 2018, Vol. 38
2018, Vol. 38文章信息
- 陈潜, 许志春, 张连生, 陆鹏飞, 张永福.
- CHEN Qian, XU Zhichun, ZHANG Liansheng, LU Pengfei, ZHANG Yongfu.
- 褐梗天牛幼虫和成虫空间分布的地统计学研究
- Geostatistical analysis of the spatial distribution of Arhopalus rusticus larvae and adults
- 生态学报. 2018, 38(3): 975-983
- Acta Ecologica Sinica. 2018, 38(3): 975-983
- http://dx.doi.org/10.5846/stxb201611182350
-
文章历史
- 收稿日期: 2016-11-18
- 网络出版日期: 2017-10-18





2. 辽宁省建平县林业局, 朝阳 122400;
3. 北京市十三陵林场, 北京 102200
2. Jianping Bureau of Forestry, Chaoyang 122400, China;
3. Beijing Ming Tombs Forest Farm, Beijing 102200, China
任何一个种群都是在空间不同位置分布的, 由于种群内个体间的相互作用以及种群对环境的适应, 使得不同种群、同一种群在不同的环境条件下表现出不同的空间分布格局[1-2]。空间格局是昆虫种群非常重要的属性之一, 由昆虫所生活的环境条件和生物学特性所决定[3]。对昆虫空间格局的研究不但可以揭示种内和种间的空间构型, 而且还能为抽样设计和预测预报提供可靠的依据[4]。近年来, 在我国运用地统计学研究昆虫种群的空间分布已经取得了不小的成就, 因此利用地统计学的方法来探究昆虫种群的空间格局已备受研究学者所青睐[5-8]。
褐梗天牛(Arhopalus rusticus Linnaeus)是一种对松、杉、柏等林木危害非常严重的蛀干害虫[9]。火灾过后的松树的树桩里和濒临死亡树皮里会有大量的褐梗天牛[10]。研究发现褐梗天牛个体携带拟松材线虫的能力最强[11], 而拟松材线虫与松材线虫是近缘种, 形态习性相似, 褐梗天牛也有可能携带松材线虫[12]。为了监测褐梗天牛的发生, 揭示其种群的空间格局, 提供更为科学的虫情调查和预报预测技术。本文利用地统计学方法, 对其幼虫和成虫的空间分布进行调查研究。
1 试验地概况试验地设在辽宁省建平县青松岭乡的油松林中, 地理位置为41°80′N、119°84′E, 平均海拔700m以上, 年平均气温5.7℃, 年平均降水量487mm。森林覆盖率为49%。植被主要以松科为主, 2016年7月初, 在青松岭乡受褐梗天牛危害的油松林内, 抽取3块不同受害程度的油松林作为试验地。
2 研究方法 2.1 调查方法2016年7月—8月, 根据试验样地的大小及油松的分布状况, 在林地内分别抽取面积大小为90m×30m的样方, 3块样地分别采取逐株取样的调查方法。从油松的基部往上至树梢统计每株油松0—3m高度区域内每厘米的排粪孔和羽化孔数量, 分别代表天牛的幼虫和成虫数, 同时在坐标纸上记录样地中每株油松的相对空间位置。
2.2 分析方法 2.2.1 幼虫和成虫的垂直分布利用Excel软件, 对3块样地内褐梗天牛幼虫及成虫0—3m高度范围垂直分布进行绘图分析。
2.2.2 半方差函数半方差函数(Semi-variogram)为地统计学中独有的一种函数工具。它既能描述区域化变量的空间结构性变化, 又能描述其随机性变化。区域化变量在某方向上相距h的增量的方差, 称为区域变量在该方向上的变异函数, 其公式如下:

|
式中, γ(h)为两样点空间分隔距离h的半方差函数值, N(h)是分隔距离为h时的样本对数, Z(xi)与Z(xi+h)分别是Z(x)在空间位置xi和xi+h上的观测值[13]。半变异函数图是半变异函数γ(h)值对距离h的函数图, 如图 1它有3个特征参数:变程(range)、块金值(nugget)和基台值(sill)[14]。

|
| 图 1 经典的变异函数图 Fig. 1 A typical variogram |
