 2018, Vol. 38
2018, Vol. 38文章信息
- 胡媛媛, 朱纪元, 闫龙, 曹阳, 高梅香, 卢廷玉.
- HU Yuanyuan, ZHU Jiyuan, YAN Long, CAO Yang, GAO Meixiang, LU Tingyu.
- 温带落叶阔叶林地表鞘翅目成虫小尺度空间格局动态分析
- Analysis of the dynamic spatial pattern of adult Coleoptera communities at fine scale in a temperate deciduous broad-leaved forest
- 生态学报. 2018, 38(5): 1841-1851
- Acta Ecologica Sinica. 2018, 38(5): 1841-1851
- http://dx.doi.org/10.5846/stxb201702200272
-
文章历史
- 收稿日期: 2017-02-22
- 网络出版日期: 2017-11-21

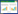




2. 黑龙江省普通高等学校地理环境遥感监测重点实验室, 哈尔滨 150025
2. Key Laboratory of Remote Sensing Monitoring of Geographic Environment, College of Heilongjiang Province, Harbin Normal University, Harbin 150025, China
空间格局与过程是土壤动物群落生态学研究的重要内容, 也是揭示生态系统生物多样性维持机制的重要前提[1]。国外学者对土壤生物群落空间异质性研究相对较早[2-3], 我国相关研究起步较晚。目前这些研究多集中于土壤属性空间异质性[4-5]、地面植被与土壤属性空间异质性的空间关联[6]、植被细根生物量空间异质性[7]等方向上, 土壤动物空间异质性研究明显落后于环境因子空间异质性研究。尺度是生态学的核心研究问题之一, 土壤动物作为生态系统中的消费者和分解者, 在不同的空间尺度上表现为不同的空间异质性特征[8]。近来研究表明, 土壤动物在小尺度空间具有多尺度空间自相关性, 并形成明显的空间异质性结构[1, 6]。但有关小尺度空间土壤动物群落空间异质性动态特征的研究仍然十分缺乏。
地表鞘翅目昆虫是土壤动物的重要组成部分, 大多以不同形态在生活史的一个时期或全部时期生活在土壤中[9], 在维持生物多样性和生态系统功能方面具有重要作用[10]。本研究以黑龙江省帽儿山森林生态系统定位站为研究平台, 以地表鞘翅目成虫为研究对象, 基于地统计学空间分析方法, 通过多次野外调查, 研究温带落叶阔叶林地表鞘翅目成虫小尺度空间格局的时间变异性特征, 揭示这种小尺度空间格局的时间动态变化规律, 促进小尺度空间土壤动物空间格局与建构机制的研究进展。
1 研究地区与研究方法 1.1 研究区概况研究区位于黑龙江省帽儿山森林生态系统研究站(45°24′N, 127°40′E)。平均海拔400m, 平均坡度15°。属温带大陆性季风气候, 夏季温暖湿润, 冬季寒冷干燥, 年均温约3.1℃[11], 早霜期一般出现在8月末, 晚霜期一般在5月末结束, 降雨期集中在7、8月份, 年均降水量为629mm[12]。实验样地设置在林龄约为60年的温带落叶阔叶林内, 主林层高约18m, 土壤为典型暗棕壤。地面植被主要包括毛榛(Corylus mandshurica)、榆(Ulmus pumila)、金刚鼠李(Rhamnus diamantiaca)、稠李(Padus avium)、白桦(Betula platyphylla)、丁香(Syringa oblata)、蚊子草(Filipendula palmata)、狭叶荨麻(Urtica angustifolia)、东北羊角芹(Aegopodium alpestre)、舞鹤草(Maianthemum bifolium)和山茄子(Brachybotrys paridiformis)等。
1.2 样地设置与野外调查方法于2014年6月在研究区内, 选择相对平坦的地段随机设置3个彼此间隔50m以上的5m×5m样地, 每个样地地表植被覆盖度95%以上, 分别记为样地A、B、C, 同时将每个样地以1m为间隔均分成25个小单元格。于2014年8月(采样期间平均气温与降水情况:18—26℃、晴-阵雨)、10月(0—15℃、多云-晴)和2015年6月(13—24℃、中雨-晴)、8月(19—28℃、晴-阵雨)、10月(1—16℃、晴-阵雨)使用陷阱法捕获地表鞘翅目昆虫。由于饱和NaCl溶液无毒无刺激性气味, 不易挥发, 不会影响土壤动物的正常活动, 防止对环境污染的同时降低保存液对地表鞘翅目成虫的引诱作用[13]。因此, 本实验用土钻(内径7cm)在网格交叉点挖一个大小合适的土坑, 然后安放内置饱和NaCl溶液的塑料杯(内径10cm、高14cm), 并加入1滴洗涤剂。保持杯口与地面齐平, 在杯口上部距离地面约10cm处支起1个一次性餐盘, 以防止凋落物、雨水等其他杂物的进入。将陷阱放置野外7d后取回, 每次调查共计75个陷阱(25个陷阱/样地×3个样地)。室内采用手捡法分拣鞘翅目昆虫, 并将样品置于95%医用酒精内。参考《中国土壤动物检索图鉴》[14]、《中国大步甲》[15]、《中国东北的葬甲科研究》[16]、《原色中国东北土壤甲虫图鉴—步行虫类》[17]、《原色中国东北土壤甲虫图鉴—隐翅虫类、拟步甲类》[18]进行分类鉴定。葬甲科鉴定到种, 步甲科大部分鉴定到种, 只有个别体型较小的个体(统称为小型步甲)无法鉴定到种, 其他鉴定到科。成虫与幼虫分别计数, 只将鞘翅目成虫数据用于后续分析。2015年8月样地受到当地村民牲畜的干扰, 该次数据不用于分析。
