 2018, Vol. 38
2018, Vol. 38文章信息
- 苏培玺, 周紫鹃, 侍瑞, 解婷婷.
- SU Peixi, ZHOU Zijuan, SHI Rui, XIE Tingting.
- 高寒草毡层基本属性与固碳能力沿水分和海拔梯度的变化
- Variation in basic properties and carbon sequestration capacity of an alpine sod layer along moisture and elevation gradients
- 生态学报. 2018, 38(3): 1040-1052
- Acta Ecologica Sinica. 2018, 38(3): 1040-1052
- http://dx.doi.org/10.5846/stxb201611222384
-
文章历史
- 收稿日期: 2016-11-12
- 网络出版日期: 2017-10-18





2. 中国科学院寒旱区陆面过程与气候变化重点实验室, 兰州 730000
2. Key Laboratory of Land Surface Process and Climate Change in Cold and Arid Regions, Chinese Academy of Sciences, Lanzhou 730000, China
青藏高原是全球海拔最高的一个巨型构造地貌单元, 具有独特的自然环境和自然分异规律[1]。位于青藏高原东部的若尔盖高原, 是黄河和长江上游重要的水源涵养地, 分布有沼泽、草甸、灌丛草甸、灌丛等高寒地区典型的植被群落类型。高寒草毡层是高原寒区自然植被下形成的草毡状耐搬运性表土层, 松软及坚韧程度因群落类型和发育程度而不同, 是高寒植被繁殖更新的芽库, 是高寒生态系统物质循环和能量流动的枢纽。草毡层是高寒区植被下广泛分布的特殊土壤表层部分, 也就是根系交织盘结极为紧密层, 又称草毡土。由于研究区域和研究侧重点的不同, 不同研究者用不同的术语, 如草毡表层、植毡层、草根层、草结皮层、寒毡土、寒冻毡土等[2-5], 都是不同的表述或其中一部分。草毡层韧性极大, 铁铲也不易切割。草毡层有机物质按分解程度可分为3部分, 即交织盘结的活根和未分解或分解程度极低, 基本保持原有外形和韧性的草毡状有机土壤物质;丧失原有形态和韧性, 处于不同分解阶段的半分解有机土壤物质;分解程度很高或由微生物合成的腐殖物质[3];简单地可分为活根、死根和腐殖质三部分。草毡层碳储量包括地下生物量碳储量和土壤碳储量两部分。认识草毡层土壤有机碳(SOC)含量及其变异规律是认识草毡层生态功能的基础。
土壤是陆地表面碳的最大储存库, 土壤碳库储存的有机碳在1500—2000Pg C (1Pg=1015g)[6-7], 植被碳库生物量碳大约为500Pg C, 大气碳库大约在785Pg C[7];土壤碳库是植被碳库的3—4倍, 大气碳库的2—2.5倍, 是全球碳循环中最为重要的组成部分。SOC库的微小变化都会引起大气中CO2浓度的极大变化[7]。由于不合理的土地利用和化石碳的排放, 大气CO2每年增加3Pg C, 到21世纪末, CO2浓度将达到工业革命前的2倍以上[7]。SOC是土壤碳库的主要组成部分, 也是大气CO2的主要源[8], 在陆地生态系统与大气交换的CO2中, 土壤有机质分解释放的CO2大约占2/3[9]。表层土壤有机碳储量对于理解不同植被类型土壤碳动态和土壤固碳潜力至关重要[10]。
土壤有机碳库的大小在不同生态区域变化很大, 寒冷和湿润区域要大于温暖和干燥区域[11];其动态变化可对生态系统主要功能、组成和发展方向产生显著影响[12]。在全球变暖的背景下, 储存在高寒区域的SOC较温暖区域的SOC更容易丢失[13]。在青藏高原高寒草地生态系统, 土壤有机碳主要分布在土壤表层[14]。高寒草毡层这种特殊的表土层, 其固碳能力反映着高寒生态系统的固碳能力, 其易于剥蚀和退化给草地保护起到警示作用, 其耐搬运性给工程施工的异地主动利用提供了可能。草毡层沿水分与海拔梯度的物理和化学属性, 有机碳密度和碳储量的变化规律, 土壤水分有效性与固碳能力的关系, 保持草毡层稳定的土壤水分阈值等, 这些问题的回答, 可为正确处理高寒草地生产功能与碳储生态功能的关系, 理解高寒植被生态过程对气候变化的响应, 减少不合理利用导致碳排放、增加土壤碳储量、延长土壤碳驻留时间等提供重要资料和理论依据。
1 材料与方法 1.1 研究区概况研究区位于青藏高原东部若尔盖高原, 地处四川、甘肃和青海三省结合部, 行政区划上包括若尔盖、红原、阿坝、碌曲、玛曲和久治6县域, 是大、小牲畜的主要牧场。林线是亚高山和高山区划分的主要依据[15-16], 亚高山带是指山地带的上限到森林界线的部分, 山地带分为浅山区(2100≤海拔(H)<2500m)和中山区(2500≤H<3300m)。若尔盖县日尔郎山紫果云杉(Picea purpurea)分布最高海拔为3674m, 紫果云杉分布在海拔2600—3800m[17], 以3800m为界将若尔盖高原划分为亚高山区(3300≤H<3800m)和高山区(H≥3800m)。
若尔盖高原又称若尔盖大草原, 为草甸草原, 以中生、湿生和水生等多年生草本植物为主, 主要由沼泽和草甸组成。沼泽以湿生植物和水生植物为主, 湿草甸以湿中生植物和湿生植物为主, 干草甸以旱中生植物和中生植物为主。草甸草原不同于分布在半干旱、半湿润气候区的典型草原和分布于干旱荒漠区的荒漠草原, 荒漠草原以旱生植物和中旱生植物为主。若尔盖高原植被在水平地带根据地表积水水位和时间长短及植被状况划分为, 沼泽、退化沼泽、沼泽化草甸、湿草甸、干草甸和退化草甸。在垂直地带根据海拔高度不同划分为, 亚高山草甸、亚高山灌丛草甸、高山灌丛草甸、高山草甸。湿草甸优势种为莎草科植物, 非禾草[18], 也叫真草甸或莎草草甸;地势缓平低洼, 有短期积水, 排水不畅, 也叫低洼草甸。干草甸优势种为禾本科植物, 也叫草原化草甸或禾草草甸;地势倾斜高亢, 无积水, 也叫高亢草甸。通常人们将青藏高原湿草甸和干草甸区分为草甸和草原。
2013年, 对若尔盖高原进行了全面考察, 根据典型性和代表性, 在33°51′—33°58′N, 102°08′—102°53′E的甘肃玛曲县河曲马场和四川若尔盖县花湖一带, 同一海拔高度(3440±4)m范围内, 布设了水分梯度固定样地;在34°03′—34°04′N, 102°10′—102°11′E的玛曲县尕玛梁山区, 海拔每升高100m布设了海拔梯度固定样地。2015年8月末到9月初, 植物生长后期, 对水分和海拔梯度植物群落和草毡层厚度进行了调查(表 1), 对草毡层及以下土壤进行了采样。
