 2018, Vol. 38
2018, Vol. 38文章信息
- 李晓雪, 贺燕燕, 杨文航, 王朝英, 燕江伟, 崔振, 李昌晓.
- LI Xiaoxue, HE Yanyan, YANG Wenhang, WANG Chaoying, YAN Jiangwei, CUI Zhen, LI Changxiao.
- 不同水分处理和密度配置对牛鞭草与狗牙根生长与种间竞争的影响
- Effects of different water conditions and plant density on the growth and interspecific competition of Hemarthria compressa and Cynodon dactylon
- 生态学报. 2018, 38(9): 3046-3058
- Acta Ecologica Sinica. 2018, 38(9): 3046-3058
- http://dx.doi.org/10.5846/stxb201703140427
-
文章历史
- 收稿日期: 2017-03-14
- 网络出版日期: 2018-01-26

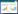




三峡水库蓄水后, 水位从145 m到175 m周期性变化[1-2]。有研究表明, 大幅度异常水位变化会使消落带生态功能退化[3], 很多原有植物无法生存, 从而造成库区水土严重流失、库区景观质量下降等生态环境问题[4]。采用构建植被对库岸带进行恢复是合理且有效的对策[5]。草本植物因其易生存、易扩张的特性成为三峡库区消落带生态恢复的先锋物种。长期以来, 关于三峡库区消落带草本植被的保护及合理利用、退化草本植被的生态修复以及经营管理都十分落后。与此同时, 对乡土优势草种在不同水淹条件下的生态适应性及竞争关系认识不足也是导致消落带草本植被退化和面积难以扩大的一个重要原因。竞争对塑造植物个体形态、生活史和种群驯化具有重要的影响作用, 同时它是决定植物群落组成、结构与动态特征的主要动力[6-7]。在三峡库区消落带, 水淹是引起植物竞争发生变化的胁迫因子, 而植物竞争能力是植物适应胁迫环境的重要指标[8]。在三峡库区消落带特定环境条件下, 为有效恢复消落带草本植被而进行人工草地建植时, 合理控制消落带混植草种比例是调节种内竞争和种间竞争的有效途径[9]。然而, 不同混植比例下乡土草种对消落带水淹的生态适应性如何、以及种内种间竞争如何变化等科学问题均不得而知。另外, 消落带不同水淹条件下哪种混植比例最适于乡土草种的生长也未见相关报道。解决这些问题是建立和恢复三峡库区消落带草地生态系统功能的关键和捷径。
狗牙根(Cynodon dactylon)与牛鞭草(Hemarthria compressa)为禾本科多年生草本植物[10-11], 二者均具有克隆习性, 能较快扩张占领生境, 在长江中上游地区牛鞭草常与狗牙根相伴而生[12], 可以利用狗牙根和牛鞭草进行三峡库区消落带草本植被重建。尽管有研究[13-14]对三峡库区消落带狗牙根和牛鞭草混种后的种间竞争进行了探讨, 但只是针对单一密度配置开展研究, 所选比例可能会影响竞争结果, 并无法预测其他密度配置下的竞争情况。因此, 本试验以狗牙根和牛鞭草为试验材料, 采用取代试验法, 按一定的梯度设置密度配置, 对两种植物在不同水淹和密度配置下的生长及种间竞争进行了研究, 可以在一定程度上弥补单一密度配置的弊端, 进而为利用优良草种进行三峡库区消落带植被恢复提供理论依据。
1 材料和方法 1.1 试验材料本试验选择狗牙根、牛鞭草当年生扦插苗作为研究对象, 采用盆栽控制试验, 盆钵规格为:高25 cm, 内径30 cm。2016年4月29日开始进行狗牙根、牛鞭草的扦插栽培, 种植时先将狗牙根截成10 cm左右带有3个芽孢的小段, 牛鞭草截成10 cm左右含有3个节的小段, 按实验设计比例进行种植。之后将所有盆栽试验用苗置于西南大学生态试验园的遮雨棚下(棚顶透明, 四面敞开)进行相同条件的适应生长, 并给予除草等常规管理。于2016年6月20日开始进行试验处理, 此时的狗牙根、牛鞭草平均株高与基茎如表 1所示。
| 密度配置 Density configuration |
牛鞭草 Hemarthria compressa | 狗牙根 Cynodon dactylon | ||
| 株高 Plant height/cm |
基茎 Stem diameter/mm |
株高 Plant height/cm |
基茎 Stem diameter/mm |
|
| H0C12 | 69.77±7.11 | 1.28±0.16 | ||
| H2C10 | 31.45±2.55 | 3.03±0.22 | 87.45±11.48 | 0.99±0.03 |
| H4C8 | 43.06±7.38 | 2.62±0.17 | 74.97±7.62 | 0.99±0.08 |
| H6C6 | 45.83±5.26 | 2.74±0.10 | 88.52±12.81 | 0.85±0.02 |
| H8C4 | 46.82±3.85 | 2.62±0.07 | 112.30±4.25 | 0.93±0.14 |
| H10C2 | 45.53±3.37 | 2.34±0.16 | 89.07±12.46 | 0.85±0.10 |
| H12C0 | 36.64±2.67 | 2.43±0.11 | ||
本研究设置3个水分处理, 分别为对照组(CK)——正常供水, 土壤含水量为田间持水量的60%—63%、浅淹组(SF)——水淹超过土壤表面10 cm和全淹组(TF)——水淹超过土壤表面2 m。密度配置采用取代系列试验法[15], 每盆种植12株植物(相当于种植密度为195株/m2), 设置7种配置比例, 每盆牛鞭草与狗牙根株数分别按2株进行递增与递减, 具体的配比分别为:H0C12, H2C10, H4C8, H6C6, H8C4, H10C2, H12C0, 采用完全随机区组实验设计, 盆内种植示意图如图 1。

|
