 2018, Vol. 38
2018, Vol. 38文章信息
- 邵蕊, 赵苗苗, 赵芬, 沈瑞昌, 刘丽香, 张丽云, 徐明.
- SHAO Rui, ZHAO Miaomiao, ZHAO Fen, SHEN Ruichang, LIU Lixiang, ZHANG Liyun, XU Ming.
- 施肥对油茶园土壤呼吸和异养呼吸及其温度敏感性的影响
- Effects of fertilization on soil respiration, heterotrophic respiration, and temperature sensitivity in an oil tea plantation
- 生态学报. 2018, 38(7): 2315-2322
- Acta Ecologica Sinica. 2018, 38(7): 2315-2322
- http://dx.doi.org/10.5846/stxb201703270531
-
文章历史
- 收稿日期: 2017-03-27
- 网络出版日期: 2018-01-04

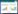




2. 中国科学院大学, 北京 100049;
3. 南昌大学生命科学研究院流域生态学研究所, 南昌 330031
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Center for Watershed Ecology, Institute of Life Science, Nanchang University, Nanchang 330031, China
人类活动引起的温室气体排放的增加是全球变暖的主要诱因, 自工业化以来, 大气中二氧化碳、甲烷和氧化亚氮浓度显著增加[1], 其中, 以CO2为主。土壤是陆地生态系统最大的碳库[2], 其碳循环过程对调节大气CO2浓度和地球系统的气候动态具有十分重要的作用[3]。其中, 土壤呼吸(Rs)是大气和陆地生态系统间仅次于光合作用的碳通量渠道。土壤呼吸一方面直接影响大气中CO2浓度, 另一方面通过对气候变暖的响应进一步影响陆地生态系统碳循环。土壤呼吸对温度变化响应的剧烈程度, 即温度敏感性, 是陆地生态系统对气候变暖响应的重要参数, 可用Q10来表示, 即温度每升高10℃, 温室气体排放速率变化的倍数[4]。
施肥对大气和陆地生态系统碳循环具有重要影响, 一方面, 施肥通过促进作物生长增加生态系统碳固定影响土壤碳库; 另一方面, 通过调节根系分泌物、土壤微生物群落组成与功能、无机氮含量等土壤理化性质, 影响土壤呼吸。但由于环境因子和实验方法等因素的差异, 施肥对土壤呼吸及其组成的影响尚没有统一结论[3, 5]。有些研究表明, 施肥显著促进土壤呼吸[6-7], 其他一些研究则显著降低土壤呼吸[8-9]或者对土壤呼吸没有显著影响[10]。除了直接影响土壤呼吸, 施肥还可以通过影响土壤碳库来影响土壤呼吸温度敏感性。根据Arrhenius方程, 难分解有机质通常具有更高的温度敏感性, 而根据Michaeli-Menten酶促反应动力学方程, 底物有效性能够通过影响Vmax和Km的温度敏感性, 影响有机质分解的温度敏感性[4, 11], 并最终影响陆地生态系统对气候变化的响应。研究施肥对土壤呼吸温度敏感性的影响对预测未来气候变化背景下陆地生态系统碳循环过程具有重要意义。
油茶(Camellia oleifera)是中国南方重要的木本食用油料树种, 但是不同于森林等自然生态系统, 油茶栽培对土壤肥力要求很高, 需肥量大, 合理的施肥是促进油茶生长、提高油茶产量和品质, 以及防范土壤肥力衰退的重要措施[12-13]。随着产业的发展和土地集约利用, 油茶园种植面积逐渐增大, 研究施肥对油茶园土壤呼吸及其温度敏感性的影响, 对于估算中国南方典型种植园林温室气体排放及其对气候变化的响应具有重要意义。本研究采用静态箱-气相色谱法, 通过多年观测, 分析探讨施肥对油茶园土壤土壤呼吸和异养呼吸及其温度敏感性的影响。
1 研究区概况本研究野外试验在中国科学院千烟洲红壤丘陵综合开发试验站(以下简称千烟洲站)展开。千烟洲站位于江西省泰和县灌溪镇, 115°04′13′′E, 26°44′48′′N。该区域属于典型的亚热带大陆性季风气候, 年平均气温为17.9℃; 多年平均降水量约为1490mm, 主要集中在3—6月, 占全年降水量的51%。土壤类型为红壤, 成土母质主要包括红色砂岩和砂砾岩的风化产物, 粘粒含量19%(<0.002mm), 粉砂含量25%(0.002—0.02mm), 砂粒含量56%(0.02—2mm)。土壤pH值为4.97, 土壤容重是1.29g/cm3。
