 2018, Vol. 38
2018, Vol. 38文章信息
- 楚彬, 马素洁, 周延山, 姬程鹏, 周建伟, 周睿, 田永亮, 花立民.
- CHU Bin, MA Sujie, ZHOU Yanshan, JI Chengpeng, ZHOU Jianwei, ZHOU Rui, TIAN Yongliang, HUA Limin.
- 祁连山东段高原鼢鼠(Eospalax baileyi)土丘空间分布格局及其与环境因子的空间关联性
- Relationship between the spatial distribution of the mounds of plateau zokor (Eospalax baileyi) and environmental factors in eastern Qilian Mountain
- 生态学报. 2018, 38(3): 964-974
- Acta Ecologica Sinica. 2018, 38(3): 964-974
- http://dx.doi.org/10.5846/stxb201612292705
-
文章历史
- 收稿日期: 2016-12-29
- 网络出版日期: 2017-10-18

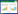




种群空间分布格局是种群对环境条件长期适应和选择的结果, 对于了解种群特征、种内和种间关系以及种群与环境关系有特殊意义[1]。目前, 动物种群的空间分布格局已有大量研究, 主要是根据经典统计学方法研究其种群和群落的格局分析, 同时也用于其数量分类和排序问题[2-4]。虽然经典统计学方法有着诸多优点, 但是在分析种群空间分布格局还是存在一定的局限性。因为种群的空间分布格局与动物选择利用栖息地的生态学过程有着紧密联系, 而生态系统的组分及其对生物生长、发育等影响是复杂的, 会在不同的空间尺度下影响动植物的空间分布格局[5]。经典统计学方法忽略了不同尺度下所揭示的生态学现象。并且经典统计学研究的变量必须是纯随机变量, 忽略了变量与地理位置的关系, 因此在研究种群空间分布格局可能会得到片面甚至错误的结果。近年来, 地理信息系统已被为生态学家所重视, 其中地统计学中的空间分析方法为确定种群空间分布格局以及揭示空间格局与环境因子的空间关联性提供了有效途径[6]。与经典统计学相比, 地统计学更注重随机变量的空间过程, 并通过研究对象在空间上不同间隔的抽样点上的差异, 定量描述了其空间变化规律, 同时, 可以揭示某一变量或两个变量在不同尺度下的空间格局及空间关系。国内关于啮齿动物种群空间分布格局的研究开展较多[7], 主要集中于单纯研究某一鼠种的空间分布。赵天彪等[8]采用最近邻体法和负二项分布法研究了大沙鼠和子午沙鼠的空间分布格局, 结果发现大沙鼠一年四季均存在聚集分布, 而子午沙鼠的空间分布随季节变化而变化。房继明和孙儒永[9]通过最近邻体法研究布氏田鼠的空间分布, 认为其在不同时期存在不同的分布类型。宗浩等[10]采用概率分布法对高原鼠兔的种群空间分布进行研究, 发现高原鼠兔在不同环境条件下分布格局会发生变化。前人的研究方法大多采用频次比较法和分布型指标法判断其分布型, 却无法知道分布的具体位置和程度, 而且这些研究方法关注的只是时间相关, 而忽略了格局内样点间的空间相关和格局间的时间相关问题, 其结果不仅造成系统误差, 而且研究的时域范围十分有限。
高原鼢鼠(Myospalax baileyi)广泛分布于青藏高原, 是青藏高原生物多样性的重要组成部分, 在草地生态系统食物网及其相对应的能量流通和物质循环中有着独特的地位[11-12], 但高原鼢鼠采食牧草以及挖掘过程中推出土丘, 影响高寒草地生产和生态功能。由于高原鼢鼠营地下生活, 确定某一种群内全部个体的空间位置存在一定难度, 而地表土丘便于识别和定位, 同时土丘数量和形成时间可间接反应种群动态变化, 因此以土丘的空间位置反映其种群空间分布是一种理想的替代方法[13]。国内关于地下鼠种群空间分布研究较少, 陈林等[14]采用聚集度指数和平均拥挤度两种指标研究景观尺度下高原鼢鼠的种群空间分布并通过相关性分析该空间分布与气候因素的关系, 研究发现所有高原鼢鼠种群空间分布格局均为聚集分布, 并且气候因素对聚集程度影响不显著。但是, 景观尺度本身就存在气候、地形和土壤的异质性, 得出的结果可能不能准确反映真实的高原鼢鼠种群空间分布与其环境因子的关系。因此, 本研究在祁连山东段高寒草甸的小尺度下, 消除景观尺度带来的环境差异性, 以高原鼢鼠土丘为研究对象, 利用地统计空间分析的方法研究其土丘空间分布格局, 并揭示这种空间格局与环境因子的空间关联性, 为合理预测高原鼢鼠扩散和保护生物多样性提供科学依据。