变程:指区域化变量在空间上具有相关性的范围。在变程范围之内, 数据具有相关性;而在变程之外, 数据之间互不相关。
块金值:半方差函数与y轴相交的y值, 反映的是最小抽样尺度下变量的变异性及测量误差。
基台值:代表变量在空间上的总变异大小。
2.2.3 变异函数理论模型的拟合半变异函数常用的几种理论模型如线型、高斯型、指数型和球型都能用来拟合实验半变异函数[15]。线性模型说明研究的种群是随机分布或均匀分布, 球型揭示所研究的种群是聚集分布拟合过程中, 为了找到最佳理论模型, 应根据实际情况, 通过应用不同的理论模型对实际得出的变异曲线进行拟合, 比较他们之间决定系数、残差平方和、变程和块金值的大小选出最适理论模型[15-16]。但在实际观测中, 变异曲线有时表现为纯块金变异图, 即在所有距离上完全缺失空间相关性。例如, 昆虫的随机分布可能会导致纯块金变异图[17]。
2.2.4 空间分布图的生成Kriging法是地质统计学的重要组成部分, 也是地质统计学的核心。它根据样品空间位置不同、样品间相关程度的不同, 对每个样品位赋予不同的权, 进行滑动加权平均, 以估计中心块段平均品位。其公式如下:
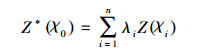
|
式中, Z*(χ0)是点(x0, y0)处的估计值, λi是权重系数, 它同样是用空间上所有已知点的数据加权求和来估计未知点的值。克里金法不仅考虑待估点位置与已知数据位置的相互关系, 而且还考虑变量的空间相关性[18]。
在选择合适的拟合模型, 得出最优函数曲线后, 借助克里金插值法对褐梗天牛幼虫和成虫在样地中的数量进行插值分析, 利用绘图软件Surfer 8.0生成其空间分布图[19]。
3 结果分析 3.1 不同受害程度样地内褐梗天牛幼虫及成虫的分布情况通过调查得出样地油松的树高、胸径和受害情况见表 1。
| 样地编号 Stand number |
林分类型 Forest types |
调查株数/株 Number of investigated trees |
平均树高 Average height/m |
平均胸径 Average breast diameter/cm |
幼虫数/头 Number of larvae |
成虫数/头 Number of adults |
受害株率/% Rate of infested trees |
| 1 | 轻度受害林 | 91 | 6.6 | 22.0 | 153 | 222 | 30.8 |
| 2 | 中度受害林 | 88 | 5.6 | 24.2 | 210 | 221 | 44.3 |
| 3 | 重度受害林 | 69 | 4.8 | 21.6 | 378 | 351 | 78.3 |
轻度、中度、重度3块不同受害林分样地内幼虫的垂直分布图见图 2, 从图 2可以看出, 褐梗天牛幼虫在油松上的分布具有明显的垂直分布特性, 轻度受害林中幼虫数量在24—25cm处密度最大, 中度受害林幼虫数量在9—10cm处密度最大, 重度受害林幼虫数量在13—14cm处密度最大, 虽然天牛数量在某些高度范围出现了小高峰, 但整体变化基本都是从最大密度向两边高度减小。

|
| 图 2 不同受害林分褐梗天牛幼虫垂直分布结果 Fig. 2 Statistical characteristics of the vertical distributions of the larva of Arhopalus rusticus in different forests |
轻度、中度、重度3块不同受害林分样地内幼虫的垂直分布图见图 3, 从图 3可以看出, 褐梗天牛幼虫在油松上的分布具有明显的垂直分布特性, 轻度受害林中幼虫数量在4—5cm处密度最大, 中度受害林幼虫数量在26—27cm处密度最大, 重度受害林幼虫数量在5—6cm处密度最大, 虽然天牛数量在某些高度范围出现了小高峰, 但整体变化基本都是从最大密度向两边高度减小。

|
| 图 3 不同受害林分褐梗天牛成虫垂直分布结果 Fig. 3 Statistical characteristics of the vertical distributions of the adult of Arhopalus rusticus in different forests |