1.3 数据处理分析单因素方差分析比较鞘翅目成虫群落类群数量和个体数量的差异显著性。采用变异系数(CV)衡量调查月份的平均状况和总变异程度, CV < 0.1为弱变异, 0.1—1.0之间为中等变异, >1.0为强变异[19]。
1.3.1 空间自相关性分析空间自相关是指同一个变量在不同空间位置上的相关性[20], 本文使用全局Moran′s I系数(Global Moran′s I index)揭示空间邻近的区域单元属性值的相似程度, 定量描述地表鞘翅目成虫群落及类群丰富度在空间上的依赖关系, 公式为:
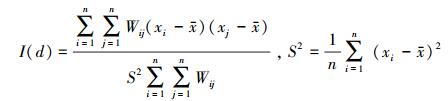
|
(1) |
式中, n是每个分离距离内样本位点的数目, xi和xj分别为样品i和样品j的观测值, x为所有样品观测值的平均值, Wij为空间权重矩阵值, 表示空间单元i和j的空间关系, 当空间单元i和j为邻近的空间单元时, Wij=1, 反之Wij=0。Moran′s I系数使用2.5m的活跃滞后距离(研究样地长度的一半), 定义第一滞后距离为1m(取样尺度), 滞后间隔为0.5m[21], Moran′s I的值域为[-1, 1], Moran′s I值为0时, 代表不存在空间自相关;Moran′s I值大于0, 说明全局自相关性是正相关, 小于0为负相关关系, 其绝对值越大, 说明自相关程度越大。通常当|I|>0.3时, 认为变量具有显著的空间自相关性[22]。
1.3.2 半方差函数及理论模型拟合半方差函数(Semivariogram)是地统计学应用最广泛的空间格局描述的基本工具[23], 用于估计地表鞘翅目成虫群落及类群丰富度的变异函数, 公式为:
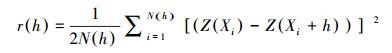
|
(2) |
式中, γ(h)为间距为h的半方差函数值, 在一定范围内随h的增加而增加;h为两分隔样点的距离;N(h)是被分隔的数据对的数量, Z(Xi)和Z(Xi +h)分别为区域变量Z(X)在点Xi和(Xi +h)处的属性值。通过变异函数的基台值(C0+C, sill)、块金值(C0, nugget)、结构比(C/(C0+C), Proportion)和变程(A, Range)定量描述空间异质性程度、组成、尺度及格局的特征。其中基台值表示样本总变量, 块金值代表了由实验误差和小于最小取样尺度(本实验为1m)引起的随机变异。结构比表示自相关部分空间异质性占总空间异质性的程度, 当(C/(C0+C)) < 0.25时空间相关性弱, 表明随机部分引起的空间变异起主要作用;(C/(C0+C))介于0.25—0.75之间时空间相关性中等, 其空间变异由随机性因素和结构性因素共同决定;(C/(C0+C)) >0.75时空间相关性强, 其空间变异主要由结构性因素引起[24-25]。变程表示研究变量空间变异中空间自相关变异的尺度范围, 在变程内空间越靠近的点之间其相关性越大, 距离大于变程的点之间不具备自相关性。
1.3.3 普通克里格空间插值普通克里格空间插值法以空间自相关为基础, 利用原始数据和半方差函数的结构性, 对区域化变量未知点进行线性无偏最优估值, 是地统计学中最常用的插值方法之一[26], 其估计方差反映了取样本身的变异以及空间内插所用分散样点内在的不确定性[27]。本文使用普通克里格空间插值绘制地表鞘翅目成虫群落丰富度的空间格局分布图, 公式为:
|
(3) |
式中, Z(x0)是在未经观测的点x0上的内插估计值, Z(xi)是在点x0附近的若干观测点上获得的实测值。
1.3.4 交叉方差函数分析交叉方差函数(Crossvariogram)用于分析鞘翅目成虫类群之间空间格局的空间关联性[28-29], 公式为:
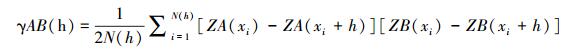
|
(4) |
式中, γAB(h)为相距为h的变量A与变量B之间的交叉方差函数值;N(h)为相隔距离为h的所有点的配对数;ZA(xi)-ZA(xi+h)分别是变量A在xi和(xi+h)处的观测值;ZB(xi)-ZB(xi+h)分别是变量B在xi和(xi+h)处的观测值。
1.3.5 数据处理软件平台群落水平分析所有捕获的地表鞘翅目成虫, 类群水平仅分析葬甲科和步甲科(除小型步甲外)。数据正态性用Kolmogorov-Smirnov检验, 对不满足正态分布的数据通过平方根转换改善样本的非正态性, 并基于转换后的数据进行后续的空间异质性分析。单因素方差分析在SPSS21.0中完成。空间自相关性、半方差函数和交叉方差函数分析在软件GS+9.0中实现。普通克里格空间插值及插值图的生成是由GS+9.0和Surfer 11.0共同完成的。
2 结果与分析4次调查共捕获鞘翅目成虫1021只, 隶属于11科29类。样地C类群数量和个体数量最多;样地A类群数量和个体数量最少。其中优势科为步甲科(Carabidae)和隐翅虫科(Staphylinidae), 常见科为葬甲科(Silphidae)、小葬甲科(Leiodidae)、闫甲科(Histeridae), 其他为稀有科。所有调查月份鞘翅目成虫群落的变异系数均大于2, 为强变异。不同科的鞘翅目成虫的个体数量在不同月份之间有显著差异(P < 0.05), 在不同样地间也差异显著(P < 0.05)。表明小尺度空间地表鞘翅目成虫群落组成具有明显的时空变化特征。
2.1 鞘翅目成虫群落及类群的空间自相关性群落水平:2014年10月样地A无空间自相关, 其他月份鞘翅目成虫丰富度均具空间正自相关性, 其中2014年8月样地B、C, 2015年6月样地A、B、C和2015年10月样地B在整个尺度上均为显著地空间正自相关性(表 1)。
| 时间-样地 Time-sample |
分割距离 Separation distance/m | |||
| 1 | 1.5 | 2 | 2.5 | |
| 2014年8月-样地A August 2014-sample A | 0.2091 | 0.1996 | 0.2275 | 0.2190 |