| 类别 Category |
群落类型 Community type |
海拔 Elevation/m |
草毡层厚度 Thickness of sod layer/cm |
总盖度 Total coverage/% |
植物种类组成Plant species composition | |
| 优势种 Dominant species |
常见种 Common species |
|||||
| 水分梯度 Moisture gradient |
沼泽 | 3440 | 30 | 80 | 木里苔草(Carex muliensis)、四川嵩草(Kobresia setchwanensis) | 华扁穗草(Blysmus sinocompressus)、双柱头藨草(Scirpus distigmaticus)、发草(Deschampsia caespitosa)、溚草(Koeleria cristata)、线叶龙胆(Gentiana farreri)、肋柱花(Lomatogonium carinthiacum)等 |
| 退化沼泽 | 3440 | 32 | 85 | 华扁穗草(B. sinocompressus)、四川嵩草(K. setchwanensis) | 木里苔草(C. muliensis)、青藏苔草(Carex moorcroftii)、发草(D. caespitosa)、双柱头藨草(S. distigmaticus)、香芸火绒草(Leontopodium haplophylloides)、鹅绒委陵菜(Potentilla anserina)等 | |
| 沼泽化草甸 | 3440 | 35 | 90 | 华扁穗草(B. sinocompressus)、四川嵩草(K. setchwanensis) | 木里苔草(C. muliensis)、发草(D. caespitosa)、双柱头藨草(S. distigmaticus)、甘肃马先蒿(Pedicularis kansuensis)、线叶龙胆(G. farreri)、鹅绒委陵菜(P. anserina)等 | |
| 湿草甸 | 3440 | 33 | 95 | 四川嵩草(K. setchwanensis)、垂穗披碱草(Elymus nutans) | 青藏苔草(C. moorcroftii)、双柱头藨草(S. distigmaticus)、垂穗鹅观草(Roegneria nutans)、溚草(K. cristata)、草地早熟禾(Poa pratensis)、鹅绒委陵菜(P. anserina)等 | |
| 干草甸 | 3440 | 31 | 90 | 垂穗披碱草(E. nutans)、四川嵩草(K. setchwanensis) | 垂穗鹅观草(R. nutans)、赖草(Leymus secalinus)、草地早熟禾(P. pratensis)、青藏苔草(C. moorcroftii)、甘肃马先蒿(P. kansuensis)、鹅绒委陵菜(P. anserina)等 | |
| 退化草甸 | 3440 | 30 | 65 | 垂穗披碱草(E. nutans)、垂穗鹅观草(R. nutans) | 赖草(L. secalinus)、草地早熟禾(P. pratensis)、四川嵩草(K. setchwanensis)、鹅绒委陵菜(P. anserina)、甘肃马先蒿(P. kansuensis Maxim.)、大籽蒿(Artemisia sieversiana)等 | |
| 海拔梯度 Elevation gradient |
亚高山草甸 | 3550 | 30 | >95 | 垂穗披碱草(E. nutans)、矮生嵩草(K. humilis) | 垂穗鹅观草(R. nutans)、双叉细柄茅(Ptilagrostis dichotoma)、高山嵩草(K. pygmaea)、珠芽蓼(Polygonum viviparum)、鹅绒委陵菜(P. anserina)、香芸火绒草(L. haplophylloides)等 |
| 亚高山草甸 | 3650 | 29 | >95 | 矮生嵩草(K. humilis)、高山嵩草(K. pygmaea) | 垂穗披碱草(E. nutans)、双柱头藨草(S. distigmaticus)、金露梅(Potentilla fruticosa)、线叶嵩草(Kobresia capillifolia)、青藏苔草(C. moorcroftii)、细柄茅(Ptilagrostis mongholica)等 | |
| 亚高山灌丛草甸 | 3750 | 30 | >95 | 金露梅(P. fruticosa)、垂穗披碱草(E. nutans) | 矮生嵩草(K. humilis)、珠芽蓼(P. viviparum)、双柱头藨草(S. distigmaticus)、垂穗鹅观草(R. nutans)、鬼箭锦鸡儿(Caragana jubata)、长毛风毛菊(Saussurea hieracioides)等 | |
| 高山灌丛草甸 | 3850 | 28 | >95 | 鬼箭锦鸡儿(C. jubata)、异针茅(Stipa aliena) | 垂穗披碱草(E. nutans)、金露梅(P. fruticosa)、矮生嵩草(K. humilis)、青藏苔草(C. moorcroftii)、珠芽蓼(P. viviparum)、双叉细柄茅(P. dichotoma)等 | |
| 高山灌丛草甸 | 3950 | 26 | >95 | 鬼箭锦鸡儿(C. jubata)、高山嵩草(K. pygmaea) | 甘肃嵩草(Kobresia kansuensis)、青藏苔草(C. moorcroftii)、垂穗披碱草(E. nutans)、珠芽蓼(P. viviparum)、紫羊茅(Festuca rubra)、双叉细柄茅(P. dichotoma)等 | |
| 高山草甸 | 4050 | 25 | >95 | 高山嵩草(K. pygmaea)、甘肃嵩草(K. kansuensis) | 青藏苔草(C. moorcroftii)、乳白香青(Anaphalis lactea)、大花肋柱花(Lomatogonium macranthum)、圆穗蓼(Polygonum macrophyllum)、线叶龙胆(G. farreri)、钝裂银莲花(Anemone obtusiloba)等 | |