1 材料与方法 1.1 研究样地概况本试验在甘肃省武威市天祝藏族自治县抓喜秀龙乡高寒草甸区进行, 该地区区位于东祁连山的天祝金强河河谷, 地理坐标37°12′N, 102°46′E, 海拔2937m, 气候寒冷潮湿, 太阳辐射强。年均温-0.1℃, 1月平均温度-18.3℃, 7月平均温度12.7℃, >0℃积温1380℃;年降水量416mm, 多为地形雨, 集中于7、8、9三个月。植被类型为高寒草甸, 以莎草科、禾本科植物为主, 混有其他杂类草, 草群盖度达到90%左右, 土壤为高寒草甸土, 主要植物有珠芽蓼(Polygonum viviparum)、鹅绒委陵菜(Potentilla anserina)、早熟禾(Poaceae annua)、矮嵩草(Kobresia humilis)、线叶嵩草(Kobresia capillifoli)、垂穗披碱草(Elymus dahuricus)、球花蒿(Artemisia smithii Mattf)、黄芪(Astragalus membranaceus)等[15]。高原鼢鼠为该地区主要优势鼠种[16]。
1.2 试验设计样地位于天祝县马营滩地区的冬季高寒草甸牧场, 高原鼢鼠是该地区唯一造丘的地下啮齿类。在该样地设置140m×100m的样方, 以10m为间隔钉入木桩标记相对坐标, 将该样地等间距划分为165个10m×10m的小样方。调查每一样方内新旧土丘数量。同时以每个小样方的对角线交叉点为中心, 测定环境因子中植物地上生物量、10—20cm地下生物量、土壤容重、水分以及根系营养物质, 选择土壤深度为10—20cm是因为研究区内高原鼢鼠洞道的平均深度为14.11cm(n=64)。野外调查于2014年8月份进行。本研究利用半方差函数分析高原鼢鼠土丘的空间分格局, 交方差函数(Crossvariogram)用于分析土丘空间格局与环境因子空间格局的空间关联性, 并进行mantel显著性检验。
1.3 测定指标及方法 1.3.1 土丘数量调查采用RTK(Real-Time Kinematic)技术记录样地内每个土丘的经纬度位置[17], 通过ArcGIS软件获取每一小样方的土丘分布及其数量。
1.3.2 土丘分布类型调查采用扩散系数(C)指标[14]确定高原鼢鼠土丘的空间分布型。
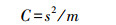
|
式中, s2为方差, m为平均土丘密度。
1.3.3 土壤容重和水分调查每一小样方的中点附近挖取1个剖面, 深度20cm, 自表土向下每隔10cm用100cm3环刀取一个土壤样品并装入铝盒, 并在中点附近50cm处重复2次, 带回实验室采用铝盒烘干法测定其容重和水分[18], 最后求取平均值。
1.3.3 地上生物量调查在每一小样方中点附近采用DWR(Dry-Weight Rank)法[19]调查地上生物量。
1.3.4 地下生物量调查采用直径为10cm根钻采集每一小样方中点处10—20cm的根系, 并将附着在根系的土洗净, 烘干根系并进行称重。
1.3.5 植物功能群丰富度调查采用DWR法[19]调查每个样点的植物学组成, 将所调查的植物划分为禾本科、莎草科和杂类草并计算每个功能群所占的比例。
1.3.6 根系可溶性碳水化合物、粗蛋白和粗脂肪含量测定将每一样点采集的10—20cm根系烘干后, 分别采用“蒽酮法”[20]测定其可溶性碳水化合物;“凯氏定氮法”[21]测定其粗蛋白和“索氏提取法”[22]测定其粗脂肪含量。
1.4 数据处理 1.4.1 半方差函数及理论模型拟合半方差函数用于分析高原鼢鼠土丘和各环境因子的空间变异程度。其是地统计学进行空间分析和揭示空间分异规律的基本数学工具, 反映了不同距离观测值之间的变化[22]。数学公式为:
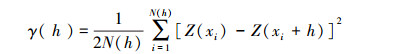
|