| 2014年8月-样地B August 2014-sample B | 0.4514* | 0.5080* | 0.4996* | 0.4472* |
| 2014年8月-样地C August 2014-sample C | 0.3056* | 0.3844* | 0.3794* | 0.3277* |
| 2014年10月-样地A October 2014-sample A | 0 | 0 | 0 | 0 |
| 2014年10月-样地B October 2014-sample B | 0.2191 | 0.2739 | 0.1709 | 0.1826 |
| 2014年10月-样地C October 2014-sample C | 0 | 0.3427* | 0.3655* | 0.2284 |
| 2015年6月-样地A June 2015-sample A | 0.3883* | 0.3542* | 0.3637* | 0.3662* |
| 2015年6月-样地B June 2015-sample B | 0.9621* | 0.9519* | 0.9594* | 0.9280* |
| 2015年6月-样地C June 2015-sample C | 0.6897* | 0.7112* | 0.6791* | 0.6978* |
| 2015年10月-样地A October 2015-sample A | 0.1361 | 0.2088 | 0.3515* | 0.3097* |
| 2015年10月-样地B October 2015-sample B | 0.5831* | 0.6017* | 0.7508* | 0.7035* |
| 2015年10月-样地C October 2015-sample C | 0.4389* | 0.2814 | 0.1760 | 0.2558 |
| *表示鞘翅目成虫群落空间自相关性显著 | ||||
类群水平:所有类群均为空间正自相关。其中2014年8月样地A的Aulonocarabus canaliculatus;2014年8月样地B的Morphocarabus venustus、Aulonocarabus canaliculatus;2014年8月样地C的Pterostichus maoershanensis、Aulonocarabus canaliculatus和2015年6月样地A的Pterostichus maoershanensis、Carabus billbergi;2015年6月样地C的Carabus billbergi、Carabus arvensis在整个空间尺度上均为显著地空间正自相关, 这些物种均为调查月份各样地的优势物种或常见物种。其他存在空间自相关性的类群, 仅在某一空间尺度上具有显著的空间正自相关性, 或在整个空间尺度上均为不显著的空间正自相关性。各月份各样地具有显著空间自相关的类群的比例分别为:2014年8月样地A(22%)、B(31%)、C(33%);2014年10月样地C(0);2015年6月样地A(44%)、B(40%)、C(46%);2015年10月样地A(25%)、B(50%)、C(67%)(表 2)。
| 时间 Time |
类群 Group |
分割距离 Separation distance/m | |||
| 1 | 1.5 | 2 | 2.5 | ||
| 2014年8月 August 2014 | QC7 | 0.1112 | 0.1390 | 0.4181* | 0.5381* |
| QC11 | 0.8455* | 0.5033* | 0.7629* | 0.5542* | |
| QC13 | 0.0625 | 0.0469 | 0.0500 | 0.0625 | |
| QC15 | 0.2000 | 0.1563 | 0.1833 | 0.1875 | |
| QC17 | 0.1746 | 0.1486 | 0.1331 | 0.1455 | |
| QC18 | 0.4648* | 0.6409* | 0.8163* | 0.7395* | |
| QC20 | 0.0375 | 0.0313 | 0.0333 | 0.0313 | |
| 2014年10月 October 2014 | QC11 | 0 | 0 | 0 | 0.2831 |
| 2015年6月 June 2015 | QC9 | 0.4424* | 0.2287 | 0.1730 | 0.1524 |
| QC11 | 0.4425* | 0.7150* | 0.1214 | 0.4574* | |
| QC13 | 0.9020* | 0.9281* | 0.5836* | 0.8133* | |
| QC14 | 0.6340* | 0.6847* | 0.5438* | 0.4807* | |
| QC16 | 0 | 0 | 0.3157* | 0.1973 | |
| QC18 | 0 | 0 | 0 | 0.3946* | |
| 2015年10月 October 2015 | QC11 | 0.3445* | 0 | 0 | 0 |
| QC13 | 0.3445* | 0 | 0 | 0 | |