通过GS+软件对3种不同受害程度样地内褐梗天牛幼虫数量空间分布进行地统计分析, 其半变异函数理论模型及各参数和空间分布表如表 2。
| 林分类型 Forest types |
理论模型 Theoretic model |
C0 | C | Sill | C/Sill | A | R2 | RSS | 分布型 Distribution |
| 轻度受害林Lightly-infested | 高斯模型 | 0.0194 | 0.0354 | 0.0548 | 0.646 | 19.10 | 0.708 | 0.147 | 聚集分布 |
| 中度受害林Medium-infested | 高斯模型 | 0.3320 | 1.2040 | 1.536 | 0.784 | 11.97 | 0.837 | 0.124 | 聚集分布 |
| 重度受害林Heavily-infested | 指数模型 | 1.2090 | 1.2100 | 2.419 | 0.500 | 61.98 | 0.749 | 0.331 | 聚集分布 |
| *C0:块金常数Nugget, C:偏基台值Partial sill, Sill:基台值Sill, C/Sill:空间变异Spatial variation, A:空间依赖范围Range, R2:决定系数Decisive coefficient, RSS:残差平方和Residual sum of square | |||||||||
由图 4和表 2可以看出, 轻度受害林内褐梗天牛幼虫种群的空间分布格局呈现较明显的空间聚集状态, 决定系数为0.708, 说明幼虫的空间分布具有较强的空间依赖性, 其变程为19.1m, 说明在此轻度受害林中该距离范围以内任何两样点褐梗天牛幼虫数量都存在着一定的空间相关关系, 空间变异值为0.646, 说明在此轻度受害林中褐梗天牛幼虫数量的总空间变异有64.6%来自于空间自相关现象。块金值为0.0194, 产生块金值的主要原因可能有:①试验地虫口密度较低。②由于调查的受危害油松上的排粪孔数量较多且新旧混有, 在统计过程中容易出现误差。

|
| 图 4 轻度受害林中褐梗天牛幼虫种群空间分布的变异函数图与等值线矢量叠加图 Fig. 4 Variogram and overlaying the contour and vector plots of the population number for the larva of Arhopalus rusticus in lightly infested stand of forest |
由褐梗天牛幼虫种群数量的等值线与矢量叠加图可以看出, 此样地中有几个明显的幼虫分布聚集点, 幼虫数量随着聚集中心点向周围逐渐减少, 反映了幼虫的聚集强度和扩散方向, 显示了其聚集分布的特点。
3.3.2 中度受害林内褐梗天牛幼虫种群数量的空间分析由图 5和表 2可以看出, 轻度受害林内褐梗天牛幼虫种群的空间分布格局呈现较明显的空间聚集状态, 决定系数为0.837, 说明幼虫在中度受害林内的空间依赖性比轻度受害林的强, 其变程为11.97m, 说明在此轻度受害林中该距离范围以内任何两样点褐梗天牛幼虫数量都存在着一定的空间相关关系, 空间变异值为0.784, 说明在此轻度受害林中褐梗天牛幼虫数量的总空间变异有78.4%来自于空间自相关现象。块金值为0.332, 产生块金值的主要原因可能有:①试验地内时常有人砍柴, 造成一定量的衰弱木减少, 进而使幼虫分布数量相对减少。②试验地所处海拔较高, 在500m以上, 试验地内大块碎石较多。

|
| 图 5 中度受害林中褐梗天牛幼虫种群空间分布的变异函数图与等值线矢量叠加图 Fig. 5 Variogram and overlaying the contour and vector plots of the population number for the larva of Arhopalus rusticusin medium infested stand of forest |
