 2018, Vol. 38
2018, Vol. 38文章信息
- 包欣欣, 刘丙万.
- BAO Xinxin, LIU Bingwan.
- 黑龙江凉水和丰林保护区鸟类和兽类多样性
- Avian and mammalian biodiversity of Heilongjiang Liangshui and Fenglin Reserve
- 生态学报. 2018, 38(2): 502-510
- Acta Ecologica Sinica. 2018, 38(2): 502-510
- http://dx.doi.org/10.5846/stxb201608291760
-
文章历史
- 收稿日期: 2016-08-29
- 网络出版日期: 2017-09-14

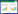




生物多样性监测和评价是自然保护区的重要工作[1-4]。鸟类和兽类是生物多样性保护和评价中的重要指示类群[4-6]。生物多样性调查主要使用样线法[7]、样带法[8]、大样方调查法[9]、访问法[10]以及近些年广泛运用的红外相机监测法[11-12]。红外相机作为一种具有人为干扰小、实时性强、非损伤性等特点的取样技术, 被广泛应用于鸟兽的监测与研究中[13]。
本文以凉水和丰林保护区为例探讨红外相机技术在鸟兽多样性中的应用, 并对保护区内鸟兽多样性进行研究, 掌握重要鸟兽物种的种群现状, 同时对凉水和丰林保护区内鸟兽多样性进行了比较研究。
1 研究地概况凉水保护区位于黑龙江省伊春市带岭区, 地处小兴安岭山脉的东南段——达里带岭支脉的东坡, 128°47′8″—128°57′19″E, 47°6′49″—47°16′10″N, 面积为12133hm2, 森林覆盖率达95%以上;丰林保护区位于黑龙江省伊春市五营区, 地处小兴安岭南坡北段, 128°58′—129°15′E、48°02′—48°12′N, 总面积18165hm2, 森林覆被率约为96%;凉水保护区和丰林保护区的主要保护对象是以红松为主的温带针阔混交林生态系统(图 1)。

|
| 图 1 凉水保护区、丰林保护区位置示意图 Fig. 1 The location of Liangshui and Fenglin Reserve |
东北林业大学分别于2005年和2009年在黑龙江凉水和丰林国家级自然保护区(以下简称为凉水和丰林保护区)建立了一块9hm2和30hm2的典型红松林监测样地, 用于长期监测该区域植物群落动态。为了研究保护区的鸟兽资源现状, 分别于2013年12月到2015年5月在凉水样地、2014年4月到2015年5月在丰林样地设置30个红外相机监测点, 每个监测点布设相机1台, 布设密度0.5台/hm2, 总覆盖面积为60hm2, 用于林下鸟类和兽类的初步监测。
将相机用绳子固定于树干上, 距离地面60—100cm保证相机与地面平行。尽量避免拍摄镜头内有干扰物, 确保视线开阔, 避免阳光直射。
进行相机、SD卡检测, 确保正常工作后将相机和SD卡统一编号, 相机和SD卡编号一一对应。统一对相机参数进行设置, 包括日期、时间、闪光、拍摄间隔、灵敏度。详细记录相机安装位点的地理坐标、林班号、树牌号、生境。
将相片逐一挑选去除无效照片, 上传于CameraData数据系统, 进行物种识别。
2.2 数据分析对相同相机位点及相近时间拍摄的同种个体进行识别, 确定是否为同种个体, 独立有效照片的确定标准为同一相机位点含同种个体的相邻有效照片间隔时间至少为30min[14]。
1) 相对丰富度(RAI):
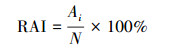
|
将各物种出现的独立有效照片数与所拍到的动物的总照片数的百分比作为各物种的相对丰富度(RAI)[15]。式中, Ai表示第i(i=1, 2, 3, 4…, 28)类物种出现的独立照片数, N表示独立相片总数。
2) 捕获率(CR):
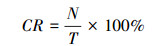
|
根据在相同位点同种物种的独立照片的相对数量, 即捕获率(CR)[16], 式中N为拍摄到某一物种的独立照片数量, T为总有效监测日。
计算结果如下定义:常拍种(>10%)、较常拍种(1%—10%)、偶拍种(0.1%—1%)和罕拍种(< 0.1%)。
3) Shannon-Wiener指数(H):