| 图 1 牛鞭草与狗牙根栽植示意图 Fig. 1 The planting diagram of Hemarthria compressa and Cynodon dactylon |
从试验处理第一天开始, 每天对试验材料进行观察, 并确保各处理组保持设定的土壤含水量。处理30 d[16]后对各项指标进行测定, 每个处理3次重复。
1.3 测定指标及方法 1.3.1 生物量生物量的计算方法为:单株生物量=盆中狗牙根或牛鞭草的总生物量/盆中狗牙根或牛鞭草的株数。取样时将植株的根、茎、叶分开, 放置于80℃烘箱中烘干至恒重。地上生物量干重=总叶生物量+茎生物量, 总生物量干重=地上生物量干重+根生物量干重, 根冠比=地下生物量干重/地上生物量干重。
1.3.2 生长及形态指标生长形态指标包括主茎长、基茎、分枝数、总根长、根直径。测定植株的主茎长用卷尺测量, 基茎用游标卡尺测量。通过WinRHIZO La2400根系扫描仪扫描根图像。
1.4 数据处理和分析植物种间竞争能力的强弱通常用竞争系数来进行比较和衡量, 竞争系数通常要与其特定的试验设计方法相互匹配[17-18]。在替代系列试验中常用的竞争系数有相对总生物量(Relative yield total, RYT), 相对竞争强度(Relative competition intensity, RCI), 竞争攻击系数(Aggressivity, A)[19]。
(1) 相对总生物量(Relative yield total, RYT)是一个用于评价间作体系中生物学效益的竞争系数, 当RYT>1时, 表明物种a、b占有不同的生态位且表现出共生关系; RYT=1时表明物种a、b利用共同的资源; RYT < 1时表明物种a、b间存在相互拮抗和竞争关系。其计算公式如下:
RYT=(RYab+RYba)=Yab/Yaa+Yba/Ybb
式中, Yab为混植牛鞭草的生物量, Yaa为单植牛鞭草的生物量; Yba为混植狗牙根的生物量, Ybb为单植狗牙根的生物量。
(2) 相对竞争强度(Relative competition intensity, RCI), 用于比较混植体系中各自物种的竞争能力。当0 < RCI < 1时, 随着RCI越大, 说明该草种在混植草地中的产量越低, 另一草种对它的影响越大, 在混植群落中竞争能力较差; 当RCI=1时, 该草种已经被排出系统; 当RCI=0时, 说明混植的草种对另一个草种在产量上没有影响。若RCI < 0, 说明一个草种的存在有利于另一个草种的生长, 并促进其产量增加。其计算公式为:
RCI=(Yaa-Yab)/Yaa或RCI=(Ybb-Yba)/Ybb
式中, Yaa为单植牛鞭草的生物量, Yab为混植牛鞭草的生物量; Ybb为单植狗牙根的生物量, Yba为混植狗牙根的生物量。
(3) 竞争攻击系数(Aggressivity, A)是评价混植体系中各自物种的竞争能力的指标, 当Aa>0时, 表明物种a竞争能力大于物种b, 且处于主导优势地位; Aa=0时, 表明物种a、b竞争能力相同; Aa < 0时, 表明物种a的竞争能力小于物种b。其计算公式为:
Aab=Yab/(Yaa×Zab)-Yba/(Ybb×Zba)
式中, Yab为混植牛鞭草的生物量, Yaa为单植牛鞭草的生物量; Yba为混植狗牙根的生物量, Ybb为单植狗牙根的生物量。式中Z代表混植体系中各自物种所占的比例, 即Zab+Zba=1。
利用SPSS 18.0软件双因素方差分析(two-way ANOVA)进行数据分析, 并运用Duncan检验法检验不同处理的各个指标差异显著性。利用软件Origin 8.5绘图, Microsoft world制表。
2 结果 2.1 水淹胁迫和密度配置对狗牙根与牛鞭草的影响 2.1.1 水淹胁迫和密度配置对狗牙根与牛鞭草形态及生长的影响各处理条件下, 狗牙根存活率达100%。水分处理显著影响狗牙根的株高和总根长(表 2)。浅淹条件下狗牙根的平均株高高于CK组(P < 0.05), 而全淹组狗牙根的平均株高低于CK组(P < 0.05)。狗牙根的总根长随着水淹深度的加深呈下降趋势(图 2)。各处理条件下, 牛鞭草存活率达100%。水分处理极显著地影响牛鞭草各形态指标(P < 0.01)(表 3)。与CK组相比, 浅淹组牛鞭草的平均株高增加, 全淹组的平均株高降低; 牛鞭草的基茎和根长变化趋势与株高相类似; 分枝数在各比例条件下随着水淹深度的加深呈下降趋势; 各比例处理下, 全淹组的根直径大于浅淹组的根直径(图 3)。
| 指标Index | 水分处理Water treatment | 密度配置Density configuration | 交互作用Interaction | |||||
| F | P | F | P | F | P | |||
| 株高Plant height | 4.8 | 0.014 | 20.1 | <0.001 | 6.8 | <0.001 | ||
| 基茎Stem diameter | 3.1 | 0.052 | 2.8 | 0.021 | 2.0 | 0.048 | ||
| 分枝数No. of branches | 1.3 | 0.521 | 13.9 | <0.001 | 11.9 | <0.001 | ||
| 根长Total root length | 6.5 | 0.002 | 0.4 | 0.834 | 2.3 | 0.006 | ||
| 根直径Diameter | 1.4 | 0.256 | 4.5 | 0.001 | 1.2 | 0.281 | ||

|
| 图 2 水分处理及密度配置对狗牙根形态及生长的影响(平均值±标准误, n=3) Fig. 2 Effects of water treatments and density configurations on morphology and growth of Cynodon dactylon (mean±SE, n=3) CK:对照, Control; SF:浅淹, Shallow flooding; TF:深淹, Total flooding; 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) |