由褐梗天牛幼虫种群数量的等值线与矢量叠加图可以看出, 此样地中有几个明显的幼虫分布聚集点, 幼虫数量随着聚集中心点向周围逐渐减少, 反映了幼虫的聚集强度和扩散方向, 显示了其聚集分布的特点。
3.3.3 重度受害林内褐梗天牛幼虫种群数量的空间分析由图 6和表 2可以看出, 轻度受害林内褐梗天牛幼虫种群的空间分布格局呈现较明显的空间聚集状态, 决定系数为0.749, 说明幼虫在重度受害林内的空间依赖性比中度受害林的较弱, 其变程为61.98m, 说明在此轻度受害林中该距离范围以内任何两样点褐梗天牛幼虫数量都存在着一定的空间相关关系, 空间变异值为0.5, 说明在此轻度受害林中褐梗天牛幼虫数量的总空间变异有50%来自于空间自相关现象。块金值为1.209, 该样地产生的块金值最大, 原因可能有:①试验地有人为砍伐树木的迹象, 附近处在进行褐梗天牛的引诱试验, 可能影响其数量的分布。②由于调查的受危害油松上的排粪孔数量较多且新旧混有, 在统计过程中容易出现误差。

|
| 图 6 重度受害林中褐梗天牛幼虫种群空间分布的变异函数图与等值线矢量叠加图 Fig. 6 Variogram and overlaying the contour and vector plots of the population number for the larva of Arhopalus rusticus in heavily infested stand of forest |
由褐梗天牛幼虫种群数量的等值线与矢量叠加图可以看出, 此样地中的褐梗天牛幼虫密度最大, 聚集性最强, 仍然有几个幼虫聚集点, 且幼虫数量随聚集点向周围扩散逐渐减少, 同样反映了幼虫的扩散方向及分布情况。
3.4 褐梗天牛成虫的空间分布分析通过GS+软件对3种不同受害程度样地内褐梗天牛成虫数量空间分布进行地统计分析, 其半变异函数理论模型及各参数和空间分布如表 3。
| 林分类型 Forest types |
理论模型 Theoretic model |
C0 | C | Sill | C/Sill | A | R2 | RSS | 分布型 Distribution |
| 轻度受害林Lightly-infested | 线性模型 | 1.71 | 0.07 | 1.78 | 0.044 | 43.08 | 0.018 | 0.456 | 随机分布 |
| 中度受害林Medium-infested | 线性模型 | 2.16 | 0.05 | 2.21 | 0.021 | 43.23 | 0.002 | 1.620 | 随机分布 |
| 重度受害林Heavily-infested | 线性模型 | 1.82 | 0.38 | 2.20 | 0.171 | 44.17 | 0.229 | 0.624 | 随机分布 |
由图 7和表 3可以看出, 3种林分中的褐梗天牛成虫的空间分布格局呈现很弱的聚集性, 非常接近于随机分布。3种林分中的空间变异值从小到大依次为0.021, 0.044, 0.171, 说明在这些样地中总空间变异只有很小一部分是由空间依赖性产生的, 块金常数所占比例很高, 分别为96.1%、97.9%、82.7%, 变异曲线图都非常接近于纯块金变异图, 这种情况下的变异曲线参数与正常变异曲线参数差别较大, 只根据某个方面下定结论是不准确的。块金值较大, 主要是由于在成虫羽化盛期, 时常有人在试验地内放羊, 对成虫数量有一定影响, 其次是在调查过程中存在误差。

|
| 图 7 褐梗天牛成虫种群空间分布的变异函数图 Fig. 7 Variogram of the population number for the adult of Arhopalus rusticus |