植物名称由兰州大学和西北师范大学的植物分类专家现场确认, 对不太确定的植物种类采集生殖器官、拍摄图片, 查阅相关资料确定, 对一些中文名易混淆的植物, 查阅《中国植物志》全文电子版进行统一, 如小嵩草(Kobresia pygmaea), 叫高山嵩草。
1.2 草毡层基本属性测定不同群落草毡层厚度不同, 平均在(30±3)cm之间(表 1), 为便于比较, 统一分为上(0—10cm)、中(10—20cm)、下(20—30cm)3层。海拔梯度植被草毡层下为岩石碎块, 质地坚硬。水分梯度群落增加了30—40、40—60、60—80、80—100cm土层的土壤采样。各层采集土壤样品1kg带回实验室, 供室内测定分析物理和化学属性。
容重:采用环刀法测定。草毡层取样困难, 在野外采用特制的环刀取样器取样, 即在环刀外面加一个不锈钢制环刀外套, 锋利口长出环刀口1cm, 这样压入草毡层, 套在里面的环刀就容易取到原状样。将环刀样称鲜重后装入铝盒, 放入烘箱中, 在105℃下烘干8h以上至恒重, 称干重, 计算单位容积的烘干重量, 即为容重。重复3—5次。
土壤颗粒组成:颗粒组成按粒径大小分为石砾(>2mm)、沙粒(0.05—2.0mm)、粉粒(0.002—0.05mm)和粘粒(<0.002mm)。将分析样品过2mm筛, 得出石砾含量。沙粒、粉粒和粘粒运用湿筛+吸管法测定。
全碳(C)、全养分和速效养分含量:全C、氮(N)用vario MACRO cube元素分析仪(Elementar, Germany), 全磷(P)用硫酸-高氯酸-钼锑抗比色法, 全钾(K)采用氢氟酸-高氯酸消煮法测定。速效N采用碱解扩散法, 速效P采用碳酸氢钠浸提-钼锑抗比色法, 速效K采用乙酸铵浸提-火焰光度计法测定。
有机C含量:采用重铬酸钾氧化-外加热法测定。在外加热条件下, 用一定浓度的重铬酸钾-硫酸溶液氧化土壤样品有机碳, 剩余的重铬酸钾量用硫酸亚铁溶液滴定, 根据重铬酸钾的消耗量计算有机碳含量。全碳含量减去有机碳含量为无机碳含量。
1.3 草毡层土壤有机碳密度和储量计算土壤有机碳密度通常是指单位面积单位深度土体中土壤有机碳质量, 国际上通常是以1m深度来计算, 单位为kg C/m2;土壤有机碳储量是指区域范围内1m深度的土壤有机碳总质量[7]。草毡层计算深度为30cm。
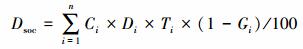
|
式中, Dsoc为土壤有机碳密度(kg C/m2), Ci为第i层土壤有机碳含量(g/kg), Di为第i层土壤容重(g/cm3), Ti为第i层土层厚度(cm), Gi为第i层土壤中直径>2mm的石砾含量(g/g)。n为土层数目, i为第i土层。
土壤有机碳储量计算公式为:
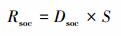
|
式中, Rsoc为土壤有机碳储量, S为面积。
1.4 统计分析对测定数据采用Excel统计分析, 各指标差异显著性利用SPSS 16.0软件, 进行单因素方差分析(One-Way ANOVA), 不同群落之间的差异显著性采用邓肯多重比较检验(Duncan′s multiple range test), 作图用Origin 8.6软件。
2 结果与分析 2.1 草毡层基本理化性质变化 2.1.1 草毡层物理性质水分梯度不同群落草毡层容重差异极显著(P<0.01), 沼泽湿地最低, 只有0.26g/cm3;退化草甸最高, 达到1.17g/cm3 (图 1)。海拔梯度群落之间草毡层容重变化不大, 差异不显著(图 1), 平均为0.94g/cm3, 与干草甸相近, 低于退化草甸。

|
| 图 1 水分和海拔梯度群落草毡层容重变化 Fig. 1 Bulk density change of sod layer in communities along the moisture and elevation gradients SW:沼泽, Swamp;DS:退化沼泽, Degraded swamp;SM:沼泽化草甸, Swampy meadow;WM:湿草甸, Wet meadow;UM:高亢草甸, Upland meadow, 也叫干草甸, Dry meadow;DM:退化草甸, Degraded meadow;SM1:海拔3550m亚高山草甸, Subalpine meadow at elevation of 3550m;SM2:海拔3650m亚高山草甸, Subalpine meadow at elevation of 3650m;SSM:海拔3750m亚高山灌丛草甸, Subalpine shrub meadow at elevation of 3750m;ASM1:海拔3850m高山灌丛草甸, Alpine shrub meadow at elevation of 3850m;ASM2:海拔3950m高山灌丛草甸, Alpine shrub meadow at elevation of 3950m;AM:海拔4050m高山草甸, Alpine meadow at elevation of 4050m;图中相同的小写字母表示不同群落之间差异不显著, 不同的大写字母表示不同群落之间差异极显著(P<0.01) |
含水量在水分梯度不同群落草毡层之间差异极显著, 沼泽和草甸之间存在极显著差异(P<0.01);草甸之间比较, 沼泽化草甸与干草甸和退化草甸之间存在显著差异(P<0.05) (图 2), 干草甸和退化草甸质量含水量分别为32.7%和25.9%。海拔梯度不同群落之间含水量比较, 高山区高于亚高山区, 含水量平均分别为56.5%和48.4%;但总体比较差异不显著(图 2), 平均为52.5%。

|
| 图 2 水分和海拔梯度群落生长后期草毡层含水量变化 Fig. 2 Moisture content change of sod layer in the late growth stage in communities along the moisture and elevation gradients 图中不同的小写字母表示不同群落之间差异显著(P<0.05), 不同的大写字母表示不同群落之间差异极显著(P<0.01) |