式中, γ(h)为间距为h的半方差函数值, 在一定范围内随h的增加而增加;h为两分隔样点的距离;N(h)是被分隔的数据对的数量, Z(xi)和Z(xi+h)分别为区域变量Z(x)在点xi和点(xi+h)处的属性值。
计算土丘数量及环境因子的变异函数值, 依据决定系数最大、残差平方和较小的原则筛选最适模型, 应用模型参数[23]定量描述土丘数量与环境因子属性值的空间异质性。块金值(C0)表示由于实验误差和小于最小取样尺度(10m)引起的随机变异。块金值与基台值之比(C0/C0+C)代表土丘和环境因子的空间依赖性, 即属性值之间的空间相关性程度, 也表示随机部分引起的空间异质性占总变异的比例。当C0/C0+C < 25%, 空间自相关性程度较强, 空间变异主要由结构性因素引起;当25% < C0/C0+C < 75%, 空间相关性中等, 其空间变异由随机性因素和结构性因素共同决定;当C0/C0+C > 75%时, 空间相关性较弱, 表明随机因素在空间变异中起主要作用[24-25]。
1.4.2 普通克里格空间插值利用环境因子和土丘属性值和半方差函数的结构性, 使用普通克里格插值对未采样点区域化变量(x0)进行最优无偏估值[23], 并对环境因子和高原鼢鼠土丘的空间分布进行制图。
1.4.3 交方差函数分析交方差函数(Crossvariogram)用于分析土丘空间格局与环境因子空间格局的空间关联性[26-27]:
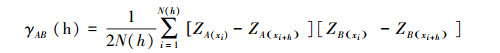
|
式中, γAB(h)为相距h的变量A与变量B之间的交方差函数值;N(h)为相距为h的所有点的配对数;ZA(xi)和ZA(xi+h)分别是变量A在xi和xi+h处的属性值;ZB(xi)和ZB(xi+h)分别是变量B在xi和xi+h处的属性值。最后使用Mantel检验分析土丘分布空间格局与环境因子空间格局空间关系的显著性[6]。
半方差函数和交方差函数在GS+10.0软件实现;克里格空间插值图和简单Mantel检验分别在ArcGIS 10.2和R(3.1.2)软件的“vegan”软件包实现。
2 结果与分析 2.1 高原鼢鼠土丘空间分布型及空间异质性特征通过对土丘的扩散系数计算, 高原鼢鼠土丘为聚集分布(C=s2/m=10.78 > 1)。对高原鼢鼠土丘的空间变异进行模型拟合, 结果表明高斯模型对高原鼢鼠土丘数量进行模拟的拟合度较高(表 1)。由表 1可看出, 土丘的空间分布存在中等强度的变异(C0/C0+C介于25%—75%之间), 这种变异由随机因素引起的所占比例为40.04%, 主要体现在10m以下的小尺度;由空间自相关引起的空间异质性占59.96%, 主要体现在10—70.15m范围内, 大于70.15m高原鼢鼠土丘不存在空间自相关。普通克里格插值也表明, 土丘的分布存在明显的斑块化特征(图 1)。
| 因子Factor | 模型Model | 块金值 Nugget (C0) |
基台值 Structural (C0+C) |
块金值/基台值 Proportion [C0/(C0+C)] |
变程Range | 残差RSS | 决定系数 R2 |
| 高原鼢鼠土丘 Mounds of plateau zokor |
Gaussian | 0.02138 | 0.05328 | 40.13% | 70.2 | 0.00025 | 0.858 |

|
| 图 1 高原鼢鼠土丘空间分布图 Fig. 1 Spatial distribution of mounds of plateau zokor |
由表 2可以看出, 10—20cm土壤水分、容重、地下生物量以及地上生物量、根系营养物质以及各植物功能群丰富度可分别用高斯、球状和指数模型拟合;土壤水分、粗脂肪含量和杂类草功能群丰富度存在中等强度的空间变异, C0/C0+C介于25%—75%之间, 表明其空间变异由随机性和结构性因素共同作用;其余环境因子(土壤容重、地上、地下生物量、根系粗蛋白含量、根系可溶性糖含量、禾本科及莎草科丰富度)的空间变异强度较高, C0/C0+C均小于25%, 表明空间变异主要由结构性因素引起。普通克里格插值表明, 这些环境因子均存在明显的斑块化特征(图 2)。
| 环境因子Factor | 模型 Model |
块金值 Nugget (C0) |
基台值 Structural (C0+C) |