在我国褐梗天牛的生活周期为2—3a[20], 整个幼虫期仅有一个排粪孔, 羽化成成虫后钻蛀一个羽化孔钻出, 因此本试验以样地中油松树上排粪孔的数量为调查指标是准确可靠的。本试验首次利用地统计学对褐梗天牛幼虫空间格局进行研究, 得到褐梗天牛幼虫在3种受害程度不同的油松林中, 其空间格局均呈现聚集分布, 且一致性较为明显。在3种不同受害程度的油松林中, 褐梗天牛幼虫的空间分布格局均呈现明显的聚集分布状态, 轻度、中度、重度受害林的变异函数理论模型分别是高斯模型、高斯模型、指数模型, 表示其幼虫聚集状态既有相似的也有不同的。从其空间分布图可以看出, 不同受害样地中都有几个幼虫分布的聚集点, 幼虫从聚集点向四周扩散, 但扩散的并不明显。褐梗天牛幼虫自身不具备扩散能力, 成虫的飞行扩散能力及产卵习性则成为决定幼虫空间分布格局的重要因素之一, 而成虫的寄主, 以油松、樟子松、侧柏等树种为主, 对成虫的扩散行为以及产卵刻槽具有重要影响。其次, 寄主树木的树高、胸径等外部因素也会影响幼虫及成虫的种群空间特征。在对褐梗天牛成虫的野外跟踪调查中发现, 成虫羽化后并没有补充营养, 性成熟后雌虫会选择衰弱树或新枯死的树产卵, 部分雌虫还产卵于土壤中, 因此林间树势衰弱与新枯死的分布状况与褐梗天牛的聚集分布特征具有密切相关性, 这与空间分布图中得出的结果中的几个分布聚集点基本吻合, 且受害林受害程度越大, 聚集点个数也随之增多。
在3种不同程度受害林中, 褐梗天牛成虫的变异函数曲线图均接近于纯块金变异图, 这表明在该3块样地中褐梗天牛成虫数量在空间分布是随机的。空间分布图表明, 3块样地中的成虫在空间中仍有几个较弱的聚集点, 但整体均匀分布。这可能是由于该样地中衰弱树木分布较均匀与成虫飞行扩散能力较强等因素所致。在半变异函数曲线图(图 2, 图 3和图 4)中均可以看到, 假设采样距离(h)超过一定值继续增大时半变异函数值和变程的变化可能有不同的响应, 所以在以后的调查研究中需注意采样的空间尺度, 增大可能对这种空间分布的分析更为全面[19]。另外, 由于调查的年限较短, 调查的面积和地点的限制, 应用地统计学对褐梗天牛种群在时间上的依赖性与连续性的研究今后还需增加调查次数, 获得更多资料进一步完善, 以便于更好地为大尺度上的病虫害发生预测提供依据。
褐梗天牛幼虫和成虫在油松上的垂直分布特征表明, 褐梗天牛幼虫和成虫在油松上主要集中聚集在某个高度, 然后随着高度增加密度降低, 随着高度接近地面密度也降低, 且分布量随受害程度的增加有明显的增大趋势。在当地对褐梗天牛的防治应多集中在树干基部, 明确几个天牛的分布聚集点, 沿着其扩散方向逐步防治, 这样既可以达到较好的防治效果又节约了防治成本。
目前, 对褐梗天牛虫害的防治, 通常采用物理、化学以及生物防治相结合进行。在产卵期人工砸卵, 可将危害率大大降低。采用5%的杀螟松、硫丹、10%敌敌畏和3%除虫菊酯乳浊液混合使用, 可以有效防止锯木不被褐梗天牛入侵[21];在被害林木附近或林中存在大量成虫的地方喷洒农药可大量毒杀成虫。利用天敌防治, 在产卵期释放卵寄生蜂或在幼虫期释放管氏硬皮肿腿蜂、花绒寄甲, 可达到防治效果。另外, 在营林时, 切勿与松、杉、柏科等褐梗天牛的寄主树种混交种植。
| [1] | Matheron G. Principles of geostatistics. Economic Geology, 1963, 58(8): 1246–1266. DOI:10.2113/gsecongeo.58.8.1246 |
| [2] | Matheron G. Kriging, or polynomial interpolation procedures. Canadian Mining and Metallurgical Bulletin, 1967, 60: 1041–1045. |
| [3] | 徐汝梅. 昆虫种群生态学. 北京: 北京师范大学出版社, 1987. |
| [4] | 丁岩钦. 昆虫数学生态学. 北京: 科学出版社, 1994. |
| [5] | Fuentes M, Chaudhuri A, Holland D M. Bayesian entropy for spatial sampling design of environmental data. Environmental and Ecological Statistics, 2007, 14(3): 323–340. DOI:10.1007/s10651-007-0017-0 |
| [6] | Li L F, Wang J F, Liu J Y. Optimal decision-making model of spatial sampling for survey of China's land with remotely sensed data. Science in China Series D:Earth Sciences, 2005, 48(6): 752–764. DOI:10.1360/03yd0376 |