| QC1:北方花葬甲, Nicrophorus tenuipes Lewis;QC2:达乌里花葬甲, Nicrophorus dauricus Motschulsky;QC3:四星花葬甲, Nicrophorus quadripunctatus Kraatz;QC4:红斑花葬甲, Nicrophorus vespilloides Herbst;QC5:黄斑花葬甲, Nicrophorus praedatro Reitter;QC6:六脊树葬甲, Dendroxena sexcarinata Motschulsky;QC7:黑光葬甲, Phosphuga atrata Linnaeus;QC8:随葬甲, Silpha perforata perforata Gebler;QC9:小葬甲, Sciodrepoides fumatus Spence;QC10:Pterostichus (Metallophilus) heilongjiangensis;QC11:Pterostichus maoershanensis;QC12:Pterostichus audax;QC13:Carabus billbergi;QC14:Carabus arvensis;QC15:Morphocarabus venustus;QC16:Tomocarabus fraperculus;QC17:金边步甲, Megodontus vietinghoffi;QC18:沟步甲, Aulonocarabus canaliculatus;QC19:Amara sp.;QC20:Acoptolabrus schrencki;QC21:Acoptolabrus constricticollis, *表示鞘翅目成虫类群空间自相关性显著; 表格未出现的类群在整个研究尺度上均不存在空间自相关性 | |||||
鞘翅目成虫群落的最优拟合理论模型:2015年6月样地A和2015年10月样地C为球状模型, 2015年10月样地A、B为指数模型, 其他均为线性模型(表 3)。所有群落的C0均大于0, 说明鞘翅目成虫群落丰富度的半方差图中均存在块金方差, 即存在由采样误差、微尺度空间变异等因素带来的块金效应。群落结构比C/(C0+C)在2015年10月样地C介于0.25与0.75之间, 表明为中等程度的空间自相关性, 且其空间变异由随机性因素和结构性因素共同决定;在2015年6月样地A、2015年10月样地A、B均大于0.75, 表明空间自相关性很强, 结构性因素引起的空间异质性对这种格局起主要作用;其他群落均为0, 为纯块金效应, 说明样地中空间变异完全由随机因素决定。
| 时间-样地 Time-sample |
模型 Model |
块金值 Nugget variance (C0) |
基台值 Structure variance sill (C0+C) |
结构比 Proportion [C/(C0+C)] |
残差 Residual sum of squares (RSS) |
决定系数 Coefficient of determination (R2) |
变程A Range/m |
| 2014年8月-样地A August 2014-sample A |
Lin. | 1.004 | 1.004 | 0 | 0.0123 | 0.388 | 2.236 |
| 2014年8月-样地B August 2014-sample B |
Lin. | 0.959 | 0.959 | 0 | 0.247 | 0.081 | 2.236 |
| 2014年8月-样地C August 2014-sample C |
Lin. | 1.246 | 1.246 | 0 | 0.534 | 0.023 | 2.236 |
| 2014年10月-样地A October 2014-sample A |
Lin. | 0.123 | 0.123 | 0 | < 0.001 | 0.105 | 2.236 |
| 2014年10月-样地B October 2014-sample B |
Lin. | 0.284 | 0.284 | 0 | < 0.001 | 0.882 | 2.236 |
| 2014年10月-样地C October 2014-sample C |
Lin. | 0.190 | 0.190 | 0 | < 0.001 | 0.067 | 2.236 |
| 2015年6月-样地A June 2015-sample A |
Sph. | 0.097 | 1.116 | 0.913 | 0.0176 | 0.406 | 1.430 |
| 2015年6月-样地B June 2015-sample B |
Lin. | 0.758 | 0.758 | 0 | 0.029 | 0.002 | 2.236 |
| 2015年6月-样地C June2015-sampleC |
Lin. | 1.376 | 1.376 | 0 | 0.690 | 0.049 | 2.236 |
| 2015年10月-样地A October 2015-sample A |
Exp. | 0.049 | 0.313 | 0.841 | < 0.001 | 0.098 | 1.041 |
| 2015年10月-样地B October 2015-sample B |
Exp. | 0.007 | 0.316 | 0.977 | 0.977 | 0.338 | 1.374 |
| 2015年10月-样地C October 2015-sample C |
Sph. | 0.104 | 0.363 | 0.713 | 0.01 | 0.325 | 2.177 |
| Exp:指数模型, Exponential;Sph:球状模型, Spherical;Lin:线性模型, Linear | |||||||
类群水平:除2014年8月样地B、2015年6月样地B、2015年10月样地A、C外, 各调查群落均有一半及以上的类群拟合为线性模型, 尤其样地C在2014年8月和2015年6月这两次调查中有100%和76.92%的类群拟合为线性模型(表 4), 结构比为0。各调查月份3个样地变程小于1m的类群只出现在2014年8月和2015年6月这两次调查中, 变程大于4m的类群只出现于2014年10月和2015年10月这两次调查中。
| 时间-样地 Time-sample |
类群 Groups |
模型 Model |
块金值 Nugget variance (C0) |
基台值 Structure variance sill (C0+C) |
结构比 Proportion [C/(C0+C)] |
残差 Residual sum of squares (RSS) |
决定系数 Coefficient of determination (R2) |