块金值/基台值 Proportion [C0/(C0+C)] |
变程 Range |
残差 RSS |
系数 R2 |
| 土壤容重Soil bulk density | Spherical | 0.00027 | 0.0066 | 4.09% | 15.2 | 3.11×-6 | 0.187 |
| 土壤水分Soil moisture content | Exponential | 0.00213 | 0.00427 | 49.88% | 34.7 | 8.87×-8 | 0.951 |
| 地下生物量Underground biomass | Exponential | 0.122 | 1.023 | 11.93% | 21.9 | 2.57×-2 | 0.527 |
| 地上生物量Aboveground biomass | Spherical | 10 | 4191 | 0.24% | 18.2 | 4.74×5 | 0.612 |
| 根系粗脂肪Root′s crude fat | Gaussian | 0.00819 | 0.0189 | 43.33% | 75.2 | 1.84×-6 | 0.961 |
| 根系粗蛋白Root′s crude protein | Spherical | 0.001 | 1.67 | 0.06% | 10.7 | 1.10×-1 | 0.445 |
| 根系可溶性糖Root′s water-soluble sugar | Spherical | 0.5 | 78 | 0.64% | 14.1 | 6.09 | 0.349 |
| 禾本科丰富度Richness of grass | Gaussian | 0.0023 | 0.0411 | 5.60% | 13.3 | 2.49×-5 | 0.532 |
| 莎草科丰富度Richness of sedges | Exponential | 0.002 | 0.028 | 7.14% | 23.1 | 5.15×-5 | 0.345 |
| 杂类草丰富度Richness of forbs | Exponential | 0.0568 | 0.1242 | 45.73% | 38.1 | 9.59×-5 | 0.856 |

|
| 图 2 环境因子空间分布图 Fig. 2 Spatial distribution of environmental factors |
由图 3可知, 土丘的空间分布在所研究的尺度下(10.74—71.85m)与土壤水分含量大体呈现正的空间相关性, 仅在38.74—44.01m范围内为负空间相关, 说明土丘的空间分布与土壤含水量在多种尺度下表现出不同的相关性;而与土壤容重在10.74—71.85m均呈现负空间相关。

|
| 图 3 高原鼢鼠土丘数量与土壤物理性状的空间关联性 Fig. 3 The spatial relationships between the mounds and soil physical properties |
高原鼢鼠土丘分布与与植物地上生物量在10.74—30.82m以及44.01—71.85m的空间范围内为正相关, 而在30.82—44.01m的空间范围内为负相关, 总体来说, 土丘分布在所研究范围内与地上生物量呈现正的空间相关性;与植物地下生物量在10.74—71.85m的空间范围内均呈现正相关(图 4)。

|
| 图 4 高原鼢鼠土丘数量与植物生物量的空间关联性 Fig. 4 The spatial relationships between the mounds and plant biomass |
由图 5可以看出, 高原鼢鼠土丘数量与根系粗蛋白含量的空间关联性大致表现为正相关, 仅在10.74—17.37m和59.23—65.01m空间范围内呈现负相关, 其余尺度下均为正空间相关性;与根系可溶性糖含量在10.74—30.82m空间范围内为正相关, 30.82—38.47m的空间范围内为负相关, 在38.47—65.01m空间范围内又表现为负相关;与根系粗脂肪含量在10.74—44.01m空间范围内表现为正相关, 44.01—65.01m空间范围内为负相关。

|
| 图 5 高原鼢鼠土丘数量与根系营养物质的空间关联性 Fig. 5 The spatial relationships between the mounds and root′s nutrient content |