| 指标Index | 水分处理Water treatment | 密度配置Density configuration | 交互作用Interaction | |||||
| F | P | F | P | F | P | |||
| 株高Plant height | 82.6 | <0.001 | 5.9 | <0.001 | 6.0 | <0.001 | ||
| 基茎Stem diameter | 12.5 | <0.001 | 9.4 | <0.001 | 1.1 | 0.354 | ||
| 分枝数No. of branches | 25.2 | <0.001 | 8.0 | <0.001 | 8.5 | <0.001 | ||
| 根长Total root length | 21.3 | <0.001 | 9.9 | <0.001 | 3.1 | 0.004 | ||
| 根直径Diameter | 70.7 | <0.001 | 5.1 | 0.001 | 2.9 | 0.006 | ||

|
| 图 3 水分处理及密度配置对牛鞭草形态及生长的影响(平均值±标准误, n=3) Fig. 3 Effects of water treatments and density configurations on morphology and growth of Hemarthria compressa (mean±SE, n=3) 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) |
密度配置对狗牙根株高、基茎、分枝数、根直径均有显著的影响(P < 0.05), 但对根长却未产生显著影响(表 2)。密度配置极显著的影响牛鞭草各形态指标(P < 0.01)(表 3)。牛鞭草在CK处理组以H2C10密度配置下株高最大, 单植(H12C0)条件下株高最小, 而浅淹与全淹条件下却以单植(H12C0)条件下的株高最大(图 3)。
水分与密度的交互作用对狗牙根的株高、基茎、分枝数、根长均有显著影响(P < 0.05), 但对根直径却没有显著影响(表 2)。对牛鞭草的基茎没有显著影响, 但对株高、分枝数、根长、根直径均有极显著影响(P < 0.01)(表 3)。
2.1.2 水淹胁迫和密度配置对狗牙根与牛鞭草生物量的影响水分处理极显著影响狗牙根总生物量、地下生物量、地上生物量和根冠比(P < 0.01)(表 4)。随着水淹深度的加深, 各部分平均生物量呈下降趋势, 根冠比呈上升趋势(图 4)。水分处理显著影响牛鞭草的各生物量及根冠比(P < 0.05)(表 5)。随着水淹深度的加深, 各部分平均生物量呈下降趋势, 而与之相反, 根冠比则随着水淹深度的增加呈现增加的趋势(图 5)
| 指标Index | 水分处理Water treatment | 密度配置Density configuration | 交互作用Interaction | |||||
| F | P | F | P | F | P | |||
| 总生物量Total biomass | 22.6 | <0.001 | 3.7 | 0.008 | 3.0 | 0.008 | ||
| 地上生物量Overground biomass | 11.9 | <0.001 | 1.1 | 0.115 | 1.0 | 0.475 | ||
| 地下生物量Root biomass | 37.2 | <0.001 | 3.4 | 0.008 | 13.9 | <0.001 | ||
| 根冠比Root/shoot | 6.5 | 0.002 | 0.4 | 0.317 | 2.7 | 0.006 | ||

|
| 图 4 水分处理及密度配置对狗牙根生物量的影响(平均值±标准误, n=3) Fig. 4 Effects of water treatments and density configurations on biomass of Cynodon dactylon (mean±SE, n=3) 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) |
| 指标Index | 水分处理Water treatment | 密度配置Density configuration | 交互作用Interaction | |||||
| F | P | F | P | F | P | |||
| 总生物量Total biomass | 126.7 | <0.001 | 12.5 | <0.001 | 8.2 | <0.001 | ||
| 地上生物量Overground biomass | 106.8 | <0.001 | 13.6 | <0.001 | 6.1 | <0.001 | ||
| 地下生物量Root biomass | 56.1 | <0.001 | 5.9 | <0.001 | 6.7 | <0.001 | ||
| 根冠比Root/shoot | 4.8 | 0.011 | 0.6 | 0.714 | 1.5 | 0.148 | ||

|
| 图 5 水分处理及密度配置对牛鞭草生物量的影响(平均值±标准误, n=3) Fig. 5 Effects of water treatments and density configurations on biomass of Hemarthria compressa (mean±SE, n=3) 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) |
密度配置极显著影响狗牙根的总生物量和地下生物量(P < 0.01)。在CK处理组, H0C12比例下的总生物量最大; 在浅淹组以H4C8比例下的总生物量最大; 在全淹组, H8C4比例下的总生物量最大(图 4)。在CK处理组, 牛鞭草各生物量在H2C10比例下最大, 而浅淹与全淹处理组的各生物量随着牛鞭草所占比例的增大而增大(H12C0比例除外)(图 5)。