变程A Range/m |
| 2014年10月 October 2014 | QC10 | Gau. | 0.041 | 0.124 | 0.666 | < 0.001 | 0.872 | 4.971 |
| 2015年6月 June 2015 | QC9 | Sph. | 0.022 | 0.218 | 0.896 | < 0.001 | 0.999 | 2.120 |
| QC14 | Sph. | 0.010 | 0.529 | 0.981 | < 0.001 | 0.000 | 1.000 | |
| QC15 | Exp. | 0.000 | 0.050 | 0.998 | < 0.001 | 0.016 | 1.554 | |
| 2015年10月 October 2015 | QC11 | Sph. | 0.000 | 0.077 | 0.999 | < 0.001 | 0.886 | 2.166 |
| QC13 | Sph. | 0.000 | 0.058 | 0.998 | < 0.001 | 0.732 | 2.147 | |
| QC18 | Exp. | 0.000 | 0.025 | 1.000 | < 0.001 | 0.016 | 1.545 | |
| Exp.:指数模型, Exponential;Sph.:球状模型Spherical;Gau.:高斯模型, Gaussian; 调查月份未在表格中出现的类群均拟合为线状模型, 结构比为0;类群代码见表 2 | ||||||||
鞘翅目成虫类群的空间关联性因时间和空间变化而有所差别, 表现为正的或负的或不存在空间关联性, 不同的类群之间具有不同的空间作用关系, 两个类群间的空间关联性在不同调查月份和不同调查样地中并不具备一致性和规律性, 仅有少部分类群间在同次调查不同样地中始终表现为相同的空间关联性。结构比显示类群之间的空间关联性形成主要是结构性因素或随机性因素单一调控的结果, 各调查月份均存在一些类群不存在空间关联性, 样地C尤为明显(表 5)。各样地在每个调查月份都有近一半或超过一半的类群之间是负相关, 负相关的比例分别为:2014年8月样地A(63%)、B(43%)、C(50%);2014年10月C(100%);2015年6月样地A(36%)、B(50%)、C(36%);2015年10月样地A(33%)、B(83%)、C(50%)。
| 类群 Groups |
空间关联性 Spatial relationship |
决定系数 Coefficient of determination (R2) |
结构比 Proportion [C/(C0+C)] |
变程A Range/m |
| QC3×QC7 | 负 | 0.353 | 0.999 | 8.556 |
| QC3×QC10 | 负 | 0.034 | 0 | 2.236 |
| QC3×QC11 | 负 | 145 | 0.999 | 1.643 |
| QC3×QC15 | 负 | 0.518 | 0.997 | 2.31 |
| QC3×QC17 | 负 | 0.522 | 0.999 | 10.407 |
| QC3×QC18 | 正 | 0 | 0.982 | 1.152 |
| QC7×QC10 | 负 | 0.121 | 1 | 1.479 |
| QC7×QC11 | 负 | 0.285 | 0 | 2.236 |
| QC7×QC13 | 正 | 0.755 | 0 | 2.236 |
| QC7×QC15 | 正 | 0.563 | 0 | 2.236 |
| QC7×QC17 | 正 | 0.038 | 0.792 | 1.402 |
| QC7×QC18 | 正 | 0.162 | 0 | 2.236 |
| QC7×QC20 | 负 | 0.457 | 0 | 2.2336 |
| QC10×QC11 | 正 | 0.085 | 0.605 | 9.558 |
| QC10×QC15 | 负 | 0.522 | 0.998 | 10.407 |
| QC10×QC18 | 负 | 0.193 | 0 | 2.236 |
| QC11×QC15 | 正 | 0.983 | 0 | 2.236 |
| QC11×QC17 | 负 | 0.203 | 0.999 | 1.567 |
| QC11×QC18 | 负 | 0 | 0.005 | 2.236 |
| QC11×QC20 | 正 | 0.067 | 0.999 | 1.594 |
| QC13×QC15 | 负 | 0.15 | 0.999 | 1.732 |
| QC13×QC18 | 正 | 0.608 | 0.856 | 5.51 |
| QC13×QC20 | 负 | 0.62 | 0 | 2.236 |
| QC15×QC17 | 负 | 0.796 | 0 | 2.236 |
| QC15×QC18 | 负 | 0.726 | 0.999 | 5.786 |
| QC15×QC20 | 负 | 0.522 | 0.999 | 10.407 |
| QC17×QC18 | 负 | 0.161 | 0.974 | 1.406 |
| QC17×QC22 | 负 | 0.179 | 1 | 3.37 |
| QC17×QC24 | 正 | 0.074 | 0 | 2.236 |
| QC17×QC29 | 负 | 0.62 | 0 | 2.236 |
| QC18×QC20 | 正 | 0.989 | 0.999 | 10.407 |
| “正”表示正相关性, “负”表示负相关性, 在表格中未出现的类群之间均不存在相关性 | ||||
鞘翅目成虫群落的空间分布格局梯度特征较明显, 形成高值斑块与低值斑块镶嵌分布的空间格局(图 1)。相同调查季节, 样地A、B、C的空间分布格局并不完全一致。3个样地在6、8月具有较复杂的空间分布格局, 较多的高值斑块和低值斑块复杂镶嵌;而在10月则表现为相对简单的空间分布格局, 仅有几个斑块简单镶嵌。对于单个样地来说, 不同调查时间其空间分布格局表现不同, 具有动态变化性, 但是各样地边缘大多有高值斑块区分布。