如图 6, 高原鼢鼠土丘数量与禾本科丰富度在多尺度下呈现正的或负的空间关联性, 在10.74—51.31m的空间范围内, 呈现负相关, 大于51.31m呈现正空间关联;土丘数量与莎草科丰富度在10.74—71.85m的空间范围内均呈现负空间相关性, 而与杂类草丰富度在所研究的空间尺度下(10.74—71.85m)均呈现正空间相关性。

|
| 图 6 高原鼢鼠土丘数量与植物功能群丰富度的空间关联性 Fig. 6 The spatial relationships between the mounds and richness of plant functional group |
由表 3看出, 土丘的空间分布与土壤容重、禾本科丰富度以及莎草科丰富度呈负相关, 与其他环境因子均呈现正相关。通过mantel检验结果表明, 在95%的置信水平下, 仅有莎草科和杂类草丰富度与土丘分布呈现显著的负相关或正相关(P < 0.05), 而在90%的置信水平下, 土丘分布与土壤容重以及莎草科丰富度呈显著负相关(P < 0.1), 与根系粗脂肪含量和杂类草丰富度呈显著正相关(P < 0.1)。
| 矩阵Matrix | Mantel test曼特尔检验 | ||
| r | 95%置信区间P | 90%置信区间P | |
| 鼠丘分布×土壤容重Distribution of mounds×soil bulk density | -0.45 | 0.0601 | 0.0457* |
| 鼠丘分布×土壤水分Distribution of mounds×soil moisture content | -0.089 | 0.068 | 0.053 |
| 鼠丘分布×地上生物量Distribution of mounds×aboveground biomass | -0.3 | 0.079 | 0.0617 |
| 鼠丘分布×地下生物量Distribution of mounds×underground biomass | 0.636 | 0.0731 | 0.0564 |
| 鼠丘分布×根系粗脂肪Distribution of mounds×Root′s crude fat | 0.175 | 0.0589 | 0.044* |
| 鼠丘分布×根系可溶性糖Distribution of mounds×Root′s water-soluble sugar content | 0.6 | 0.0879 | 0.065 |
| 鼠丘分布×根系粗蛋白Distribution of mounds×Root′s crude protein | 0.54 | 0.0671 | 0.0526 |
| 鼠丘分布×禾本科丰富度Distribution of mounds×richness of grass | -0.07 | 0.0658 | 0.098 |
| 鼠丘分布×莎草科丰富度Distribution of mounds×richness of sedges | -0.302 | 0.0422** | 0.0347* |
| 鼠丘分布×杂类草丰富度Distribution of mounds×richness of forbs | 0.133 | 0.0465** | 0.0336* |
| **表示在95%置信水平下有显著性差异, *表示在90%置信水平下有显著性差异 | |||
国内对啮齿动物种群空间分布研究开展较早并取得多项成果[7]。但对于高原鼢鼠的种群空间分布格局及其与环境因子的关系的研究鲜见报道。本试验采用地统计分析方法重点探讨高原鼢鼠种群空间分布格局及其与环境因子的空间关联性, 研究发现高原鼢鼠种群表现出明显的异质性特征, 各环境因子也存在不同程度的空间变异, mantel检验表明高原鼢鼠种群主要分布在土壤较为疏松、莎草科丰富度较低、杂类草丰富度及根系粗脂肪含量较高的栖息地。