2 研究方法 2.1 试验设计设置施肥(OF)与对照(CK)两个处理, 每个处理3个重复。每个样方为14m×10m的矩形, 植株间距为2.8m×2m。样方内随机设置2个静态箱进行观测。为了研究施肥对油茶园土壤异养呼吸及其温度敏感性的影响, 2013年3月初, 采用壕沟法[14-15]在每个样方内设置1m×1m断根子样方。根据前期调查油茶根系分布深度, 挖沟槽深70cm, 宽10cm。沟槽挖好后将PVC板贴断根区内壁放入槽内, 回填底层土壤压实, 依次固定四周PVC板, 并保证PVC板连接处闭合, 然后再回填其余土壤并压实。然后每个子样方中设置1个静态箱, 其中对照和施肥处理的子样方分别标记为CK-T和OF-T。各处理土壤CO2通量观测从2013年5月1日持续至2016年4月30日。CK采样期内, 定期清理断根区内新生草芽, 选择在草芽较小时用剪刀齐地剪掉, 尽量减少对表层土壤的干扰。试验所用化肥为复合肥(N:P2O5:K2O 15%:15%:15%), 施肥量为360kg N hm-2 a-1, 在每年的4月中旬和8月中旬分两次施入。施肥前将肥料充分溶解于水中, 然后将肥料溶液均匀喷洒于各样方中。
2.2 土壤CO2通量测定与计算本研究采用静态箱-气相色谱法进行土壤呼吸测定。静态箱采用底面直径为49cm, 高为39cm的不锈钢暗箱。在8:00—11:00之间, 通过半自动人工采样系统, 在扣箱0、10、20、30、40min后直接将气体采集到12mL已抽真空的顶空进样瓶。生长季通常为每4—5d采集1次气样, 在施肥或强降雨后加密采样, 每天1次持续1周。非生长季每10—15d采集1次气样。
利用气相色谱仪(GC System, 7890A, Agilent Technologies)测定气样CO2浓度, 利用公式1计算通量。

|
(1) |
式中, F为CO2净交换通量(mg C m-2 h-1), ρ为标准状态下CO2气体密度(0.536kg/C m3), V是采样箱的有效体积(m3), A为采样箱所覆盖的土壤面积(m2), T和T0为观测时箱内温度和标准状态温度(℃), P和P0为观测时的大气压和标准大气压(kPa), dCt/dt为箱内气体浓度随时间变化的斜率(μL/L)。
2.3 土壤理化性质及微气象因子观测静态箱采取地表温室气体的同时, 手工测定箱内空气温度。土壤温度通过土壤小气候自动观测系统获取, 温度传感器埋在土壤表层下5cm深处。在气体采样结束后的24h内, 采集土壤表层(0—10cm)土壤样品。为减少土样采集对样地干扰, 研究期第1年生长季每12—15d采集1次土壤样品, 非生长季每月底取样; 第2年和第3年, 平均每月采集1次土壤样品。将新鲜土样过2mm筛后取80g左右放置冷柜保存。土壤无机氮浓度按照土水比1:5(W/V), 利用1mol/L KCl溶液提取后过滤, 滤液上流动分析仪(AA3 HR AutoAnalyzer, SEAL, Germany)进行分析测定。土壤DOC按照土水比1:5(W/V)利用超纯水提取, 离心并取上清液抽滤后, 用流动分析仪测定(AA3 HR AutoAnalyzer, SEAL, Germany)。
2.4 数据处理与分析本研究采用Van′t Hoff指数模型(公式(2))来模拟土壤呼吸与温度的关系。
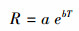
|
(2) |
式中, R为土壤呼吸速率, T为土壤温度, a指温度为0℃时的土壤呼吸速率, b为温度反应系数。
采用公式(3)来计算土壤CO2通量的温度敏感性Q10,
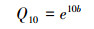
|
(3) |
式中, b为公式(2)计算所得温度反应系数。
所有数据经统计正态性检验, 不满足正态性分布的情况下, 采用自然对数转换将其转变为服从正态性分布后在进行统计分析。本研究采用单因素方差分析方法检验处理间土壤CO2通量, 年累积排放量和温度敏感性(Q10)差异的显著性。所有统计分析均在IBM SPSS 22.0(IBM, New York, United States)中进行。图形由SigmaPlot 10.0软件实现。
3 研究结果 3.1 气温、土壤温度和降水研究期间, 千烟洲试验站日均气温变化范围为-1.75—31.79℃, 年内气温呈单峰形态变化, 最高日均温出现在7月份, 最低日均温出现在1月份(图 1)。土壤温度表现出与气温一致的季节变化规律(图 1), 日均温变化范围为3.83—37.81℃。降水年内分配不均, 主要集中在3—6月, 8—10月降水较少, 易出现伏旱。

|
| 图 1 研究期间空气温度、土壤温度和降水量 Fig. 1 Seasonal dynamics of air temperature, soil temperature and precipitation during the study period |
不同处理下, CO2通量季节变化规律相似(图 2), 均与温度的季节变化基本一致。研究期间, OF和CK处理CO2通量变化范围分别为16.93—195.82mg C m-2 h-1和12.83—179.21mg C m-2 h-1, 平均值分别为(77.91±2.585)mg C m-2 h-1和(73.71±0.97)mg C m-2 h-1; OF-T和CK-T处理CO2通量变化范围分别为9.32—175.58mg C m-2 h-1和9.24—146.00mg C m-2 h-1, 平均值分别为(66.82±1.02)mg C m-2 h-1和(66.84±3.94)mg C m-2 h-1。方差分析结果表明, 研究期间OF和CK处理间CO2通量无显著差异(F=0.064, P=0.802), OF-T和CK-T处理间CO2通量也无显著差异(F=0.020, P=0.889), 即施肥对土壤呼吸和异养呼吸均无显著影响。