|
| 图 1 鞘翅目成虫群落空间分布格局 Fig. 1 Spatial distribution patterns of Coleoptera adult communities |
调查月份鞘翅目群落始终体现为空间正自相关, 且丰富度较大的类群通常具有显著地空间正自相关性。就各月份各样地存在显著自相关性的类群比例看:样地间百分比数值的不同体现了样地间的差异性;2014年10月样地C是0, 2015年10月则相对较高, 体现了相同月份在不同年际也可能存在差异;同一样地, 4次调查月份的不同, 体现了类群空间自相关性特征具有时间动态性。受距离约束的空间自相关分析结果表明, 可能是受到某些小尺度干扰或各向异性导致空间自相关性的局部变化, 复杂的变化反映了鞘翅目成虫空间结构的特殊性以及影响因子的复杂性[30]。
鞘翅目成虫群落大部分拟合为线性模型, 表现为块金效应, 说明研究尺度下鞘翅目群落的空间分布多为随机分布, 这可能与采样尺度和类群丰富度有关, 进一步研究须加大采样密度。接近一半的类群的空间变异是结构性因素或结构性因素和随机性因素共同调控的结果, 可能受到如环境筛选、种间竞争以及自身生物学特性等多方面的控制[31]。各调查月份各样地都有接近甚至超过一半类群的结构比为0, 表现为纯块金效应, 即存在由实验误差(陷阱法反映的是类群的活动性密度[32]、诸如植被类型、植被密度等一些影响类群活动性的因素会给取样结果带来误差)、小于取样尺度和微尺度空间变异等因素带来的块金效应, 说明小于采样尺度(1m)的空间过程不容忽视, 可在现有的空间尺度内, 缩小空间粒度并加大采样密度进行研究。
两个类群的空间关联性在相同调查月份不同样地上不具稳定性, 在同一样地不同调查月份也不具稳定性。鞘翅目成虫类群间存在复杂的或正或负或不存在的空间关联性。结构比表明这种空间作用大多受结构性因素或随机性因素单一调控。各样地各调查月份都有近一半或超过一半的类群之间是负相关, 这可能与类群间重叠的生态位、食性类型相似引起的竞争有关, 比如Nicrophorus quadripunctatus Kraat与Phosphuga atrata Linnaeus这两个类群同属葬甲科, 可能会因食物喜好相近导致大多以负相关关系出现。在同一调查月份样地A、B、C的类群间关联性呈正相关、负相关、不相关的比例差别较大, 反映出类群空间关联性的复杂性和不确定性。同一样地在同一年的两次调查中负相关所占的比例均随采样季节推移而变大, 这说明由多重原因(如温度)导致的类群丰富度的变化可能会对种间关联性均产生影响, 也表现出类群之间的合作在这个尺度可能并不十分明显。另本文的类群水平仅分析葬甲科和除去小型步甲的步甲科昆虫, 并不是实验所获得全部鞘翅目昆虫, 这可能会对以上类群水平的分析产生影响, 以后还需对更精细的数据进行分析。
鞘翅目成虫丰富度空间分布不均匀, 群落具有明显的空间异质性特征。随着时间的变化, 斑块复杂程度变化较大, 斑块的形状和大小均存在较大差异, 这种复杂性和不确定性均体现出鞘翅目群落的空间异质性具有动态性特征, 这与半方差函数得出鞘翅目成虫群落大多受随机因素影响相符。对比朱纪元等[33]在帽儿山人工红松林针对20m×20m尺度下鞘翅目群落的调查:两个研究均表现出梯度特征明显的空间分布格局, 可进一步说明“高值”斑块与“低值”斑块并存的空间格局可能是鞘翅目成虫分布的典型特征。本研究空间格局的复杂程度相对较低, 且群落空间异质性的形成中较多为随机性因素控制。可见调查尺度变小, 随机性因素调控能力加强。这种现象的形成可能与鞘翅目昆虫的扩散、迁移能力较强, 活动距离较大有关。近些年来, 已有研究表明土壤水分、土壤表层温度、凋落物厚度及土壤养分等本身也存在明显的空间异质性[34-37, 26], 刘继亮[38]等的研究表明在小尺度上, 林地的坡向和微地形条件主要通过影响局部小生境的生物和非生物因子组合特征而对土壤动物的空间分布格局产生重要影响。Sharavari等[39]的研究表明甲虫的活动密度与杂草的高密度斑块之间有着显著的重叠。Parisi等[40]研究发现甲虫群落受森林结构的影响。另外, 本实验为开放样地, 样地边缘的高值斑块分布区(如2014年8月和2015年6月)可能同时受到因干扰产生的边缘效应[41]的影响。然而本文并未考虑环境因子及物种迁入、迁出过程, 在今后的研究中需进一步深化该方面的研究。
4 结论本研究表明鞘翅目成虫在小尺度空间的分布往往是不均匀的, 存在着格局的异质性。在5m×5m研究尺度上, 鞘翅目成虫群落和类群具有复杂的正的空间自相关性, 表现为空间异质性特征和一定的时间动态变化特征, 且随机性因素或结构性因素和随机性因素对这种空间分异起主要调控作用, 类群(葬甲科和部分步甲科)之间呈现为复杂的负相关居多的空间关联性。群落的空间格局分布特征较明显, 但不具有时间稳定性, 体现出明显的时间动态性。本研究仅能表明5m×5m尺度下的鞘翅目成虫的空间格局, 更大或更小尺度有待于进一步研究。
| [1] | 高梅香, 刘冬, 张雪萍, 吴东辉. 三江平原农田地表和地下土壤螨类丰富度与环境因子的空间关联性. 生态学报, 2016, 36(6): 1782–1792. |
| [2] | Webster R, Boag B. Geostatistical analysis of cyst nematodes in soil. Journal of Soil Science, 2010, 43(3): 583–595. |
| [3] | Robertson G P, Gross K L. Assessing the heterogeneity of belowground resources:quantifying pattern and scale. Exploitation of Environmental Heterogeneity by Plants, 1994: 237–253. |