土壤机械组成分析表明, 水分梯度各群落草毡层粘粒含量>10%, 粉粒含量>15%, 沙粒含量>35%(图 3);地势低洼的沼泽、退化沼泽和沼泽化草甸石砾含量高, 在10%以上。从图 3可以看出, 随着海拔升高, 粘粒含量有降低的趋势, 亚高山平均为18.7%, 高山平均为16.2%;粉粒含量有增加的趋势, 亚高山平均为42.6%, 高山平均为49.6%。亚高山和高山沙粒含量平均为36.5%。

|
| 图 3 水分和海拔梯度群落草毡层土壤颗粒组成变化 Fig. 3 Soil particle composition of sod layer in communities along the moisture and elevation gradients |
pH值在水分梯度不同群落草毡层之间存在极显著差异(P<0.01);沼泽湿地最高, 为8.2, 与不同草甸之间存在极显著差异(P<0.01);沼泽化草甸、湿草甸、干草甸和退化草甸平均为7.5。pH值在海拔梯度不同群落草毡层之间不存在显著差异, 平均值为7.5, 变幅很小, 与水分梯度不同草甸平均值一致。
总含盐量在水分梯度不同群落草毡层之间存在极显著差异(P<0.01), 沼泽最高, 为3.1g/kg, 与不同草甸之间存在极显著差异;干草甸最低, 为0.5g/kg, 退化草甸开始有所升高;沼泽化草甸为1.9g/kg, 介于沼泽和其他草甸之间。总含盐量在海拔梯度不同群落草毡层之间不存在显著差异, 各群落草毡层总含盐量<1g/kg。
土壤养分含量的变化直接反映土壤质量和土地生产力的变化。水分梯度群落草毡层土壤全养分分析表明(图 4), 全N含量存在极显著差异, 沼泽、退化沼泽和沼泽化草甸显著大于湿草甸、干草甸和退化草甸(P<0.01), 前者平均为21.2g/kg, 后者平均为5.5g/kg;草甸全N含量的变化趋势很明显, 随着土壤水分有效性的降低和草甸退化, 全N含量明显降低。全P含量不同群落之间亦存在极显著差异, 沼泽、退化沼泽和沼泽化草甸显著大于湿草甸、干草甸和退化草甸, 前者平均为2.7g/kg, 后者平均为1.7g/kg。全K含量不同群落之间同样差异极显著, 草甸极显著高于沼泽, 湿草甸、干草甸和退化草甸平均为18.0g/kg, 沼泽和退化沼泽平均只有6.3g/kg。

|
| 图 4 水分和垂直梯度群落草毡层土壤养分含量变化 Fig. 4 Soil nutrient content of sod layer in communities along the moisture and elevation gradients 图中同一指标不同的小写字母表示不同群落之间差异显著(P<0.05), 不同的大写字母表示不同群落之间差异极显著(P<0.01) |
海拔梯度不同群落草毡层土壤全N含量差异不显著(图 4), 平均为6.0g/kg, 高于干草甸和退化草甸。全P、K含量存在显著差异(P<0.05), 全P平均为2.0g/kg, 低于沼泽湿地, 高于退化草甸;全K总体为亚高山低于高山, 平均值分别为20.7g/kg和22.6g/kg。
速效养分分析表明, 水分梯度不同群落草毡层土壤速效N存在极显著差异, 沼泽、退化沼泽和沼泽化草甸极显著高于湿草甸、干草甸和退化草甸(P<0.01, 图 4)。速效P含量沼泽和退化沼泽与湿草甸、干草甸和退化草甸之间存在极显著差异, 沼泽和退化沼泽平均为34.2mg/kg, 湿草甸、干草甸和退化草甸平均为9.0mg/kg。速效K含量沼泽、退化沼泽和沼泽化草甸极显著低于湿草甸、干草甸和退化草甸, 前者平均为123.3mg/kg, 后者平均为372.2mg/kg。
海拔梯度不同群落草毡层土壤速效N、P、K含量差异均不显著(图 4), 速效N含量随着海拔升高有增加的趋势, 平均为507.4mg/kg;速效P含量平均为12.5mg/kg;速效K含量随着海拔升高有降低的趋势, 平均为230.0mg/kg。
2.2 草毡层有机碳含量变化 2.2.1 草毡层土壤有机碳含量草毡层土壤无机碳含量在水分梯度各群落之间无显著差异, 沼泽、退化沼泽、沼泽化草甸、湿草甸、干草甸和退化草甸草毡层分别为15.2、6.7、6.8、10.2、4.1、4.5g/kg, 沼泽最高, 平均为7.9g/kg。同样, 在海拔梯度上各群落亦无显著差异, 亚高山草甸、亚高山灌丛草甸、高山灌丛草甸和高山草甸分别为4.4、4.9、5.4、3.7g/kg, 平均为4.6g/kg。总体可见, 土壤无机碳含量低洼地要大于高亢地, 沼泽湿地最高。
土壤有机碳(SOC)含量在水分梯度不同群落草毡层之间存在极显著差异(P<0.01), 沼泽显著高于草甸, 沼泽SOC含量在300g/kg以上, 退化草甸只有30g/kg(图 5)。海拔梯度不同群落SOC之间不存在显著差异(图 5), 平均为67.5g/kg, 高于干草甸和退化草甸。

|
| 图 5 水分和海拔梯度群落草毡层土壤有机碳含量变化 Fig. 5 Soil organic carbon content of sod layer in communities along the moisture and elevation gradients 图中相同的小写字母表示不同群落之间差异不显著, 不同的大写字母表示不同群落之间差异极显著(P<0.01) |
分析表明, 在水分梯度上, 沼泽、退化沼泽、沼泽化草甸、湿草甸、干草甸和退化草甸草毡层无机碳含量占有机碳含量的比例分别为4.9%、2.2%、3.0%、9.3%、9.2%和14.7%, 平均为7.2%;在海拔梯度上亚高山草甸、亚高山灌丛草甸、高山灌丛草甸和高山草甸其比例分别为7.1%、6.8%、7.7%和5.4%, 平均为6.8%。无疑, 提高SOC含量是增强草毡层固碳能力的根本。
2.2.2 水分梯度群落1m深度土壤有机碳含量在水分梯度上, 不同群落SOC含量在1m深土层中差别很大(图 6), 总体上看, 从沼泽→退化沼泽→沼泽化草甸→湿草甸→干草甸→退化草甸的水分递减梯度中, 有机碳含量亦呈递减趋势;不同群落随着土壤深度的增加, 有机碳含量呈明显下降趋势, 0.3—1m土层SOC含量平均分别为267.7、260.5、182.0、59.6、13.2、9.1g/kg, 分别为草毡层的86%、86%、80%、55%、30%和30%。可见, 高寒地区土壤有机碳主要聚集在草毡层, 随着土壤水分减少, 这种趋势越明显。

|
| 图 6 水分梯度群落1m深度土壤有机碳含量变化 Fig. 6 Soil organic carbon content in the 1m depth soil layer in communities along the moisture gradient |