水分与密度的交互作用极显著影响狗牙根的总生物量、地下生物量和根冠比(P < 0.01), 但对地上生物量却没有显著影响(表 4)。水分与密度的交互作用对牛鞭草各生物量有极显著影响(P < 0.01), 但对根冠比却无显著影响(表 5)。
2.2 水淹胁迫和密度配置对两草种种间竞争的影响 2.2.1 相对总生物量(RYT)不同水分处理及不同密度配置条件下两草种的RYT值均大于1。CK组H2C10比例下的RYT显著高于H10C2比例下的RYT(P < 0.05)。浅淹组, H2C10比例的RYT显著低于H8C4比例的RYT(P < 0.05)。在全淹组, 不同密度配置下的RYT无显著差异(表 6)。
| 处理Treatment | H2C10 | H4C8 | H6C6 | H8C4 | H10C2 |
| CK | 2.54±0.70a | 1.46±0.22ab | 1.61±0.16ab | 1.62±0.09ab | 1.29±0.15b |
| SF | 1.04±0.20b | 1.45±0.30ab | 1.80±0.23ab | 2.02±0.40a | 1.46±0.21ab |
| TF | 1.79±0.43a | 1.79±0.30a | 2.08±0.37a | 2.34±0.45a | 2.29±0.23a |
| CK:对照, Control; SF:浅淹, Shallow flooding; TF:深淹, Total flooding; 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) | |||||
在CK处理组, 狗牙根的相对竞争强度(RCIc)均大于0, H10C2密度配置下的RCIc最大。在浅淹组, H10C2比例下的RCIc最大且大于0, 其余比例下的RCIc值均小于0。在全淹组, H10C2比例下的RCIc值最大且大于0, 其余比例下的RCIc均小于0, H8C4比例下最小。
在CK组, 牛鞭草的相对竞争强度RCIh在H2C10比例下小于0, 在其他比例下均大于0, 且在H10C2比例下最大。浅淹组, RCIh均大于0, 且在H2C10比例下最大, 基本随着牛鞭草所占比例的增大而减小, H8C4比例下最小。全淹组, RCIh在H10C2比例下最小, 且小于0(表 7)。
| 物种Species | 处理Treatment | H2C10 | H4C8 | H6C6 | H8C4 | H10C2 |
| 狗牙根 | CK | 0.41±0.11a | 0.31±0.07a | 0.29±0.22a | 0.26±0.15a | 0.47±0.16a |
| Cynodon dactylon | SF | -0.13±0.20a | -0.17±0.26a | -0.28±0.24a | -0.22±0.16a | 0.18±0.13a |
| TF | -0.35±0.23a | -0.46±0.30a | -0.30±0.22a | -0.60±0.40a | 0.05±0.17a | |
| 牛鞭草 | CK | -0.95±0.65a | 0.23±0.29a | 0.09±0.14ab | 0.12±0.23ab | 0.32±0.85a |
| Hemarthria compressa | SF | 0.83±0.01a | 0.72±0.08a | 0.48±0.23ab | 0.01±0.29b | 0.16±0.21ab |
| TF | 0.55±0.29a | 0.67±0.04a | 0.22±0.16ab | 0.26±0.09a | -0.33±0.24b | |
| 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P<0.05) | ||||||
在CK组, 密度配置H4C8、H6C6处理下的牛鞭草A值大于0(狗牙根的A值为牛鞭草A值的相反数, 故不再单独分析), 其值分别为0.58和0.18, 其余比例下的值均为负值。在浅淹组, 密度配置H2C10处理下牛鞭草A值为0.70, 其余比例下的值均为负值。在全淹组, 密度配置H2C10处理下的A值为1.97, 其余比例下的值均为负值。且在不同水分处理条件下, A值随牛鞭草所占的比例的增加而降低(表 8)。
| 物种Species | 处理Treatment | H2C10 | H4C8 | H6C6 | H8C4 | H10C2 |
| 牛鞭草 | CK | -0.06±0.57ab | 0.58±0.10a | 0.18±0.32ab | -0.74±0.25a | -0.23±0.39ab |
| Hemarthria compressa | SF | 0.70±0.08a | -0.03±0.16ab | -0.76±0.41bc | -1.55±0.26c | -3.89±0.66d |
| TF | 1.97±1.40a | -0.07±0.18a | -0.52±0.07ab | -2.85±0.78b | -5.88±0.66c | |
| 不同小写字母分别表示同一水分处理组内不同密度配置之间有显著差异(P < 0.05) | ||||||
研究表明, 当水分等环境因子发生改变时, 植物的形态特征、生物量积累与根冠比[20]等均有不同程度的响应, 且植物会通过生物量的分配策略来适应环境的变化[21]。水淹环境中, 缺氧是影响植物生长的主要因素之一。长时间的供氧不足将导致植物由有氧呼吸逐渐向无氧呼吸转变, 在此过程中植物体内ATP的合成以及光合产物的同化也将大量减少, 植物生长因此受到明显的影响[22]。浅淹条件下耐水淹的植物会通过促进茎的伸长, 加快植物露出水面, 使其露出水面部分与空气接触, 获取足够的氧气以维持较强的光合作用, 从而缓解水淹对植物的损伤[23]。本研究中, 在相同比例条件下, 狗牙根与牛鞭草(除了H8C4、H10C2、H12C0)在浅淹处理组的株高大于对照组, 以此使植株与水面的空气尽快接触从而逃离水淹条件下的缺氧环境。可见, 在浅淹条件下, 牛鞭草和狗牙根通过维持或者促进茎偏上性伸长从而增加对水淹的耐受性。但对于全淹组来说, 牛鞭草和狗牙根却表现出不同的响应方式。牛鞭草在全淹环境下生长“停滞”, 植物株高与水淹前(表 1)相比无显著变化, 狗牙根却能够在水下持续伸长生长。植物在水下不同的生长表现反映了植物对完全水淹不同的耐受机制[24]。在全淹环境中, 牛鞭草生长“停滞”可以保存更多的营养物质以提供更长时间的能量供应, 维持植株存活; 狗牙根伸长的枝条是植物对全淹环境积极的适应性响应, 对于植物缓解水下缺氧及低光照环境, 甚至是“逃离”水淹逆境都是有利的[25-26]。因此可以利用狗牙根和牛鞭草对水淹的不同响应进行三峡库区消落带不同海拔位草本植被的恢复。