| [4] | Aviron S, Burel F, Baudry J, Schermann N. Carabid assemblages in agricultural landscapes:impacts of habitat features, landscape context at different spatial scales and farming intensity. Agriculture, Ecosystems & Environment, 2005, 108(3): 205–217. |
| [5] | 李从娟, 雷加强, 徐新文, 高培, 邱永志, 许波, 钟显彬, 王永东, 闫健, 王桂芬. 古尔班通古特沙漠土壤水分与化学性质的空间分布. 生态学报, 2014, 34(15): 4380–4389. |
| [6] | Gutiérrez-López M, Jesús J B, Trigo D, Fernández R, Novo M, Díaz-Cosín D J. Relationships among spatial distribution of soil microarthropods, earthworm species and soil properties. Pedobiologia, 2010, 53(6): 381–389. DOI:10.1016/j.pedobi.2010.07.003 |
| [7] | Gao Y, Mao L, Miao C Y, Zhou P, Cao J J, Zhi Y E, Shi W J. Spatial characteristics of soil enzyme activities and microbial community structure under different land uses in Chongming Island, China:Geostatistical modelling and PCR-RAPD method. Science of The Total Environment, 2010, 408(16): 3251–3260. DOI:10.1016/j.scitotenv.2010.04.007 |
| [8] | 路有成, 李景科. 皖南低丘板栗林土壤甲虫群落研究. 生态学杂志, 1994, 13(6): 1–6. |
| [9] | 闫海冰, 韩有志, 杨秀清, 李乐. 关帝山云杉天然更新与土壤有效氮素异质性的空间关联性. 应用生态学报, 2010, 21(3): 533–540. |
| [10] | 王树堂, 韩士杰, 张军辉, 王存国, 徐媛, 李雪峰, 王树起. 长白山阔叶红松林表层土壤木本植物细根生物量及其空间分布. 应用生态学报, 2010, 21(3): 583–589. |
| [11] | 焦振, 王传宽, 王兴昌. 温带落叶阔叶林冠层CO2浓度的时空变异. 植物生态学报, 2011, 35(5): 512–522. |
| [12] | 王静, 王兴昌, 王传宽. 基于不同浓度变量的温带落叶阔叶林CO2储存通量的误差分析. 应用生态学报, 2013, 24(4): 975–982. |
| [13] | 周红章, 于晓东, 罗天宏, 李晓燕, 王凤艳, 李德娥, 周毓灵子, 赵彩云. 土壤步甲和隐翅虫的采集与田间调查取样技术. 应用昆虫学报, 2014, 51(5): 1367–1375. DOI:10.7679/j.issn.2095-1353.2014.164 |
| [14] | 尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998. |
| [15] | 李景科, 张雪萍. 中国大步甲. 长春: 吉林教育出版社, 2005. |
| [16] | 李景科, 张雪萍, 林琳, 福鲁茨, 于强. 中国东北的葬甲科研究. 哈尔滨: 哈尔滨地图出版社, 2011. |
| [17] | 李井科, 林琳, 张雪萍. 原色中国东北土壤甲虫图鉴——步行虫类. 哈尔滨: 哈尔滨地图出版社, 2015. |
| [18] | 李井科, 张利敏, 张雪萍. 原色中国东北土壤甲虫图鉴——隐翅虫类、拟步甲类. 哈尔滨: 哈尔滨地图出版社, 2015. |
| [19] | Yonker C M, Schimel D S, Paroussis E, Heil R D. Patterns of organic carbon accumulation in a semiarid shortgrass steppe, Colorado. Soil Science Society of America Journal, 1988, 52(2): 478–483. DOI:10.2136/sssaj1988.03615995005200020032x |
| [20] | Cliff A D, Ord J K. Spatial Processes:Models & Applications. London: Pion Ltd, 1981. |
| [21] | Jung W K, Kitchen N R, Sudduth K A, Anderson S H. Spatial characteristics of claypan soil properties in an agricultural field. Soil Science Society of America Journal, 2006, 70(4): 1387–1397. DOI:10.2136/sssaj2005.0273 |
| [22] | 张海森, 宋向东, 邓建梅. 基于模型的Moran's I检验在信息软件业中的应用. 河北北方学院学报:自然科学版, 2008, 24(1): 66–68. |