草毡层SOC密度和储量变化见图 7, 水分梯度上沼泽和退化沼泽草毡层SOC密度和储量平均分别为20.1kg C/m2和201t C/hm2, 沼泽化草甸和湿草甸二者平均分别为23.5kg C/m2和235t C/hm2, 退化草甸最低(图 7), 分别为10.7kg C/m2和107t C/hm2, 退化草甸只有湿草甸的45%。海拔梯度不同群落中, SOC密度和储量草甸草毡层平均分别为17.7kg C/m2和177t C/hm2, 灌丛草甸平均分别为20.3kg C/m2和203t C/hm2, 灌丛草甸高出草甸15%。

|
| 图 7 水分和海拔梯度群落草毡层土壤有机碳密度和碳储量 Fig. 7 Soil organic carbon density and storage of sod layer in communities along the moisture and elevation gradients |
从图 8可以看出, 沼泽、退化沼泽、沼泽化草甸和湿草甸1m深度SOC密度和储量在55kg C/m2和550t C/hm2以上, 平均分别为60.4kg C/m2和604t C/hm2。干草甸和退化草甸明显低, SOC密度和储量平均分别为21.5kg C/m2和215t C/hm2, 退化草甸为湿草甸的32%。从沼泽→退化沼泽→沼泽化草甸→湿草甸→干草甸→退化草甸的水分递减梯度中, 0.3—1m土层SOC储量分别为354、387、471、347、115、81t C/hm2, 分别为草毡层的163%、212%、206%、146%、90%和76%。比较图 2看出, 过多过少的土壤水分, 都不利于SOC储存, 土壤水分有效性制约着土壤的有机碳储量。

|
| 图 8 水分梯度群落1m深度土壤有机碳密度和碳储量 Fig. 8 Soil organic carbon density and storage in the 1m depth soil layer in community along the moisture gradient |
高寒地区常年气温低, 湿度大, 冻结期长;植物生长期短, 生长缓慢, 土壤微生物活性低, 死亡后的根系难以分解, 以有机残体或粗有机质积累于土壤表层, 形成松软而坚韧的草毡层, 交织盘结耐搬运, 也容易与下层土壤分离, 是高寒生态系统重要的生态水文功能层, 类似于农业上的“耕作层”。
在青藏高原的许多地区, 草毡层之下就为基岩层, 砾石含量高, 质地坚硬[2];我们调查的海拔梯度群落草毡层下是风化程度低弱的母质层或基岩。关于表土层的划分不同地区不同研究者不同, 有的为表层40cm[22], 有的为20cm[23];有的采用土壤剖面A层[16, 19, 24], A层不同土壤不同地区厚度不同;大部分研究者采用30cm[20-21, 25-27](表 2)。我们的调查结果得出, 水分梯度群落土壤深厚, 草毡层厚度≥30cm;海拔梯度群落土壤浅薄, 草毡层厚度≤30cm(表 1), 高寒区草毡层平均厚度30cm, 与大部分研究者的表土层一致。
| 术语 Term |
位置和类型 Location and type |
深度 Depth/cm |
容重 Bulk density/(g/cm3) |
有机碳含量 Organic carbon content/(g/kg) |
有机碳密度 Organic carbon density/(kg C/m2) |
参考文献 References |
| 有机质层 Organic layer |
中国 | A层 | 0.16—34.37 | [19] | ||
| 土层 Soil layer |
中国北部草地 | 0—30 | 5.33 | [20] | ||
| 土层 Soil layer |
青藏高原 | 0—30 | 5.37 | [20] | ||
| 有机质层 Organic layer |
青藏高原 | A层 | 0.13—16.98 | [24] | ||
| 草毡表层 Mattic epipedon layer |
青藏高原东北部 | 0—14 | 4.99 | [34] | ||
| 土层 Soil layer |
青藏高原高山海拔4950 m | 0—30 | 0.50±0.01 | 13.7±1.9 | [26] | |
| 土层 Soil layer |
青藏高原高山海拔5300 m | 0—30 | 1.30±0.03 | 1.0±0.2 | [26] | |
| 土层 Soil layer |
奥地利阿尔卑斯山灌木地 | A层 | 245 | [16] | ||
| 土层 Soil layer |
瑞士阿尔卑斯山莎草草地 | A层 | 25-70 | [16] | ||
| 土层 Soil layer |
瑞士阿尔卑斯山 | 0—30 | 8.1—8.6 | [21] | ||
| 土层 Soil layer |
落基山脉草地 | A层 | 20-75 | [16] | ||
| 土层 Soil layer |
青海省高寒草甸 | 0—30 | 1.05 | 13.6 | [27] | |
| 草毡表层 Mattic epipedon layer |
海北高寒矮生嵩草草甸 | 0—30 | 1.08±0.10 | 44.1±14.4 | 12.9 | [25] |
| 土层 Soil layer |
海北原生高寒草甸 | 0—40 | 1.20±0.09 | 34.2 | 15.8±0.4 | [22] |
| 土层 Soil layer |
海北退化高寒草甸 | 0—40 | 1.46±0.06 | 19.1 | 11.2±0.3 | [22] |
| 土壤表层 Soil surface layer |
四川阿坝州高寒草甸 | 0—20 | 9.3—11.8 | [23] | ||
| 草毡层 Sod layer |
若尔盖高原沼泽湿地 | 0—30 | 0.26±0.01 | 312.3±13.4 | 21.8 | 本文 |
| 草毡层 Sod layer |
若尔盖高原湿草甸 | 0—30 | 0.73±0.02 | 109.3±9.8 | 23.9 | 本文 |
| 草毡层 Sod layer |
若尔盖高原干草甸 | 0—30 | 0.96±0.04 | 44.5±5.1 | 12.8 | 本文 |
| 草毡层 Sod layer |
若尔盖高原退化草甸 | 0—30 | 1.17±0.03 | 30.6±2.8 | 10.7 | 本文 |