|
其中:Pi为第i种个体数占群落内总个体数的比例, 此处以i物种的独立照片数占总独立照片数的比例代替。
4) 均匀度指数(J′)
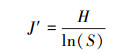
|
其中H为Shannon-Wiener指数, S为监测到的总物种数。
5) Sorenson指数(Cs):
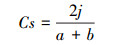
|
式中:j为两个保护区中的共有种, a和b分别为两个保护区的物种数。
6) 利用EstimateS9.0进行物种与相机数拟合的稀疏化, 利用ACE(基于多度的物种估计量)、ICE (基于盖度的物种估计量)的估计方法进行物物种丰富度估计。
3 结果与分析 3.1 鸟类、兽类组成2013年12月至2015年5月、2014年4月至2015年5年利用红外相机分别对凉水样地和丰林样地内的鸟兽资源进行监测。凉水、丰林样区共发现鸟兽29种, 隶属9目18科。其中兽类中, 偶蹄目2科2种, 兔形目1科1种, 食肉目1科4种。啮齿目3科5种。鸟类中雀形目7科13种, 鴷形目1科1种, 鸮形目1科1种, 鸡形目1科1种, 隼形目1科1种(表 1)。
| 分类 Taxonomy | 独立照片Photograph | 区系 Fauna | 分布型 Distribution | |
| 凉水Liangshui | 丰林Fenglin | |||
| Ⅰ食肉目Carnivora | ||||
| 1 鼬科 Mustelida | ||||
| (1)黄鼬Mustela sibirica | 1 | 4 | 广布种 | 古北型 |
| (2)伶鼬Mustela nivalis | 1 | 古北界 | 古北型/东洋型 | |
| (3)紫貂Martes zibellina | 2 | 古北界 | 古北型 | |
| (4)狗獾Meles meles | 6 | 23 | 广布种 | 古北型 |
| Ⅱ偶蹄目Artiodactyla | ||||
| 2 鹿科 Cervida | ||||
| (5)西伯利亚狍Capriolus pygargus | 80 | 31 | 古北界 | 古北型 |
| 3 猪科 Suidae | ||||
| (6)野猪Sus scrofa | 1 | 1 | 广布种 | 古北型 |
| Ⅲ兔形目Lagomorpha | ||||
| 4 兔科 Leporidae | ||||
| (7)东北兔Lepus mandshuricus | 1 | 古北界 | 东北型(北部为主) | |
| Ⅳ啮齿目Rodentia | ||||
| 5 松鼠科 Sciuridae | ||||
| (8)松鼠Sciurus vulgari | 117 | 235 | 广布种 | 古北型 |
| (9)花鼠Eutamias sibiricus | 260 | 314 | 古北界 | 古北型 |
| 6 仓鼠科 Circetidae | ||||
| (10)棕背Clethrionomys rufocanus | 3 | 18 | 古北界 | 古北型 |
| 7 鼠科 Muridae | ||||
| (11)大林姬鼠Apodemus peninsulae | 1 | 4 | 广布种 | 东北-华北型 |
| (12)黑线姬鼠Apodemus agrarius Pallas | 1 | 古北界 | 古北型 | |
| Ⅴ雀形目Passeriformes | ||||
| 8 山雀科 Paridae | ||||
| (13)沼泽山雀Parus palustris | 2 | 3 | 古北界 | 古北型 |
| 9 燕雀科 Passeriformes | ||||
| (14)黑头蜡嘴雀Eophona personata | 3 | 2 | 广布种 | 东北型(东部为主) |
| 10 鳾科 Sittidae | ||||
| (15)普通鳾Sitta europaea | 20 | 57 | 东洋界 | 古北型 |
| 11 鹀科 Emberizidae | ||||