水淹环境存在光线较弱、氧气不足等不利条件, 这都将严重影响植物的生长及形态变化。有研究表明湿地植物往往通过控制根长、增加根的直径、扩大根冠比、产生不定根等方式克服氧气供应不足, 维持植物正常的生长发育[27]。本试验全淹处理下, 两草种的根长与CK组相比均有显著降低, 这是两草种在深水淹条件下, 通过减少根长和根系分布的深度以减少厌氧呼吸产生的有害物质对根部的伤害[28]。同时两草种的根直径都有显著增加, 这是由于深水淹时, 植物能够增加根的直径, 提高气体在根部的运输能力[29]以适应深水淹条件下的缺氧环境。Pace等[30]对棉花的幼苗研究表明水分胁迫对其地上部分的影响作用明显高于地下部分, 因而这种不同步的影响作用导致了根冠比的增加。本试验结果与之相似。在浅淹处理组, 狗牙根和牛鞭草虽然表现出茎伸长的形态响应, 但同时水下的叶片也枯萎脱落, 使得地上生物量增加的比重不是很大。相反, 在此处理下, 植株会产生不定根来减缓水下缺氧环境对植株造成的伤害, 从而使根生物量比重增大; 在全淹处理组, 狗牙根和牛鞭草由于无法获取充足的光资源进行光合作用, 从而植株变的矮小且地上生物量也没有增加, 而植株在全淹处理前积累的营养物质通过物质运输转移到根部存储以维持植株的存活从而使得根部的生物量增加[31], 这是二者根冠比增加的原因。
有研究指出, 全淹条件下, 植物能通过减缓植物生长、损失部分生物量以增加对水淹的耐受性[32]。在本试验中, 无论是单植还是混植条件下, 牛鞭草与狗牙根的生物量在各比例条件下均随着水分胁迫的加剧而显著降低, 但两者之间又表现出不尽相同的变化特征。对于狗牙根而言, 其总生物量在水淹处理下低于CK组, 且其地下部分生物量下降显著, 这可能是由于在水淹条件下其茎的伸长消耗了大量营养储备, 造成营养储备器官(地下部分)生物量显著下降[32]; 对于牛鞭草而言, 其各部分生物量随着水淹的深度加深而呈下降趋势, 但在全淹处理下其地上部分生物量下降显著, 这可能是由于在全淹组, 牛鞭草生长“停滞”, 其地上部分的营养物质运输到地下储存以维持其存活的原因。
3.2 不同密度配置对狗牙根及牛鞭草的影响种植密度主要通过影响植物的光资源和养分资源来影响植物的生长。在CK组, 密度配置H8C4比例下狗牙根的株高大于单植(H0C12)下的株高, 这可能是由于在对地上资源光照的竞争中, 在冠层中占优势的牛鞭草减少了狗牙根进行光合作用所需要的阳光辐射, 狗牙根为了获取更多的光照不得不采取增加高度的策略来满足自身生长的需求, 从而促进了高度增加[33]。但狗牙根在比例H8C4下的总生物量却低于狗牙根单植(H0C12)下的总生物量, 其原因可能是由于在H8C4比例下, 牛鞭草所占比例较大, 且其叶片宽大, 郁闭度好, 并且生长初期保持直立生长, 冠层距离地面较高, 这导致从根部到冠层很大空间内都是郁闭环境[34], 使得狗牙根很难突破牛鞭草的覆盖, 从而导致地上生物量减少; 另一方面, 经过对两个物种根部的分析发现牛鞭草的根长大于狗牙根的根长, 这就使得二者在对地下部分营养物质竞争的过程中, 牛鞭草更能够深入土壤来获取营养物质, 从而使狗牙根在地下竞争过程中处于劣势而导致其地下生物量减小, 所以在H8C4比例下, 狗牙根的地下生物量小于狗牙根单植(H0C12)下的地下生物量。在水淹(SF组、TF组)处理下, 狗牙根在不同比例处理下的株高及总生物量变化没有规律, 但值得注意的是在TF组, 其总生物量在H10C2下最小, 而牛鞭草的总生物量在此比例下最大, 这可能时由于牛鞭草的种内竞争限制了狗牙根的生长。
通过研究密度配置对牛鞭草的影响发现, 在CK组, 牛鞭草的株高和总生物量都是在牛鞭草所占比例最低(H2C10)的条件下达到最大值, 且显著高于其他比例; 而其株高在牛鞭草所占最高比例即单种(H12C0)下最低, 且其单种条件下的总生物量也显著低于H2C10比例下的生物量, 这是因为牛鞭草种内竞争比较激烈。实验结果可以看出牛鞭草的株高和生物量对比例响应明显, 混种比例H2C10使牛鞭草受益, 这可能是因为牛鞭草的株型较大, 盆栽环境对其来说显得太过拥挤, 因而在低比例下株高和生物量明显增大[34]。在水淹处理下, 牛鞭草的最大株高和最大总生物量基本出现在其所占比例较大的处理下, 出现这些现象的原因可能是由于牛鞭草属于直立生长的禾本科草种, 在其密度较大时, 资源生态位重叠较大, 植株对光资源的竞争激烈, 促使植株增加高度来获得更多资源空间[35]。
3.3 不同水分处理及密度配置对狗牙根及牛鞭草种间竞争的影响相对总生物量可以用来精确的评价混植条件下的最大生物学效率[36]。在本试验中, 狗牙根与牛鞭草的混植体系相对总生物量(RYT)均大于1, 说明在狗牙根与牛鞭草混植条件下, 两物种均占有不同的生态位, 在整个试验过程中表现出一定的共生关系。这是因为牛鞭草属于直立生长的禾本科草种, 而狗牙根在生长过程中整个植株大部分处于匍匐状态, 依赖自身生长特性横向扩展, 这使得二者的空间生态位分离[35], 种间竞争较弱, 两草种表现为共生关系。相对总生物量(RYT)值只能说明植物种间在资源利用上不同, 而相对竞争强度(RCI)则可以用来衡量混植植物种间竞争强度和种内竞争强度的大小[37]。