| 草毡层 Sod layer |
若尔盖高原高山草甸 | 0—30 | 0.92±0.07 | 63.8±7.9 | 17.7±0.7 | 本文 |
| 草毡层 Sod layer |
若尔盖高原高山灌丛草甸 | 0—30 | 0.95±0.05 | 71.0±6.0 | 20.3±1.5 | 本文 |
| A层, A layer, 是土壤剖面表土层, 各地区不同土壤类型不同, 大致在0—25cm之内 | ||||||
土壤容重可以表明土壤的松紧程度及孔隙状况[28], 反映土壤的透水性、通气性和植物根系的阻力状况。青藏高原东部若尔盖高原草毡层土壤容重在0.3—1.2g/cm3之间(图 1, 表 2), 随着土壤含水量的降低(图 2)和草甸退化而提高, 但明显低于土壤容重平均值1.4g/cm3[29]。青海省海北退化草甸0—40cm表土层的容重达到1.46g/cm3[22] (表 2), 土壤容重随着土层加深而增大。得出, 高寒草毡层的容重小于1.4g/cm3。
土壤通过它的水分有效性、元素循环和土壤温度影响植被[30]。在植物生长季(5—9月份), 海北草毡寒冻雏形土土壤的平均含水量为36.0%[25], 我们测定的干草甸和退化草甸草毡层的含水量分别为32.7%和25.9%(图 2), 高山草甸和灌丛草甸的含水量为52.5%(图 2), 只有退化草甸草毡层质量含水量低于30%, 不同时期水分测定结果也证实了这一点。SOC密度随着土壤湿度的增加而增加, 当土壤含水量在30%以上时达到稳定状态[20]。高寒草甸的退化, 不仅造成草毡层碳储量显著下降, 而且将造成植被生长与恢复困难。高寒区草毡层质量含水量应保持在30%以上, 低于此值可能导致草地退化。
在青藏高原海拔4650m以上, pH值在6.4以下;海拔4400—4500m范围内pH值平均为7.0[26]。得出, 在海拔3440—4050m的范围内pH值平均为7.5, 在同一海拔高度不同水分条件下, 长期积水的沼泽湿地明显升高, 可达8.2。草毡层pH值在沼泽和积水低洼地高于草甸和高亢地;在较小的海拔跨度上变化不明显, 在较大的海拔跨度上, 随着海拔升高, pH值降低。
在所有植被类型中, 草甸的无机碳含量最小[31]。海北高寒矮生嵩草草甸的草毡寒冻雏形土表层(0—10cm)的土壤无机碳含量为5.0g/kg[25]。若尔盖高原草毡层土壤无机碳含量平地为7.9g/kg, 山地为4.6g/kg, 无机碳含量为SOC含量的7%。分析判断影响该地区土壤无机碳含量的主要因素是风, 其次是水, 该地区草毡层无机碳主要是风尘沉积物。
土壤机械组成是保持土壤质量的一个自然属性, 合适比例的土壤机械组成有利于植物根系活动以及从土壤中吸收水分和养分。本研究得出, 石砾含量越高的沼泽, 越容易退化;沙粒含量越高的草甸, 越容易退化。土壤粘粒能够通过粘粒胶体的吸附对SOC起到保护作用[32], SOC含量随着土壤湿度、粘粒和粉粒含量的提高而显著提高[14]。土壤养分主要吸附在细颗粒物质上, 在不同土壤组分中, 粘粒中SOC和总N含量最高, 其次是粉粒, 沙粒随着粒径的增大含量降到最少[33]。海北高寒矮生嵩草草甸的草毡寒冻雏形土表层(0—30cm)的SOC含量为44.1g/kg(表 2), 全N含量为3.8g/kg, 全P含量为0.9g/kg[25];SOC含量高于本研究的退化草甸草毡层(0—30cm), 与干草甸草毡层一致(表 2);全N高于本研究的退化草甸草毡层全N含量3.1g/kg, 低于干草甸的全N含量4.3g/kg(图 4);全P含量低于本研究的退化草甸全P含量1.6g/kg。海北退化高寒草甸0—40cm表土层的SOC为19.1g/kg, 原生高寒草甸在30g/kg以上[22], 阿尔卑斯山草地土壤剖面A层的SOC在25g/kg以上, 落基山脉草地A层的SOC在20g/kg以上[16](表 2), 本研究的退化草甸草毡层SOC为30.6g/kg, 综合看出, 高寒草毡层SOC含量低于30g/kg可能是草地退化的一个标志。
SOC是全球气候变化的敏感指标[35]。植物根系是SOC输入的主要来源, SOC来自植物分泌物及残体的归还, 其储量主要受植被光能利用率、群落分布、水位变化, 以及气候、土壤、人类活动等因素的影响[36]。气候变暖导致山区SOC损失, 但是海拔升高可以削弱气候变化的影响[37]。高寒草甸0—30cm土层的SOC含量在长期增温下无显著变化[38]。研究结果表明, 不同海拔高度群落草毡层SOC含量差异不显著(图 5)。温度升高会使SOC输入增加, 但同时会使SOC损失加快, 二者相互抵消。
草地灌丛化会改变土壤碳库[39]。位于奥地利蒂罗尔的阿尔卑斯山小灌木地A层SOC含量达到245g/kg, 明显高于阿尔卑斯山草地和落基山脉草地(表 2)[16]。本研究的高山(包括亚高山和高山)灌丛草甸草毡层SOC含量为71g/kg, 高于高山草甸草毡层。高山灌丛草甸草毡层SOC密度和储量比草甸高出15%。
在青藏高原海拔4400—5300m的高山-雪线梯度, 海拔4950m处, 植被盖度85%的群落表土层30cm范围内, SOC密度为13.7kg C/m2, 海拔5300m雪带植被盖度为0时, SOC密度很小(表 2) [26]。本研究的植被盖度除了退化草甸外, 都在80%以上(表 1), 在海拔3440—4050m的亚高山-高山梯度, 干草甸和退化草甸的草毡层SOC密度较低, 其他群落类型都较高, 最高的湿草甸达到24kg C/m2(图 7)。干草甸草毡层SOC密度与海北高寒矮生嵩草草甸草毡寒冻雏形土表层一致[25] (表 2)。比较看出(表 2), 青藏高原高寒草毡层(0—30cm表土层) SOC密度, 区域结果高于位于瑞士伯尔尼的阿尔卑斯山, 青藏高原东北部和全高原平均值与我国北部草地接近[20, 34], 都在我国土壤有机质层(A层)的范围[19]。植被盖度是生态系统功能和土壤退化的重要表征, 我国青藏高原东部高寒区不同群落草毡层SOC密度在10—24kg C/m2之间, SOC储量随着草甸退化和土壤水分有效性的降低而降低。
| [1] | Zheng D. The system of physico-geographical regions of the Qinghai-Xizang (Tibet) Plateau. Science in China (Series D):Earth Sciences, 1996, 39(4): 410–417. |