| [7] | Gust N, Choat J H, McCormick M I. Spatial variability in reef fish distribution, abundance, size and biomass:a multi-scale analysis. Marine Ecology Progress Series, 2001, 214: 237–251. DOI:10.3354/meps214237 |
| [8] | Stark K E, Arsenault A, Bradfield G E. Variation in soil seed bank species composition of a dry coniferous forest:spatial scale and sampling considerations. Plant Ecology, 2008, 197(2): 173–181. DOI:10.1007/s11258-007-9368-8 |
| [9] | 泽桑梓, 赵涛, 季梅, 张星耀, 闫争亮. 褐梗天牛寄主挥发性成分分析和林间引诱效果. 动物学研究, 2011, 32(增刊): 139–146. |
| [10] | Whitehouse N J. Forest fires and insects:palaeoentomologicalresearch from a subfossil burntforest. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 164(1/4): 231–246. |
| [11] | 赵锦年, 余盛明, 姚剑飞, 林长春, 丁德贵, 王浩杰. 黄山风景区松材线虫病危险性评估:Ⅱ.松天牛携带线虫状况的监测. 林业科学研究, 2004, 17(1): 72–76. |
| [12] | 高娜, 姚洪锡, 姜雪峰, 张栩, 李艳, 卢希平. 松材线虫染病黑松上松褐天牛与褐梗天牛生态位的研究. 中国森林病虫, 2013, 32(5): 4–7. |
| [13] | 章一巧, 宗世祥, 刘永华, 温俊宝, 闫占文, 骆有庆. 陕西省栎黄枯叶蛾蛹的空间分布. 生态学报, 2012, 30(20): 6308–6317. |
| [14] | 宗世祥, 骆有庆, 许志春, 温俊宝, 贾峰勇. 沙棘木蠹蛾卵和幼虫空间分布的地统计学分析. 生态学报, 2005, 25(4): 831–836. |
| [15] | 李咏玲, 韩福生, 张金桐. 昆虫性信息素研究综述. 山西农业科学, 2010, 38(6): 51–54. |
| [16] | 赵静, 赵鑫, 王玉军, 李光强, 刘丽平, 孟家华, 郑方强. 烟盲蝽及其天敌蜘蛛空间格局的地统计学分析. 生态学报, 2010, 30(15): 4196–4205. |
| [17] | 李友常, 夏乃斌, 屠泉洪, 骆有庆, 温俊宝. 杨树光肩星天牛种群空间格局的地统计学研究. 生态学报, 1997, 17(4): 393–401. |
| [18] | 向昌盛, 袁哲明. 地统计方法在昆虫学研究中的应用. 中国农学通报, 2009, 25(17): 191–194. |
| [19] | 胡瑜珩, 武海卫, 许志春. 云斑天牛幼虫空间分布格局的地统计学分析. 西北林学院学报, 2015, 30(5): 165–171. |
| [20] | 张霖. 褐梗天牛生物学特性和引诱剂的初步研究[D]. 北京: 北京林业大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10022-1016145147.htm |
| [21] | Dominik J. Summer control of xylophagous insect pests in Scots pine stands. Sylwan, 1981, 125(7/9): 111–117. |