植物的大小特征是预测植物相对竞争力的最佳指标, 生物量是其中最重要的参数, 它可以反映植物相对竞争力的64%左右[38]。在CK处理组不同的密度配置下, 狗牙根混种时的单株生物量与单种时相比, 按照比例H2C10、H4C8、H6C6、H8C4、H10C2的变化分别减少45.8%、33.9%、35.8%、26.1%、52.1%, 同样, 牛鞭草的单株生物量也随种植比例的变化而变化, 不同的是与单种时相比, 牛鞭草的单株生物量在H2C10比例下增加了26.8%, 在其他比例下, 按照H4C8、H6C6、H8C4、H10C2的变化分别减少33.5%、15.5%、19.6%、23.3%, 以上结果表明, 狗牙根和牛鞭草的生长均具有明显的密度制约效应, 但狗牙根的反应更为强烈。秦先燕等[39]在2010年的研究中表明在环境适宜时, 植物能获得较多的资源, 此时种间关系以竞争关系为主。如在非淹水条件下, 海滨碱茅(Inellia maritime)和大米草(Spartina anglica)间竞争强烈。在同一种植比例下, 狗牙根的单株生物量均低于相同比例下牛鞭草的单株生物量, 狗牙根的生长受到牛鞭草的抑制。同时结合狗牙根与牛鞭草在CK处理组各混植体系中的相对竞争强度RCI, 发现狗牙根的RCI值均大于0, 即说明狗牙根的种间竞争大于种内竞争, 牛鞭草在该处理下对其产量负影响较大。这是因为本试验在花盆中进行的, 因此提供给植物生长所必须的养分是一定的, 在营养资源相对充足的条件下, 生长较快的植物能够迅速占据生态位并较早利用资源, 能使其更具有竞争优势[40]。试验发现牛鞭草与狗牙根相比, 具有发达的根系, 宽大的叶片以及粗壮的基茎, 使其在生长过程中更强地捕获周围的土壤养分、阳光以及水分, 使其更具有竞争优势。
有研究表明湿地植物竞争强度与水位相关。在水位适宜的条件下, 植物间竞争激烈, 但随着淹水深度增加, 其竞争强度有明显减弱, 甚至产生对相邻物种的促进作用[41]。如在0 cm水位处, 三江平原地区狭叶甜茅(Glyceria spiculosa)的存在会抑制毛薹草(Carex lasiocarpa)和小叶章的生长; 但当水位增加到40 cm时, 反而促进了毛薹草和小叶章的生物量积累, 植物间关系表现为促进作用[42]。在浅淹组, 狗牙根的相对竞争强度除了在比例H10C2下大于0外, 在其他比例下均小于0, 即种间竞争小于种内竞争, 再结合其在浅淹组的总生物量来看, 混植体系中狗牙根的生物量大于其单植条件下的生物量, 说明牛鞭草的存在促进了其产量的增加。在全淹组, 狗牙根的相对竞争强度在大多比例下的值小于0, 说明牛鞭草对其的影响作用较小, 而牛鞭草在该水分处理下的相对竞争强度在大多比例下大于0, 说明狗牙根对其的影响作用较大, 再结合该水分条件下的竞争攻击系数(A), 牛鞭草的竞争攻击系数在大多比例下小于0, 说明在全淹组, 狗牙根更具有竞争优势。这可能与狗牙根和牛鞭草各自的耐水淹性质有关, 有研究指出水淹时耐淹植物野稗(Echinochloa crusgali)、芦苇(Phragmites australis)、香蒲(Typha orientalis)比不耐淹植物碱茅(Puccinellia hinampoensis)具有竞争优势[43], 也有研究表明湿地植物对环境胁迫的耐受性与竞争能力在特定环境中存在权衡关系, 如在低盐环境中, 强耐盐性的物种竞争力反而较差[44]。当环境条件适宜时, 植物之间竞争占主导, 当环境胁迫较严重时, 植物之间的耐受性占主导作用, 植物倾向于将大部分资源分配到相应的性状或器官, 或通过形态和生理的变化适应环境, 但关于狗牙根与牛鞭草的耐水淹性与竞争能力之间的权衡关系还需进一步研究了解。
4 结论本研究结果表明, 狗牙根和牛鞭草面对水淹胁迫和不同密度配置均表现出一定的敏感性和可塑性。在各处理下植株的存活率达100%, 且植株能通过生物量的分配、分枝数与根长的减少和根直径的增加等对水淹胁迫做出积极的响应。不同密度配置之间的植株也表现出了一定的差异。在CK组, 随着牛鞭草所占比例的增大, 狗牙根的单株生物量有减小的趋势, 该处理下, 狗牙根与牛鞭草表现出的是竞争关系; 而在水淹处理下, 二者的种间竞争作用减小, 并表现出一定的促进作用。狗牙根与牛鞭草混植在一定条件下优于单植。从混植体系相对总生物量的角度考虑, 本试验建议, 常规供水处理下建植牛鞭草和狗牙根人工草地的最佳配置比例为H2C10, 而水淹处理下最佳配置比例为H8C4。
| [1] | 吴炳方, 刘远新, 臧小平, 程温鸣, 张静, 周维. 三峡工程建设期库区生态环境保护措施及效果评价. 长江流域资源与环境, 2011, 20(3): 276–282. |
| [2] | Yang F, Liu W W, Wang J, Liao L, Wang Y. Riparian vegetation's responses to the new hydrological regimes from the Three Gorges Project:Clues to revegetation in reservoir water-level-fluctuation zone. Acta Ecologica Sinica, 2012, 32(2): 89–98. DOI:10.1016/j.chnaes.2012.02.004 |
| [3] | Holmes P M, Esler K J, Richardson D M, Witkowski E T F. Guidelines for improved management of riparian zones invaded by alien plants in south Africa. South African Journal of Botany, 2008, 74(3): 538–552. DOI:10.1016/j.sajb.2008.01.182 |
| [4] | 陶敏, 鲍大川, 江明喜. 三峡库区9种植物种子萌发特性及其在植被恢复中的意义. 生态学报, 2011, 31(4): 906–913. |
| [5] | 洪明. 三峡库区消落带3种草本植物对水陆生境变化的响应[D]. 北京: 中国林业科学研究院, 2011. |
| [6] | 姜汉侨, 段昌群, 杨树华, 王崇云, 苏文华. 植物生态学. 北京: 高等教育出版社, 2004. |