啮齿动物空间分布的研究方法大多采用频次比较法和分布型指标法判断其分布型, 或是利用经典统计学方法分析非生物因素对分布格局的影响[7]。本研究基于小尺度(100m×140m)来探讨高原鼢鼠种群空间分布格局及其与环境因子的关系, 消除了由于地理差异而导致的环境对于高原鼢鼠空间分布的影响。本研究采用地统计学的分析方法是因为啮齿动物的分布是复杂的并且在不同的空间尺度具有结构性特征, 同时生态系统生态学过程在不同的空间下可能会表现出不同的空间分布格局, 而地统计分析方法恰好可以定量分析各变量在不同尺度下的空间关系[28]。因此, 在研究高原鼢鼠土丘分布的空间异质性以及与不同环境因子的空间关系中采用地统计学方法, 合理描述土丘分布与各环境因子分布在不同尺度下的空间关系。
高寒草甸生态系统的各组分在空间上具有格局和缀块状的分布, 这种空间异质性也是生态系统中比较普遍的现象[29]。本研究采用半方差函数分析结果表明, 高原鼢鼠土丘分布存在中等程度的空间变异, 这种变异主要由随机性因素和结构性因素共同调控, 随机性因素可能由于试验误差引起, 或是由于小于10m的空间过程也存在变异;结构性因素是由于非生物因素或是生物因素引起。通过计算土丘扩散系数以及对土丘进行普通克里格制图发现, 高原鼢鼠土丘呈现聚集分布现象。该研究结果与陈林等[13]和宗浩等[10]的研究结果相一致。同时, 与高原鼢鼠分布密切相关的环境因子也存在不同程度的空间变异, 其空间格局可分别用高斯、球状、指数模型进行拟合。块金值和基台值的比值(C0/C0+C)表明这些环境因子的空间异质性均表现为明显的结构性特征, 土壤容重、地上、地下生物量、根系蛋白质含量、根系可溶性糖含量、禾本科丰富度以及莎草科丰富度的空间分异主要受结构型因素调控。
交方差函数分析结果表明, 高原鼢鼠土丘分布与土壤容重、莎草科丰富度以及杂类草丰富度在所研究的尺度下均完全表现为负的或正空间关联性;而与土壤水分、地上生物量、根系营养物质和禾本科丰富度在多种尺度下表现出复杂的正的或负的空间关联性, 说明高原鼢鼠土丘的分布与这些环境因子并不总是呈现正的或负的相关性, 而是在特定的尺度下呈现出特定的相关性, 大于该尺度或小于该尺度相关性都会发生变化。通过mantel检验发现, 在90%的置信水平下高原鼢鼠土丘与土壤容重和莎草科丰富度存在显著的负空间关联性(P < 0.1), 与根系粗脂肪含量和杂类草丰富度存在显著的正的空间关联性(P < 0.1), 而在95%的置信水平下, 土丘分布仅与杂类草丰富度和莎草科丰富度存在显著的正的或负的空间关联性(P < 0.05)。其他环境因子与高原鼢鼠土丘分布存在伪(spurious)空间关联性, 这种空间关联性可能由简单的空间格局形成, 并不是正真的空间关联性。非生物因子被认为是影响高原鼢鼠分布的重要因子。在土壤物理性质方面, 土壤容重被认为是影响高原鼢鼠栖息地选择的主要因素之一[30]。而本研究发现在90%的置信水平下高原鼢鼠土丘分布与土壤容重具有显著的负空间关联性(P < 0.1), 这与刘丽[31]的研究结果相一致。有研究发现, 高原鼢鼠挖掘一个单位长度所消耗的能量是相同体重地上鼠行走相同长度所消耗能量360—3400倍[32], 因此高原鼢鼠为避免大量的能量消耗, 选择土壤较为疏松的地方作为其日常活动场所。在根系营养方面, 土丘分布仅与根系粗脂肪含量存在显著的正空间关联性(P < 0.1), 可能由于高原鼢鼠在越冬前期要储存大量食物, 而较高的脂肪摄入会保证其安全过冬。此外, 本试验在8月份进行采样恰好是高原鼢鼠为了储存越冬食物而向地面推土丘的第二个高峰前期。在植物组成方面, 土丘分布与杂类草丰富度存在显著的正空间关联性(P < 0.05), 而与莎草科丰富度呈现显著的负空间关联性(P < 0.05), 说明高原鼢鼠多分布于杂类草较多的地方而莎草科较少的地方, 因为杂类草是高原鼢鼠的主要食物来源。此研究结果与楚彬等[30]对高原鼢鼠栖息地选择分析研究的结果相一致, 同时Wang等[33]也认为杂类草较多的地方是高原鼢鼠较为偏好的。而莎草科并不是高原鼢鼠较为喜食的植物, 王权业等[34]对高原鼢鼠的食性研究发现在其胃容物内莎草科植物仅有两种。
综上所述, 在本研究的尺度下(100m×140m)引起高原鼢鼠分布的主要非生物因素是土壤疏松程度、根系粗脂肪含量以及杂类草和莎草科丰富度。动物栖息地利用的描述取决于空间尺度, 某一尺度上的研究结果不能用于推断其他尺度上的状态[35]。因此本研究结果也不能否定其他环境因子与土丘分布的空间关系, 这种现象恰恰说明在不同尺度条件下考察高原鼢分布与环境因子的研究有很大的必要性。同时, 生物因子(种内作用)对高原鼢鼠分布的影响也是不能被忽视的重要因素。