|
| 图 2 土壤呼吸和异养呼吸季节变化动态 Fig. 2 Seasonal dynamics of soil respiration and heterotrophic respiration during the study period OF:施肥, oil tea plots with fertilization; CK:对照, oil tea plots without fertilization; OF-T:断根施肥, trenching plots with fertilization; CK-T:断根, trenching plots without fertilization |
不同处理下, CO2通量均与土壤温度呈显著指数相关关系(P<0.01), 其中土壤温度约可解释OF和CK处理63.6%—66.5%的变异(图 3), 可解释OF-T和CK-T处理54.4%—64.2%的变异(图 4)。根据指数方程系数计算得不同处理下CO2通量的温度敏感性(CO2-Q10)。其中, OF和CK处理CO2-Q10变化范围分别为1.74—1.80和1.68—1.80;OF-T和CK-T处理CO2-Q10变化范围分别为1.94—1.97和1.73—1.85。各处理Q10依次为OF-T (1.96±0.01)>> CK-T(1.79±0.03)> OF(1.77±0.01)> CK(1.75±0.03)(图 5)。方差分析结果表明, OF和CK处理CO2-Q10无显著差异(F=0.257, P=0.639), 但OF-T处理CO2-Q10显著高于CK-T处理(F=21.739, P=0.010)(图 5)。

|
| 图 3 OF样方和CK样方, 土壤呼吸与土壤温度的指数相关关系 Fig. 3 The exponential relationships between soil respiration and soil temperature in the OF and CK plots |

|
| 图 4 OF-T样方和CK-T样方, 土壤异养呼吸与土壤温度的指数相关关系 Fig. 4 The exponential relationships between soil heterotrophic respiration and soil temperature in the OF-T and CK-T plots |

|
| 图 5 不同处理下土壤CO2通量温度敏感性 Fig. 5 Temperature sensitivity (Q10) of soil CO2 effluxes in different treatments OF:施肥, oil tea plots with fertilization; CK:对照, oil tea plots without fertilization; OF-T:断根施肥, trenching plots with fertilization; CK-T:断根, trenching plots without fertilization.不同字母表示处理间差异显著(P<0.05) |
结果显示, 施肥对土壤温度、湿度和土壤表层DOC含量均无显著影响, 但是显著增加了土壤表层NH4+-N和NO3--N含量(表 1)。不同处理下, 土壤表层NH4+-N和NO3--N含量均有OF-T > OF > CK-T > CK(表 1)。其中, OF-T样方土壤表层NH4+-N和NO3--N含量明显高于其他处理。这与土壤CO2通量Q10表现一致(图 5)。将不同处理Q10分别与土壤表层NH4+-N和NO3--N含量做简单线性相关分析, 结果表明, Q10与土壤表层NH4+-N和NO3--N含量表现出显著的正相关关系(图 6)。
| 处理 Treatment |
NH4+-N/(mg/kg) | NO3--N/(mg/kg) |
| OF | 28.67±1.55b | 12.89±1.71b |
| CK | 7.23±0.48d | 4.89±1.68c |
| OF-T | 98.17±3.12a | 24.34±2.66a |
| CK-T | 15.52±0.82c | 9.10±1.37bc |

|
| 图 6 土壤CO2通量温度敏感性与土壤表层NH4+-N和NO3--N含量的相关关系 Fig. 6 The relationships between temperature sensitivity (Q10) of soil CO2 effluxes (Q10) and soil NH4+-N and NO3--N content OF:施肥, oil tea plots with fertilization; CK:对照, oil tea plots without fertilization; OF-T:断根施肥, trenching plots with fertilization; CK-T:断根, trenching plots without fertilization.不同字母表示土壤表层同一物质含量处理间差异显著(P<0.05) |