| (16)白眉鹀Emberiza tristram | 4 | 6 | 广布种 | 东北型(北部为主) |
| 12 伯劳科 Laniidae | ||||
| (17)红尾伯劳Lanius cristatus | 1 | 广布种 | 东北-华北型 | |
| 13 鹟科 Muscicapidae | ||||
| (18)灰背鸫Turdus hortulorum | 4 | 24 | 古北界 | 东北型(北部为主) |
| (19)白腹鸫Turdus pallidus | 94 | 41 | 古北界 | 东北型(北部为主) |
| (20)斑鸫Turdus eunomus | 1 | 广布种 | 东北型(北部为主) | |
| (21)白眉地鸫Zoothera sibirica | 2 | 4 | 古北界 | 东北型(北部为主) |
| (22)紫啸鸫Myophonus caeruleus | 1 | 广布种 | 东洋型 | |
| (23)虎斑地鸫Zoothera dauma | 1 | 东洋界 | 古北型 | |
| (24)红胁蓝尾鸲 Tarsiger cyanurus | 1 | 广布种 | 东北型(北部为主) | |
| 14 鸦科 Corvidae | ||||
| (25)松鸦Garrulus glandarius | 1 | 2 | 广布种 | 古北型 |
| Ⅵ鸡形目Galliformes | ||||
| 15 雉科 Phasianidae | ||||
| (26)花尾榛鸡Bonasia bonasia | 9 | 10 | 古北界 | 古北型 |
| Ⅶ鴷形目Piciformes | ||||
| 16 啄木鸟科 Picidae | ||||
| (27)白背啄木鸟Dendrocopos leucotos | 4 | 3 | 广布种 | 古北型 |
| Ⅷ鸮形目Strigiformes | ||||
| 17 鸱鸮科 Strigidae | ||||
| (28)长尾林鸮Strix uralensis | 1 | 古北界 | 古北型 | |
| Ⅸ隼形目Falconiformes | ||||
| 18 鹰科 Accipitridae | ||||
| (29)松雀鹰Accipiter virgatus | 1 | 广布种 | 东洋型 | |
凉水保护区内累计捕获日为10736, 累计获得独立照片621张, 经过鉴定, 隶属于9目14科22种。兽类独立照片514张, 古北界5种, 广布种6种, 古北型9种。共4目7科11种, 偶蹄目2科2种, 兔形目1科1种, 食肉目1科3种, 啮齿目3科5种。其中国家一级保护动物1种-紫貂(Martes zibellina)。丰林保护区累计捕获日为7460, 累计获得独立照片884张, 隶属于8目16科25种。兽类独立照片638张, 古北界4种, 广布种5种, 古北型8种。共3目6科9种, 偶蹄目2科2种;食肉目1科3种;啮齿目3科4种。保护区兽类以古北界及广布种为主, 分布型以古北型为主(表 1)。
凉水保护区获得鸟类独立照片107张, 古北界5种, 广布种5种, 古北型6种, 东北型4种, 共4目8科11种, 鸟类中雀形目5科8种, 鴷形目1科1种, 鸡形目1科1种, 隼形目1科1种。国家二级保护动物2种, 为花尾榛鸡(Bonasia bonasia)和松雀鹰(Accipiter virgatus)(表 1)。丰林保护区获得鸟类独立照片166张, 古北界6种, 广布种9种, 古北型7种, 东北型6种;共4目10科16种, 雀形目7科13种, 鴷形目1科1种, 鸡形目1科1种, 鸮形目1科1种。国家二级保护动物2种, 分别为花尾榛鸡和长尾林鸮(Strix uralensis)保护区鸟类以古北界和广布种为主, 分布型以古北型和东北型占优势(表 1)。
3.2 凉水、丰林保护区物种稀疏化曲线利用EstimateS9.0对凉水和丰林保护区内兽类、鸟类物种数及二者之和与红外相机台数进行拟合的稀疏化曲线(图 2—图 4), 对两个保护区进行对比。