致谢: 感谢王巧玲, 刘丽, 王贵珍, 在野外调查和样品化验过程中的帮助。| [1] | 赵天飙. 大沙鼠种群空间分布格局、栖息地选择及种群动态的研究[D]. 呼和浩特: 内蒙古大学, 2006, http://cdmd.cnki.com.cn/Article/CDMD-10126-2006126332.htm |
| [2] | 高福元, 赵成章, 卓马兰草. 高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性. 生态学报, 2014, 34(3): 605–612. |
| [3] | 申跃武, 廖文波, 胡锦矗. 南充市微尾鼩种群空间分布的研究. 西华师范大学学报:自然科学版, 2005, 26(2): 149–151. |
| [4] | Davidson A D, Lightfoot D C. Burrowing rodents increase landscape heterogeneity in a desert grassland. Journal of Arid Environments, 2008, 72(7): 1133–1145. DOI:10.1016/j.jaridenv.2007.12.015 |
| [5] | 孙儒泳. 动物生态学原理. 北京: 北京师范大学出版社, 2001: 161–164. |
| [6] | Gutiérrez-López M, Jesús J B, Trigo D, Fernández R, Novo M, Díaz-Cosín D J. Relationships among spatial distribution of soil microarthropods, earthworm species and soil properties. Pedobiologia, 2010, 53(6): 381–389. DOI:10.1016/j.pedobi.2010.07.003 |
| [7] | 房继明. 啮齿动物的空间分布格局. 生态学杂志, 1994, 13(1): 39–44. |
| [8] | 赵天彪, 李新民, 张忠兵, 张春福, 齐林. 大沙鼠和子午沙鼠种群空间分布格局的研究. 兽类学报, 1998, 18(2): 131–136. |
| [9] | 房继明, 孙儒泳. 布氏田鼠空间分布格局的季节动态. 生态学报, 1991, 11(2): 111–116. |
| [10] | 宗浩, 樊乃昌, 于福溪, 朱嘉城. 高寒草甸生态系统优势鼠种高原鼢鼠(Myospalax baileyi)和高原鼠兔(Ochotona curzoniae)种群空间格局的研究. 生态学报, 1991, 11(2): 125–129. |
| [11] | 樊乃昌, 王权业, 周文扬, 景曾春. 高原鼢鼠种群数量与植被破坏程度的关系//高寒草甸生态系统国际学术讨论会文集. 北京: 科学出版社, 1988: 109-115. |
| [12] | 张堰铭, 刘季科. 地下鼠生物学特征及其在生态系统中的作用. 兽类学报, 2002, 22(5): 145–155. |
| [13] | 何俊龄, 张金沙, 杨莹博, 邓汉刚, 魏进招, 张卫国. 高原鼢鼠土丘空间格局及主要特征研究. 草业学报, 2006, 15(1): 107–112. |
| [14] | 陈林, 廖文波, 杨志松, 张阳, 何娅, 周材权. 高原鼢鼠土丘空间分布研究. 西北师范大学学报:自然科学版, 2010, 31(2): 122–125. |
| [15] | 李凤霞, 张德罡. 草地退化指标及恢复措施. 草原与草坪, 2005(1): 24–28. |
| [16] | 汪志刚, 刘荣堂, 陈艳宇, 如甲. 高原鼢鼠繁殖指数探讨. 草业学报, 1995, 4(1): 61–68. |
| [17] | 周延山, 花立民, 楚彬, 刘丽, 姬程鹏, 田永亮. 祁连山东段高原鼢鼠对高寒草甸危害评价. 生态学报, 2016, 36(18): 5922–5930. |
| [18] | 吕贻忠, 李保国. 土壤学. 北京: 中国农业出版社出版, 2006. |
| [19] | 李帅, 花立民, 聂中南, 郑阳. 草地干重排序快速监测方法在高寒草甸中的应用. 生态学杂志, 2015, 34(12): 3575–3580. |