本研究中, 油茶园土壤呼吸在该地区人工林土壤呼吸的变化范围48.67—84.02mg C m-2 h-1内[9-10, 16-17], 与Sheng等[18]对中国南方亚热带地区橘园和王超等[19]对福建栗园(66.96mg C m-2 h-1)的研究结果相接近(73.01mg C m-2 h-1), 但明显低于Liu等[20]对龙眼种植园(109.89mg C m-2 h-1)和王超等[19]对柑橘园(115.78mg C m-2 h-1)的观测结果, 高于Iqbal等[21]对桃园土壤呼吸的研究结果(22.64mg C m-2 h-1)。这种差异可能是由于区域气候[3], 不同园林细根生物量、植株生产力及地下碳分配、土壤碳库[20], 以及施肥等管理方式的不同造成的。此外, 研究方法不同也可产生一定的差异[5]。
本研究中, 施肥对土壤呼吸和异养呼吸均无显著影响, 但在一定程度上促进了Rs, 这与Deng等[22]对中国亚热带森林的研究结果一致, 可能是由于施肥促进了油茶植株生长, 使根系生物量增加, 因此促进了Rs。已有研究表明, 施肥对Rs的促进作用主要是由于施肥对根系自养呼吸的影响造成的[3, 22]。此外, 施肥对Rh表现出微弱的削减作用, 这与Zhou等[3]的萃取分析结果一致, 其研究表明氮肥可通过改变土壤微生物群落、降低微生物生物量、抑制土壤氧化酶活动等途径抑制土壤异养呼吸, 因此削弱了土壤异养呼吸。
4.2 施肥对油茶园土壤呼吸温度敏感性的影响一项萃取分析的研究结果表明, 全球各类陆地生态系统土壤呼吸的温度敏感性(Q10)均值为(2.6±1.2), 中位数为2.39, 其中沼泽湿地(3.3±0.3)和苔原(3.4±0.3)显著高于森林(2.5±0.2)和草地(2.5±0.2)[23]。本研究中, 油茶园土壤呼吸温度敏感性Q10变化范围为1.68—1.97, 低于全球森林土壤呼吸温度敏感性, 稍高于Sheng等[18]和王超等[19]对中国南方亚热带橘园的研究结果(Q10分别为1.66和1.58), 但是与中国亚热带森林(1.86)[22]、中国亚热带地区果园有机碳矿化Q10(1.81)[24]以及锥栗园土壤呼吸Q10(1.75)[21]相近的研究结果相近。此外, 油茶园土壤异养呼吸Q10与Winkler等[25]的研究结果一致。
本研究中, 施肥显著增加了油茶园土壤异养呼吸Q10, 对土壤呼吸Q10也表现出促进作用, 与Cusack等[12]和Deng等[22]的研究结果一致, 即施肥可以增加一些土壤碳库的温度敏感性。这可能是由于施肥后土壤中无机氮含量增加, 抑制了土壤氧化酶活性, 导致相对于对照处理, 施肥增加了土壤碳库中难分解复杂碳化合物[12]。根据Arrhenius方程, 分解复杂化合物需要更高的活化能, 因此具有比简单化合物更高的温度敏感性[4]。同时, 试验结果也表明Q10与土壤无机氮含量表现出显著的相关性(图 6)。虽然施肥显著增加了OF处理土壤NH4+-N和NO3--N含量, 但施肥对土壤呼吸Q10的影响并不显著。这可能是由于除土壤异养呼吸外, 土壤呼吸还包括植株根系自养呼吸和土壤动物呼吸, 而油茶根系自养呼吸温度敏感性主要受温度、湿度和光合作用速率的影响[26]。然而, 本研究并未对分析施肥对土壤微生物活动、土壤氧化酶、土壤碳库组分等的影响, 仅初步探讨了施肥对油茶园土壤呼吸和异养呼吸及其温度敏感性的影响, 有关影响机理需要进一步研究。
| [1] | IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change//Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M, Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2013: 1535. |
| [2] | 杨庆朋, 徐明, 刘洪升, 王劲松, 刘丽香, 迟永刚, 郑云普. 土壤呼吸温度敏感性的影响因素和不确定性. 生态学报, 2011, 31(8): 2301–2311. |
| [3] | Zhou L Y, Zhou X H, Zhang B C, Lu M, Luo Y Q, Liu L L, Li B. Different responses of soil respiration and its components to nitrogen addition among biomes:a meta-analysis. Global Change Biology, 2014, 20(7): 2332–2343. DOI:10.1111/gcb.2014.20.issue-7 |
| [4] | Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165–173. DOI:10.1038/nature04514 |
