝”的大熊猫曾经广泛分布在华北、西北、华东、西南、华南以至越南和缅甸北部。但自近代以来, 由于人口急剧增加, 导致所需的土地越来越多, 很多大熊猫曾经的生境已转变为人类生活的城市和乡村, 剩余的生境也因为人类对于森林的过度砍伐而遭到严重破坏。因此, 大熊猫的数量急剧减小到不足1000只, 成为濒危物种。为保护大熊猫, 自卧龙大熊猫自然保护区创建以来, 中国已陆续建立起67个大熊猫保护区。保护措施取得了明显的成就, 一系列大范围国家调查显示, 此前的大熊猫数量下滑趋势已被扭转为稳定上升, 目前野外大熊猫的数量已上升到2000只左右。但随着保护区周围地区经济的发展, 城市和乡村不断扩大, 也由于交通的发展, 公路和铁路密度越来越大, 这些都造成了大熊猫生境的进一步丧失和空间聚集程度进一步降低。为了应对这一变化, 国家决定将所有67个大熊猫保护区整合为由岷山、邛崃山——大相岭、秦岭和白水江这4个片区组成的大熊猫公园, 以加强以大熊猫为核心的生物多样性保护。大熊猫生境生境丧失程度的增加和空间聚集程度的降低, 导致现今近70个保护区将被整合为4个, 这成为证明“生境丧失比例增加或被破坏生境的空间聚集程度降低时, 少量大保护区更优”的现实例子。
4 讨论生境丧失不仅造成很多珍稀的动植物濒临灭绝, 还造成其所在的生境破碎化[35-36]。虽然可以借助生态技术帮助动植物恢复部分被破坏的生境[37-38], 但要恢复所有生境, 并任意选择所需的生境建立自然保护区, 已没有可能。因而在建立保护区时, 必须处理生境丧失所带来的问题。本文选择其中的部分问题进行探讨, 在一个已经遭受过生境丧失的景观上, 选取一片区域, 并保证此区域中未被破坏生境的面积为一个固定常数, 然后探讨应该将未被破坏生境设成少量大保护区还是大量小保护区。结果表明:(1)生境丧失比例和被破坏生境的空间分布形式共同影响FLOMS问题——若被破坏生境的空间分布形式保持不变, 生境丧失比例越高, 少量大保护区模式的优势越明显; 若生境丧失比例恒定, 被破坏生境的空间聚集程度越高, 大量小保护区模式的优势越明显。(2)生境丧失还对FLOMS问题存在间接影响, 扩散率增加或扩散死亡率降低可导致从少量大保护区更有利于物种到大量小保护区最优的转变, 且被破坏生境的聚集程度越高, 转变程度越剧烈。
之前的研究表明生境丧失会通过对其他因子(种群增长率、扰动或环境承载量)产生作用来影响FLOMS问题[20, 22]。例如:当被破坏的生境随机分布在景观上时, Reed[23]的研究表明, 种群增长率越高, 越适宜采用大量小保护区模式。本文的部分研究也考虑生境丧失通过对其他因子(个体扩散)产生作用来影响FLOMS问题, 但之前的研究均假设被破坏的生境随机分布在景观上, 而相关研究表明被破坏生境的分布方式是非随机的[29-30], 因而这些假设与实际不符。此外, 这些研究也仅涉及到生境丧失对FLOMS问题的间接作用, 而没有探讨直接作用。本文解决了以上问题, 结果表明:不但生境丧失比例直接对FLOMS问题造成影响, 而且即使生境丧失比例保持不变, 被破坏生境的空间分布特征对FLOMS问题也造成影响。所以, 在探讨生境丧失时, 必须同时考虑生境丧失比例和被破坏生境的空间分布特征(即:生境丧失的空间特征), 尤其是后者, 因为以前的研究往往重视前者, 而忽视后者[20, 22-23]。本文的研究结果还表明:被破坏生境的聚集程度还会影响个体扩散(扩散率和扩散死亡率)对FLOMS问题的作用——被破坏生境的聚集度较高时, 保护区最优数量对个体扩散的反应更剧烈。
本文基于元胞自动机模型, 研究生境丧失和个体扩散的共同作用对自然保护区设计中关键问题——FLOMS问题——的影响。研究结果可为生物多样性保护和保护区设计提供理论基础, 有助于人们加深对物种如何响应生境丧失的了解, 增强人们对生境破坏的重视程度, 提高人们保护自然环境的意识。但个体不仅能短距离扩散到邻近地区, 还可以借助廊道, 长距离扩散到相距较远的区域; 生境丧失可能与短距离扩散和长距离扩散共同作用于种群动态[39], 从而影响影响FLOMS问题。但这些本文都没涉及, 此外本文还存在需要改进的问题, 虽然很多自然保护区都有其主要保护的物种, 如中国的大熊猫自然保护区, 但自然保护区内一般会有多个需要保护的物种, 因此在设计自然保护区时, 除了考虑其主要保护的对象外, 还必须兼顾所有需要保护的物种, 对生境丧失如何影响FLOMS问题的研究还需深入讨论。
| [1] | 陈成忠, 葛绪广, 孙琳, 邵冬, 柯文莉. 物种急剧丧失·生态严重超载·跨越"地球边界"·区域公平失衡·"一个地球"生活——《地球生命力报告2014》解读. 生态学报, 2016, 36(9): 2779–2785. |
| [2] | Soule M E. Conservation:tactics for a constant crisis. Science, 1991, 253(5021): 744–750. DOI:10.1126/science.253.5021.744 |
| [3] | Van Cuong C, Dart P, Dudley N, Hockings M. Factors influencing successful implementation of biosphere reserves in vietnam:challenges, opportunities and lessons learnt. Environmental Science & Policy, 2017, 67: 16–26. |
| [4] | Gong M H, Fan Z Y, Wang J Y, Liu G, Lin C. Delineating the ecological conservation redline based on the persistence of key species:giant pandas (Ailuropoda melanoleuca) inhabiting the Qinling mountains. Ecological Modelling, 2017, 345: 56–62. DOI:10.1016/j.ecolmodel.2016.11.011 |
| [5] | Diamond J M. The island dilemma:lessons of modern biogeographic studies for the design of natural reserves. Biological Conservation, 1975, 7(2): 129–146. DOI:10.1016/0006-3207(75)90052-X |
| [6] | Wilson E O, Willis E O. Applied biogeography//Cody M L, Diamond J M, eds. Ecology and Evolution of Communities. Cambridge, Massachusetts: Belknap Press, 1975: 522-534. |