| [7] | 樊江文, 钟华平, 杜占池, 韩彬, 梁飚. 草地植物竞争的研究. 草业学报, 2004, 13(3): 1–8. |
| [8] | Donald C M, Hamblin J. The Convergent Evolution of Annual Seed Crops in Agriculture. Advances in Agronomy, 1983, 36: 97–143. DOI:10.1016/S0065-2113(08)60353-3 |
| [9] | 谢开云, 赵云, 李向林, 何峰, 万里强, 王丹, 韩冬梅. 豆-禾混播草地种间关系研究进展. 草业学报, 2013, 22(3): 284–296. DOI:10.11686/cyxb20130337 |
| [10] | 谭淑端, 朱明勇, 党海山, 王勇, 张全发. 三峡库区狗牙根对深淹胁迫的生理响应. 生态学报, 2009, 29(7): 3685–3691. |
| [11] | 李兆佳, 熊高明, 邓龙强, 谢宗强, 樊大勇. 狗牙根与牛鞭草在三峡库区消落带水淹结束后的抗氧化酶活力. 生态学报, 2013, 33(11): 3362–3369. |
| [12] | Yang C H, Zhang X Q, Li X L, Du Y, Wu Y C. Hemarthria germplasm resources and breeding. Acta Prataculturae Sinica, 2004, 13(2): 7–12. |
| [13] | 曾成城, 王振夏, 陈锦平, 顾艳文, 贾中民, 魏虹. 不同水分处理对狗牙根种内相互作用的影响. 生态学报, 2016, 36(3): 696–704. |
| [14] | 陈锦平, 王振夏, 曾成城, 李帅, 魏虹. 不同水分和植株密度处理对牛鞭草生长及形态的影响. 草业学报, 2015, 24(1): 39–46. DOI:10.11686/cyxb20150106 |
| [15] | 黄玺, 李春杰, 南志标. 紫花苜蓿与醉马草的竞争效应. 草业学报, 2012, 21(1): 59–65. DOI:10.11686/cyxb20120108 |
| [16] | 郭泉水, 洪明, 康义, 裴顺祥, 程瑞梅. 消落带适生植物研究进展. 世界林业研究, 2010, 23(4): 14–20. |
| [17] | Weigelt A, Jolliffe P. Indices of plant competition. Journal of Ecology, 2003, 91(5): 707–720. DOI:10.1046/j.1365-2745.2003.00805.x |
| [18] | 蒋智林, 刘万学, 万方浩, 李正跃. 植物竞争能力测度方法及其应用评价. 生态学杂志, 2008, 27(6): 985–992. |
| [19] | Jolliffe P A. The replacement series. Journal of Ecology, 2000, 88(3): 371–385. DOI:10.1046/j.1365-2745.2000.00470.x |
| [20] | Ares A, Burner D M, Brauer D K. Soil phosphorus and water effects on growth, nutrient and carbohydrate concentrations, δ13C, and nodulation of mimosa (Albizia julibrissin Durz.) on a highly weathered soil. Agroforestry Systems, 2009, 76(2): 317–325. DOI:10.1007/s10457-009-9213-2 |
| [21] | Mcconnaughay K D M, Coleman J S. Biomass allocation in plants:ontogeny or optimality? A test along three resource gradients. Ecology, 1999, 80(8): 2581–2593. DOI:10.1890/0012-9658(1999)080[2581:BAIPOO]2.0.CO;2 |
| [22] | Mukassabi T A, Polwart A, Coleshaw T, Thomas P A. How long can young Scots pine seedlings survive waterlogging?. Trees, 2012, 26(5): 1641–1649. DOI:10.1007/s00468-012-0740-5 |
| [23] | Voesenek L A C J, Colmer T D, Pierik R, Millenaar F F, Peeters A J M. How plants cope with complete submergence. New Phytologist, 2006, 170(2): 213–226. DOI:10.1111/nph.2006.170.issue-2 |
| [24] | Crawford R M M. Whole plant adaptations to fluctuating water tables. Folia Geobotanica, 1996, 31(1): 7–24. DOI:10.1007/BF02803990 |
| [25] | Canadell J, López-Soria L. Lignotuber reserves support regrowth following clipping of two Mediterranean shrubs. Functional Ecology, 1998, 12(1): 31–38. DOI:10.1046/j.1365-2435.1998.00154.x |