| [5] | Zhong Y Q W, Yan W M, Shangguan Z P. The effects of nitrogen enrichment on soil CO2 fluxes depending on temperature and soil properties. Global Ecology and Biogeography, 2016, 25(4): 475–488. DOI:10.1111/geb.2016.25.issue-4 |
| [6] | Cleveland C C, Townsend A R. Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(27): 10316–10321. DOI:10.1073/pnas.0600989103 |
| [7] | Allison S D, Martiny J B H. Resistance, resilience, and redundancy in microbial communities. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(S1): 11512–11519. |
| [8] | Janssens I A, Dieleman W, Luyssaert S, Subke J-A, Reichstein M, Ceulemans R, Ciais P, Dolman A J, Grace J, Matteucci G, Papale D, Piao S L, Schulze E-D, Tang J, Law B E. Reduction of forest soil respiration in response to nitrogen deposition. Nature Geoscience, 2010, 3(5): 315–322. DOI:10.1038/ngeo844 |
| [9] | Mo J M, Zhang W, ZhuW X, Fang Y T, Li D J, Zhao P. Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China. Plant and Soil, 2007, 296(1/2): 125–135. |
| [10] | Wang Y S, Cheng S L, Fang H J, Yu G R, Xu X F, Xu M J, Wang L, Li X Y, Si G Y, Geng J, He S. Contrasting effects of ammonium and nitrate inputs on soil CO2 emission in a subtropical coniferous plantation of southern China. Biology and Fertility of Soils, 2015, 51(7): 815–825. DOI:10.1007/s00374-015-1028-x |
| [11] | Stone M M, Weiss M S, Goodale G L, Adams M B, Fernandez I J, German D P, Allison S D. Temperature sensitivity of soil enzyme kinetics under N-fertilization in two temperate forests. Global Change Biology, 2012, 18(3): 1173–1184. DOI:10.1111/j.1365-2486.2011.02545.x |
| [12] | Cusack D F, Torn M S, McDowell W H, Silver W L. The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Global Change Biology, 2010, 16(9): 2555–2572. |
| [13] | 唐健, 李娜, 欧阳洁英, 曹继钊, 覃其云. 油茶苗期生物量积累及营养分配规律研究. 南方农业学报, 2011, 42(8): 964–967. |
| [14] | Hanson P J, Edwards N T, Garten C T, Andrews J A. Separating root and soil microbial contributions to soil respiration:A review of methods and observations. Biogeochemistry, 2000, 48(1): 115–146. DOI:10.1023/A:1006244819642 |