| [7] | May R M. Islands biogeography and the design of wildlife preserves. Nature, 1975, 254(5497): 177–178. DOI:10.1038/254177a0 |
| [8] | Gilpin M E, Diamond J M. Subdivision of nature reserves and the maintenance of species diversity. Nature, 1980, 285(5766): 567–568. DOI:10.1038/285567a0 |
| [9] | Groeneveld R. Economic considerations in the optimal size and number of reserve sites. Ecological Economics, 2005, 52(2): 219–228. DOI:10.1016/j.ecolecon.2004.06.023 |
| [10] | Baker T P, Jordan G J, Dalton P J, Baker S C. Impact of distance to mature forest on the recolonisation of bryophytes in a regenerating Tasmanian wet eucalypt forest. Australian Journal of Botany, 2013, 61(8): 633–642. DOI:10.1071/BT13250 |
| [11] | Lindenmayer D B, Wood J, McBurney L, Blair D, Banks S C. Single large versus several small:the SLOSS debate in the context of bird responses to a variable retention logging experiment. Forest Ecology and Management, 2015, 339: 1–10. DOI:10.1016/j.foreco.2014.11.027 |
| [12] | Etienne R S, Heesterbeek J A P. On optimal size and number of reserves for metapopulation persistence. Journal of Theoretical Biology, 2000, 203(1): 33–50. DOI:10.1006/jtbi.1999.1060 |
| [13] | Burkey T V. Metapopulation extinction in fragmented landscapes:using bacteria and protozoa communities as model ecosystems. The American Naturalist, 1997, 150(5): 568–591. DOI:10.1086/286082 |
| [14] | Tjørve E. How to resolve the SLOSS debate:lessons from species-diversity models. Journal of Theoretical Biology, 2010, 264(2): 604–612. DOI:10.1016/j.jtbi.2010.02.009 |
| [15] | McCarthy M A, Thompson C J, Moore A L, Possingham H P. Designing nature reserves in the face of uncertainty. Ecology Letters, 2011, 14(5): 470–475. DOI:10.1111/ele.2011.14.issue-5 |
| [16] | Boecklen W J. Nestedness, biogeographic theory, and the design of nature reserves. Oecologia, 1997, 112(1): 123–142. DOI:10.1007/s004420050292 |
| [17] | Skaggs R W, Boecklen W J. Extinctions of montane mammals reconsidered:putting a global-warming scenario on ice. Biodiversity & Conservation, 1996, 5(6): 759–778. |
| [18] | Le Roux D S, Ikin K, Lindenmayer D B, Manning A D, Gibbons P. Single large or several small? Applying biogeographic principles to tree-level conservation and biodiversity offsets. Biological Conservation, 2015, 191: 558–566. DOI:10.1016/j.biocon.2015.08.011 |
| [19] | Ovaskainen O. Long-term persistence of species and the SLOSS problem. Journal of Theoretical Biology, 2002, 218(4): 419–433. DOI:10.1016/S0022-5193(02)93089-4 |
| [20] | Robert A. The effects of spatially correlated perturbations and habitat configuration on metapopulation persistence. Oikos, 2009, 118(10): 1590–1600. DOI:10.1111/oik.2009.118.issue-10 |
| [21] | Saunders D A, Hobbs R J, Margules C R. Biological consequences of ecosystem fragmentation:a review. Conservation Biology, 1991, 5(1): 18–32. DOI:10.1111/cbi.1991.5.issue-1 |
| [22] | Pelletier J D. Model assessments of the optimal design of nature reserves for maximizing species longevity. Journal of Theoretical Biology, 2000, 202(1): 25–32. DOI:10.1006/jtbi.1999.1030 |