| [26] | Ishizawa K, Murakami S, Kawakami Y, Kuramochi H. Growth and energy status of arrowhead tubers, pondweed turions and rice seedlings under anoxic conditions. Plant Cell & Environment, 1999, 22(5): 505–514. |
| [27] | 杨娇, 厉恩华, 蔡晓斌, 王智, 王学雷. 湿地植物对水位变化的响应研究进展. 湿地科学, 2014, 12(6): 807–813. |
| [28] | Bouma T J, Nielsen K L, Van Hal J, Koutstaal B. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Functional Ecology, 2001, 15(3): 360–369. DOI:10.1046/j.1365-2435.2001.00523.x |
| [29] | Visser E J W, Blom C W P M, Voesenek L A C J. Flooding-induced adventitious rooting in Rumex:morphology and development in an ecological perspective. Acta Botanica Neerlandica, 1996, 45(1): 17–28. DOI:10.1111/plb.1996.45.issue-1 |
| [30] | Pace P F, Cralle H T, Cothren J T, Senseman S A. Photosynthate and dry matter partitioning in short-and long-season cotton cultivars. Crop Science, 1999, 39(4): 1065–1069. DOI:10.2135/cropsci1999.0011183X003900040018x |
| [31] | Miller R C, Zedler J B. Responses of native and invasive wetland plants to hydroperiod and water depth. Plant Ecology, 2003, 167(1): 57–69. DOI:10.1023/A:1023918619073 |
| [32] | 王海锋, 曾波, 乔普, 李娅, 罗芳丽, 叶小齐. 长期水淹条件下香根草(Vetiveria zizanioides)、菖蒲(Acorus calamus)和空心莲子草(Alternanthera philoxeroides)的存活及生长响应. 生态学报, 2008, 28(6): 2571–2580. |
| [33] | 杨春华. 扁穗牛鞭草与混生种互作的生理生态机理研究[D]. 雅安: 四川农业大学, 2004. |
| [34] | 刘冰. 2种替代植物与紫茎泽兰竞争效应及竞争相关指标适用性探索[D]. 秦安: 山东农业大学, 2011. |
| [35] | 张静, 赵成章, 盛亚萍, 张军霞, 史丽丽. 高寒山区混播草地燕麦和毛苕子种间竞争对密度的响应. 生态学杂志, 2012, 31(7): 1605–1611. |
| [36] | Zand E, Beckie H J. Competitive ability of hybrid and open-pollinated canola (Brassica napus) with wild oat (Avena fatua). Canadian Journal of Plant Science, 2002, 82(2): 473–480. DOI:10.4141/P01-149 |
| [37] | 王晋萍, 董丽佳, 桑卫国. 不同氮素水平下入侵种豚草与本地种黄花蒿、蒙古蒿的竞争关系. 生物多样性, 2012, 20(1): 3–11. |
| [38] | Keddy P, Nielsen K, Weiher E, Lawson R. Relative competitive performance of 63 species of terrestrial herbaceous plants. Journal of Vegetation Science, 2002, 13(1): 5–16. DOI:10.1111/j.1654-1103.2002.tb02018.x |
| [39] | 秦先燕, 谢永宏, 陈心胜. 湿地植物间竞争和促进互作的研究进展. 生态学杂志, 2010, 29(1): 117–123. |
| [40] | 盛亚萍, 赵成章, 高福元, 张军霞, 石福习, 屈勇. 高寒山区混播草地燕麦和毛苕子种间的竞争关系. 生态学杂志, 2011, 30(11): 2437–2441. |
| [41] | 章泉. 鄱阳湖湿地虉草的生态适应性研究[D]. 南昌: 南昌大学, 2012. |
| [42] | 罗文泊. 三江平原沼泽植物带状分布格局的成因[D]. 长春: 中国科学院东北地理与农业生态研究所, 2008. |
| [43] | 王正文, 邢福, 祝廷成, 李宪长. 松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应. 植物生态学报, 2002, 26(6): 708–716. |
| [44] | Egan T P, Ungar I A. Competition between Salicornia europaea and Atriplex prostrata (Chenopodiaceae) along an experimental salinity gradient. Wetlands Ecology and Management, 2001, 9(6): 457–461. DOI:10.1023/A:1012276510818 |