| [20] | 杨柳, 王建立, 王淑英, 路苹. 糖类物质测定方法评价. 北京农学院学报, 2009, 24(4): 68–71. |
| [21] | 鲁如坤. 土壤农业化学分. 北京: 中国农业科技出版社, 2000: 263–264. |
| [22] | Maxwel B, Wiatr S M, Fay P K. Energy potential of leafy spurge (Euphorbia esula). Economic Botany, 1985, 39(2): 150–156. DOI:10.1007/BF02907837 |
| [23] | 王政权. 地统计学及在生态学中的应用. 北京: 科学出版社, 1999. |
| [24] | Cambardella C A, Moorman T B, Parkin T B, Karlen D L, Novak J M, Turco R F, Konopka A E. Field-scale variability of soil properties in central Iowa soils. Soil Science Society of America Journal, 1994, 58(5): 1501–1511. DOI:10.2136/sssaj1994.03615995005800050033x |
| [25] | 郭旭东, 傅伯杰, 马克明, 陈利顶, 杨福林. 基于GIS和地统计学的土壤养分空间变异特征研究——以河北省遵化市为例. 应用生态学报, 2000, 11(4): 557–563. |
| [26] | Rossi J P, Lavelle P, Tondoh J E. Statistical tool for soil biology X. Geostatistical analysis. European Journal of Soil Biology, 1995, 31(4): 173–181. |
| [27] | 王正军, 李典谟, 商晗武, 程家安. 地质统计学理论与方法及其在昆虫生态学中的应用. 昆虫知识, 2002, 39(6): 405–411. DOI:10.7679/j.issn.2095-1353.2002.116 |
| [28] | 郑祥, 鲍毅新, 葛宝明. GIS在野生动物空间分布格局研究中的应用. 四川动物, 2003, 22(4): 277–280. |
| [29] | 陈玉福, 董鸣. 生态学系统的空间异质性. 生态学报, 2003, 23(2): 346–352. |
| [30] | 楚彬, 花立民, 周延山, 刘丽, 李帅. 祁连山东段不同放牧强度下高原鼢鼠栖息地选择分析. 草业学报, 2016, 25(1): 179–186. DOI:10.11686/cyxb2015234 |
| [31] | 刘丽, 花立民, 杨思维, 周建伟, 王巧玲, 王贵珍, 楚彬, 周延山. 放牧干扰下高原鼢鼠栖息地选择因素. 动物学杂志, 2015, 50(5): 725–734. |
| [32] | Vleck D. The energy cost of burrowing by the pocket gopher Thomomys bottae. Physiological Zoology, 1979, 52(2): 122–136. DOI:10.1086/physzool.52.2.30152558 |
| [33] | Wang Q Y, Zhou W Y, Wei W H, Zhang Y M, Fan N C. The burrowing behavior myospalax bailey and its relation to soil hardness. Acta Theriologica Sinica, 2000, 20(4): 277–283. |
| [34] | 王权业, 张堰铭, 魏万红, 边疆晖. 高原鼢鼠食性的研究. 兽类学报, 2000, 20(3): 193–199. |
| [35] | 杨维康, 钟文勤, 高行宜. 鸟类栖息地选择研究进展. 干旱区研究, 2000, 17(3): 71–78. |