| [23] | Reed D H. Extinction risk in fragmented habitats. Animal Conservation, 2004, 7(2): 181–191. DOI:10.1017/S1367943004001313 |
| [24] | Richards C M. Inbreeding depression and genetic rescue in a plant metapopulation. The American Naturalist, 2000, 155(3): 383–394. DOI:10.1086/303324 |
| [25] | Harding K C, McNamara J M. A unifying framework for metapopulation dynamics. The American Naturalist, 2002, 160(2): 173–185. DOI:10.1086/341014 |
| [26] | Gonzalez A, Lawton J H, Gilbert F S, Blackburn T M, Evans-Freke I. Metapopulation dynamics, abundance, and distribution in a microecosystem. Science, 1998, 281(5385): 2045–2047. DOI:10.1126/science.281.5385.2045 |
| [27] | Debinski D M, Holt R D. A survey and overview of habitat fragmentation experiments. Conservation Biology, 2000, 14(2): 342–355. DOI:10.1046/j.1523-1739.2000.98081.x |
| [28] | Quinn J F, Hastings A. Extinction in subdivided habitats. Conservation Biology, 1987, 1(3): 198–208. DOI:10.1111/cbi.1987.1.issue-3 |
| [29] | Boakes E H, Mace G M, McGowan P J K, Fuller R A. Extreme contagion in global habitat clearance. Proceedings of the Royal Society B:Biological Sciences, 2010, 277(1684): 1081–1085. DOI:10.1098/rspb.2009.1771 |
| [30] | Xie Y J, Ng C N. Exploring spatio-temporal variations of habitat loss and its causal factors in the Shenzhen river cross-border watershed. Applied Geography, 2013, 39: 140–150. DOI:10.1016/j.apgeog.2013.01.001 |
| [31] | Grimm V, Berger U, Bastiansen F, Eliassen S, Ginot V, Giske J, Goss-Custard J, Grand T, Heinz S K, Huse G, Huth A, Jepsen J U, Jørgensen C, Mooij W M, Müller B, Pe'er G, Piou C, Railsback S F, Robbins A M, Robbins M M, Rossmanith E, Rüger N, Strand E, Souissi S, Stillman R A, Vabø R, Visser U, DeAngelis D L. A standard protocol for describing individual-based and agent-based models. Ecological Modelling, 2006, 198(1/2): 115–126. |
| [32] | Grimm V, Berger U, DeAngelis D L, Polhill J G, Giskee J, Railsback S F. The ODD protocol:a review and first update. Ecological Modelling, 2010, 221(23): 2760–2768. DOI:10.1016/j.ecolmodel.2010.08.019 |
| [33] | Hiebeler D. Populations on fragmented landscapes with spatially structured heterogeneities:landscape generation and local dispersal. Ecology, 2000, 81(6): 1629–1641. DOI:10.1890/0012-9658(2000)081[1629:POFLWS]2.0.CO;2 |
| [34] | Johst K, Drechsler M. Are spatially correlated or uncorrelated disturbance regimes better for the survival of species?. Oikos, 2003, 103(3): 449–456. DOI:10.1034/j.1600-0706.2003.12770.x |
| [35] | De Montis A, Martín B, Ortega E, Ledda A, Serra V. Landscape fragmentation in Mediterranean Europe:a comparative approach. Land Use Policy, 2017, 64: 83–94. DOI:10.1016/j.landusepol.2017.02.028 |
| [36] | 陈利顶, 刘雪华, 傅伯杰. 卧龙自然保护区大熊猫生境破碎化研究. 生态学报, 1999, 19(3): 291–297. |
| [37] | 刘春霞, 韩烈保. 高速公路边坡植被恢复研究进展. 生态学报, 2007, 27(5): 2090–2098. |
| [38] | 刘二佳, 张晓萍, 谢名礼, 陈妮, 张亭亭, 郭敏杰, 张建军. 生态恢复对流域水沙演变趋势的影响——以北洛河上游为例. 生态学报, 2015, 35(3): 622–629. |
| [39] | Hiebeler D. Competition between near and far dispersers in spatially structured habitats. Theoretical Population Biology, 2004, 66(3): 205–218. DOI:10.1016/j.tpb.2004.06.004 |