| [2] | 曹广民, 龙瑞军, 张法伟, 李以康, 林丽, 郭小伟, 韩道瑞, 李婧. 青藏高原高寒矮嵩草草甸碳增汇潜力估测方法. 生态学报, 2010, 30(23): 6591–6597. |
| [3] | 中国土壤系统分类研究丛书编委会. 中国土壤系统分类新论. 北京: 科学出版社, 1994: 302-309, 360-376. |
| [4] | 周兴民. 中国嵩草草甸. 北京: 科学出版社, 2001: 1–5. |
| [5] | 赵新全. 高寒草甸生态系统与全球变化. 北京: 科学出版社, 2009: 1–40. |
| [6] | Schlesinger W H. Evidence from chronosequence studies for a low carbon-storage potential of soils. Nature, 1990, 348(6298): 232–234. DOI:10.1038/348232a0 |
| [7] | Janzen H H. Carbon cycling in earth systems-a soil science perspective. Agriculture, Ecosystems & Environment, 2004, 104(3): 399–417. |
| [8] | Van Groenigen K J, Qi X, Osenberg C W, Luo Y Q, Hungate B A. Faster decomposition under increased atmospheric CO2 limits soil carbon storage. Science, 2014, 344(6183): 508–509. DOI:10.1126/science.1249534 |
| [9] | Post W M, Emanuel W R, Zinke P J, Stangenberger A G. Soil carbon pools and world life zones. Nature, 1982, 298(5870): 156–159. DOI:10.1038/298156a0 |
| [10] | Song G H, Li L Q, Pan G X, Zhang Q. Topsoil organic carbon storage of China and its loss by cultivation. Biogeochemistry, 2005, 74(1): 47–62. DOI:10.1007/s10533-004-2222-3 |
| [11] | Lal R. Soil carbon sequestration to mitigate climate change. Geoderma, 2004, 123(1/2): 1–22. |
| [12] | 王蓓, 孙庚, 罗鹏, 王萌, 吴宁. 模拟升温和放牧对高寒草甸土壤有机碳氮组分和微生物生物量的影响. 生态学报, 2011, 31(6): 1506–1514. |
| [13] | Kirschbaum M U F. Will changes in soil organic carbon act as a positive or negative feedback on global warming?. Biogeochemistry, 2000, 48(1): 21–51. DOI:10.1023/A:1006238902976 |
| [14] | Yang Y H, Fang J Y, Tang Y H, Ji C J, Zheng C Y, He J S, Zhu B. Storage, patterns and controls of soil organic carbon in the Tibetan grasslands. Global Change Biology, 2008, 14(7): 1592–1599. DOI:10.1111/j.1365-2486.2008.01591.x |
| [15] | 王襄平, 张玲, 方精云. 中国高山林线的分布高度与气候的关系. 地理学报, 2004, 59(6): 871–879. DOI:10.11821/xb200406009 |
| [16] | Körner C. Alpine plant life:functional plant ecology of high mountain ecosystems. Berlin: Springer, 2003: 9-11, 70-74. |
| [17] | 中国科学院《中国植物志》编委会. 中国植物志. 北京: 科学出版社, 1978: 156–158. |
| [18] | 曲仲湘, 吴玉树, 王焕校, 姜汉侨, 唐廷贵. 植物生态学.第二版. 北京: 高等教育出版社, 1990: 226–227. |
| [19] | Wu H B, Guo Z T, Peng C H. Distribution and storage of soil organic carbon in China. Global Biogeochemical Cycles, 2003, 17(2): 1048–1058. |
| [20] | Yang Y H, Fang J Y, Ma W H, Smith P, Mohammat A, Wang S P, Wang W. Soil carbon stock and its changes in northern China's grasslands from 1980s to 2000s. Global Change Biology, 2010, 16(11): 3036–3047. DOI:10.1111/gcb.2010.16.issue-11 |
| [21] | Hoffmann U, Hoffmann T, Jurasinski G, Glatzel S, Kuhn N J. Assessing the spatial variability of soil organic carbon stocks in an alpine setting (Grindelwald, Swiss Alps). Geoderma, 2014, 232-234: 270–283. DOI:10.1016/j.geoderma.2014.04.038 |