| [23] | 王政权. 地统计学及在生态学中的应用. 北京: 科学出版社, 1999. |
| [24] | 颜亮, 周广胜, 张峰, 隋兴华, 平晓燕. 内蒙古荒漠草原植被盖度的空间异质性动态分析. 生态学报, 2012, 32(13): 4017–4024. |
| [25] | 王猛, 汪季, 蒙仲举, 柴享贤, 吕世杰, 王德慧, 乌云嘎. 巴丹吉林沙漠东缘天然梭梭种群空间分布异质性. 生态学报, 2016, 36(13): 4055–4063. |
| [26] | 苏松锦, 刘金福, 何中声, 洪伟, 张金彪. 格氏栲天然林土壤养分空间异质性. 生态学报, 2012, 32(18): 5673–5682. |
| [27] | 毕华兴, 李笑吟, 刘鑫, 李俊, 郭孟霞. 晋西黄土区土壤水分空间异质性的地统计学分析. 北京林业大学学报, 2006, 28(5): 59–66. |
| [28] | 王正军, 李典谟, 商晗武, 程家安. 地质统计学理论与方法及其在昆虫生态学中的应用. 应用昆虫学报, 2002, 39(6): 405–411. DOI:10.7679/j.issn.2095-1353.2002.116 |
| [29] | Rossi J P, Lavelle P, Tondoh J E. Statistical tool for soil biology:X. Geostatistical analysis. European Journal of Soil Biology, 1995, 31(4): 173–181. |
| [30] | 高梅香, 何萍, 刘冬, 郭传伟, 张雪萍, 李景科. 温带落叶阔叶林土壤螨群落多尺度空间自相关性. 土壤通报, 2014, 45(5): 1104–1112. |
| [31] | Ingimarsdóttir M, Caruso T, Ripa J, Magnúsdóttir Í B, Migliorini M, Hedlund K. Primary assembly of soil communities:disentangling the effect of dispersal and local environment. Oecologia, 2012, 170(3): 745–754. DOI:10.1007/s00442-012-2334-8 |
| [32] | Mitchell B. Ecology of two carabid beetles, Bembidion lampros (Herbst) and Trechus quadristriatus (Schrank)——Ⅱ. Studies on populations of adults in the field, with special reference to the technique of pitfall trapping. Journal of Animal Ecology, 1963, 32(3): 377–392. DOI:10.2307/2599 |
| [33] | 朱纪元, 李景科, 高梅香, 胡媛媛, 张雪萍. 帽儿山红松人工林鞘翅目成虫群落小尺度空间异质性变化特征. 生态学报, 2017, 37(6): 1975–1986. |
| [34] | 杨秀清, 韩有志, 李乐, 陈欣, 游静. 华北山地典型天然次生林土壤氮素空间异质性对落叶松幼苗更新的影响. 生态学报, 2009, 29(9): 4656–4664. |
| [35] | 邵方丽, 余新晓, 杨志坚, 王贺年. 天然杨桦次生林表层土壤水分与枯落物的空间异质性. 水土保持学报, 2012, 26(3): 199–204. |
| [36] | 范宁宁, 马超, 王树力. 不同经营类型红松林下土壤腐殖质的空间异质性. 水土保持学报, 2012, 26(3): 184–188. |
| [37] | 周李磊, 朱华忠, 钟华平, 杨华, 索菲娅, 邵小明, 周星杰. 新疆伊犁地区草地土壤容重空间格局分析. 草业学报, 2016, 25(1): 64–75. DOI:10.11686/cyxb2015249 |
| [38] | 刘继亮, 李锋瑞. 坡向和微地形对大型土壤动物空间分布格局的影响. 中国沙漠, 2008, 28(6): 1104–1112. |
| [39] | Kulkarni S S, Dosdall L M, Spence J R, Willenborg C J. Field density and distribution of weeds are associated with spatial dynamics of omnivorous ground beetles (Coleoptera:Carabidae). Agriculture, Ecosystems & Environment, 2017, 236: 134–141. |
| [40] | Parisi F, Lombardi F, Sciarretta A, Tognetti R, Campanaro A, Marchetti M, Trematerra P. Spatial patterns of saproxylic beetles in a relic silver fir forest (Central Italy), relationships with forest structure and biodiversity indicators. Forest Ecology and Management, 2016, 381: 217–234. DOI:10.1016/j.foreco.2016.09.041 |
| [41] | 杨贵军, 王新谱, 贾彦霞, 张大治. 人工柠条-荒漠草地交错带拟步甲昆虫群落多样性. 生态学报, 2016, 36(3): 608–619. |