| [15] | Subke J A, Inglima I, Cotrufo M F. Trends and methodological impacts in soil CO2 efflux partitioning:A meta-analytical review. Global Change Biology, 2006, 12(6): 921–943. DOI:10.1111/gcb.2006.12.issue-6 |
| [16] | Wang Y D, Li Q K, Wang H M, Wen X F, Yang F T, Ma Z Q, Liu Y F, Sun X M, Yu G R. Precipitation frequency controls interannual variation of soil respiration by affecting soil moisture in a subtropical forest plantation. Canadian Journal of Forest Research, 2011, 41(9): 1897–1906. DOI:10.1139/x11-105 |
| [17] | Wang H, Liu S R, Wang J X, Shi Z M, Lu L H, Zeng J, Ming A G, Tang J X, Yu H L. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China. Forest Ecology and Management, 2013, 300: 4–13. DOI:10.1016/j.foreco.2012.04.005 |
| [18] | Sheng H, Yang Y S, Yang Z J, Chen G S, Xie J S, Guo J F, Zou S Q. The dynamic response of soil respiration to land-use changes in subtropical China. Global Change Biology, 2010, 16(3): 1107–1121. DOI:10.1111/gcb.2010.16.issue-3 |
| [19] | 王超, 黄蓉, 杨智杰, 刘强, 陈光水, 万晓华. 万木林保护区柑橘和锥栗园土壤呼吸的比较. 应用生态学报, 2012, 23(6): 1469–1475. |
| [20] | Liu H, Zhao P, Lu P, Wang Y S, Lin Y B, Rao X Q. Greenhouse gas fluxes from soils of different land-use types in a hilly area of South China. Agriculture, Ecosystems & Environment, 2008, 124(1/2): 125–135. |
| [21] | Iqbal J, Hu R G, Du L J, Lu L, Lin S, Chen T, Ruan L L. Differences in soil CO2 flux between different land use types in mid-subtropical China. Soil Biology and Biochemistry, 2008, 40(9): 2324–2333. DOI:10.1016/j.soilbio.2008.05.010 |
| [22] | Deng Q, Zhou G, Liu J, Liu S, Duan H, Zhang D. Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences, 2010, 7(1): 315–328. DOI:10.5194/bg-7-315-2010 |
| [23] | Hamdi S, Moyano F, Sall S, Bernoux M, Chevallier T. Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions. Soil Biology and Biochemistry, 2013, 58: 115–126. DOI:10.1016/j.soilbio.2012.11.012 |
| [24] | 邬建红, 潘剑君, 葛序娟, 王恒钦, 余文飞, 李炳亚. 不同土地利用方式下土壤有机碳矿化及其温度敏感性. 水土保持学报, 2015, 29(3): 130–135. |
| [25] | Winkler J P, Cherry R S, Schlesinger W H. The Q10 relationship of microbial respiration in a temperate forest soil. Soil Biology and Biochemistry, 1996, 28(8): 1067–1072. DOI:10.1016/0038-0717(96)00076-4 |
| [26] | 盛浩, 杨玉盛, 陈光水, 高人, 曾宏达, 钟羡芳. 植物根呼吸对升温的响应. 生态学报, 2007, 27(4): 1596–1605. |