| [22] | 韩道瑞, 曹广民, 郭小伟, 张法伟, 李以康, 林丽, 李婧, 唐艳鸿, 古松. 青藏高原高寒草甸生态系统碳增汇潜力. 生态学报, 2011, 31(24): 7408–7417. |
| [23] | 杨树晶, 李涛, 干友民, 王永, 纪磊, 宋中齐, 刘焘. 阿坝高寒草甸土壤有机碳储量对不同利用方式与程度的响应. 中国草地学报, 2014, 36(6): 12–17. |
| [24] | 田玉强, 欧阳华, 徐兴良, 宋明华, 周才平. 青藏高原土壤有机碳储量与密度分布. 土壤学报, 2008, 45(5): 933–942. |
| [25] | 张金霞, 曹广民, 周党卫, 胡启武, 赵新全. 高寒矮嵩草草甸大气-土壤-植被-动物系统碳素储量及碳素循环. 生态学报, 2003, 23(4): 627–634. |
| [26] | Ohtsuka T, Hirota M, Zhang X Z, Shimono A, Senga Y, Du M G, Yonemura S, Kawashima S, Tang Y H. Soil organic carbon pools in alpine to nival zones along an altitudinal gradient (4400-5300 m) on the Tibetan Plateau. Polar Science, 2008, 2(4): 277–285. DOI:10.1016/j.polar.2008.08.003 |
| [27] | 刘淑丽, 林丽, 张法伟, 杜岩功, 李以康, 郭小伟, 欧阳经政, 曹广民. 放牧季节及退化程度对高寒草甸土壤有机碳的影响. 草业科学, 2016, 33(1): 11–18. DOI:10.11829/j.issn.1001-0629.2015-0286 |
| [28] | Vepraskas M J. Plant response mechanisms to soil compaction//Wilkinson R E, ed. Plant environment interaction. New York: Marcel Dekker Inc., 1994: 263-287. |
| [29] | 潘根兴. 中国土壤有机碳和无机碳库量研究. 科技通报, 1999, 15(5): 330–332. |
| [30] | Cheddadi R, Guiot J, Jolly D. The Mediterranean vegetation:what if the atmospheric CO2increased?. Landscape Ecology, 2001, 16(7): 667–675. DOI:10.1023/A:1013149831734 |
| [31] | Mi N, Wang S Q, Liu J Y, Yu G R, Zhang W J, Jobbágy E. Soil inorganic carbon storage pattern in China. Global Change Biology, 2008, 14(10): 2380–2387. DOI:10.1111/gcb.2008.14.issue-10 |
| [32] | 田佳倩, 周志勇, 包彬, 孙建新. 农牧交错区草地利用方式导致的土壤颗粒组分变化及其对土壤碳氮含量的影响. 植物生态学报, 2008, 32(3): 601–610. |
| [33] | He N P, Wu L, Wang Y S, Han X G. Changes in carbon and nitrogen in soil particle-size fractions along a grassland restoration chronosequence in northern China. Geoderma, 2009, 150(3/4): 302–308. |
| [34] | Yang R M, Zhang G L, Yang F, Zhi J J, Yang F, Liu F, Zhao Y G, Li D C. Precise estimation of soil organic carbon stocks in the northeast Tibetan Plateau. Scientific Reports, 2016, 6: 21842. DOI:10.1038/srep21842 |
| [35] | Xiao H L. Climate change in relation to soil organic matter. Soil and Environmental Sciences, 1999, 8(4): 300–304. |
| [36] | Davidson E A, Trumbore S E, Amundson R. Biogeochemistry:soil warming and organic carbon content. Nature, 2000, 408(6814): 789–790. DOI:10.1038/35048672 |
| [37] | Prietzel J, Zimmermann L, Schubert A, Christophel D. Organic matter losses in German Alps forest soils since the 1970s most likely caused by warming. Nature Geoscience, 2016, 9(7): 543–548. DOI:10.1038/ngeo2732 |
| [38] | 余欣超, 姚步青, 周华坤, 金艳霞, 杨月娟, 王文颖, 董世魁, 赵新全. 青藏高原两种高寒草甸地下生物量及其碳分配对长期增温的响应差异. 科学通报, 2015, 60(4): 379–388. |
| [39] | Brantley S T, Young D R. Shrub expansion stimulates soil C and N storage along a coastal soil chronosequence. Global Change Biology, 2010, 16(7): 2052–2061. DOI:10.1111/gcb.2010.16.issue-7 |