|
| 图 2 鸟类物种数与红外相机台数所拟合的稀疏化曲线 Fig. 2 Rarefaction curves (Sobs) for estimating species diversity of avians with increased camera traps |

|
| 图 3 兽类物种数与红外相机台数所拟合的稀疏化曲线 Fig. 3 Rarefaction curves (Sobs) for estimating species diversity of mammals with increased camera traps |

|
| 图 4 鸟、兽物种数与红外相机台数所拟合的稀疏化曲线 Fig. 4 Rarefaction curves (Sobs) for estimating species diversity of species with increased camera traps |
从稀疏化曲线可以看出鸟类和兽类及二者之和随着相机数目的增加而增加;监测中兽类曲线在急剧上升后变为渐近线, 上升舒缓慢, 说明取样充分, 鸟类的曲线比兽类的更陡, 说明鸟类取样不很充分;图 3中两条曲线相交说明交点前后物种多样性大小排序相反。
通过EstimateS9.0获得凉水鸟类ACE值为12.17, ICE值为13.82, 这2种方法对保护区目标鸟类的物种丰富度估计为12种和14种。丰林保护区鸟类ACE值为18.78, ICE值为22.46, 保护区目标鸟类的物种丰富度估计大约为19种和22种。从监测结果看, 凉水实际监测到的鸟类种数目为11种, 约为全部物种的78%—91%。丰林实际监测到的鸟类物种数目为16种, 约为全部物种的72%—84%, 保护区内大部分鸟类被监测到。
凉水兽类ACE值为13.69, ICE值为13, 丰林兽类ACE值为9.46, ICE值为9.63, 凉水实际监测到的兽类种数目为11种, 约为全部物种的78%—84%。丰林实际监测到的兽类物种数目为9种, 约为全部物种的90%—95%, 保护区内大部分兽类被监测到。
凉水鸟兽ACE值为28.7, ICE值为31.17。丰林鸟兽ACE值为28.1, ICE值为31.27。凉水实际监测到的鸟兽物种数目为22种, 约为全部物种的70%—78%。丰林保护区实际监测到的鸟兽物种数目为25种, 约为全部物种的80%—89%, 保护区内大部分鸟兽被监测到。
3.3 凉水、丰林保护区鸟兽多样性凉水和丰林保护区鸟、兽多样性及物种多样性无显著差异, 均匀度指数相近(表 2)。
| 地点 Sites | 物种数 Number of species | Shannon-Wiener 指数H | Shannon-Wiener 指数(兽)H′ | Shannon-Wiener 指数(鸟)H″ | 均匀度指数 J′ |
| 凉水保护区 | 21 | 2.556 | 1.732 | 1.880 | 0.840 |
| 丰林保护区 | 25 | 2.518 | 1.659 | 2.668 | 0.782 |
凉水、丰林保护区物种相似度指数为Cs=0.621, 说明两地的物种有较多重叠。
3.4 凉水、丰林保护区物种丰富度凉水、丰林保护区内相对丰富度最高的兽类均为松鼠和花鼠, 其次为西伯利亚狍。凉水保护区兽类相对丰富度最低的是野猪(Sus scrofa)和东北兔(Lepus mandshuricus), 丰林保护区兽类相对丰富度最低的是香鼬(Mustela altaica);鸟类中相对丰富度最高的是普通鳾(Sitta europaea)和白腹鸫(Turdus pallidus)(表 3)。
| 鸟类Avians | 兽类Mammals | |||||
| 物种 | 凉水 | 丰林 | 物种 | 凉水 | 丰林 | |
| 沼泽山雀Parus palustris | 0.32 | 0.34 | 黄鼬Mustela sibirica | 0.16 | 0.79 | |
| 黑头蜡嘴雀Eophona personata | 0.48 | 0.23 | 伶鼬Mustela nivalis | 0.11 | ||
| 普通鳾Sitta europaea | 3.21 | 6.44 | 紫貂Martes zibellina | 0.32 | ||
| 白眉鹀Emberiza tristram | 0.64 | 0.68 | 狗獾Meles meles | 4.17 | 2.60 | |
| 红尾伯劳Lanius cristatus | 0.11 | 西伯利亚狍狍Capriolus pygargus | 12.52 | 3.50 | ||
| 灰背鸫Turdus hortulorum | 0.64 | 2.71 | 野猪Sus scrofa | 0.16 | 0.11 | |
| 白腹鸫Turdus pallidus | 15.09 | 4.63 | 东北兔Lepus mandshuricus | 0.16 | ||
| 紫啸鸫Myophonus caeruleus | 0.11 | 松鼠Sciurus vulgari | 18.78 | 36.72 | ||
| 白眉地鸫Zoothera sibirica | 0.45 | 花鼠Eutamias sibiricus | 41.73 | 35.60 | ||
| 斑鸫Turdus eunomus | 0.11 | 棕背Clethrionomys rufocanus | 0.48 | 2.03 | ||
| 虎斑地鸫Zoothera dauma | 0.11 | 大林姬鼠Apodemus peninsulae | 0.16 | 0.90 | ||
| 红胁蓝尾鸲Tarsiger cyanurus | 0.11 | 黑线姬鼠Apodemus agrarius Pallas | 0.16 | |||
| 松鸦Garrulus glandarius | 0.16 | 0.23 | ||||
| 花尾榛鸡Bonasia bonasia | 1.61 | 1.13 | ||||
| 白背啄木鸟Dendrocopos leucotos | 0.64 | 0.34 | ||||
| 长尾林鸮Strix uralensis | 0.16 | 0.11 | ||||
| 松雀鹰Accipiter virgatus | 0.16 | |||||
相对丰富度较高的松鼠、花鼠和西伯利亚狍为为较长拍种, 狗獾(Meles meles)、棕背(Clethrionomys rufocanus Sundevall)和白腹鸫为偶拍种, 其他均为罕拍种。
利用红外相机监测兽类的结果中发现监测中夜行性小型啮齿如棕背、大林姬鼠、黑线姬鼠及小型鼬科伶鼬, 监测效果并不理想, 由于这些物种个体小, 以夜间活动为主, 对其监测到的个体很难识别到种;对松鼠、花鼠等体积较大的啮齿类及野猪、西伯利亚狍、黄鼬监测效果较好(表 1—表 3), 所以在利用红外相机对兽类多样性研究中为避免对小型啮齿类动物的评估不足, 结合传统方法能够使得研究结果更为准确。
本次监测的鸟类中既有留鸟, 也有迁徙较晚的候鸟如红胁蓝尾鸲(Tarsiger cyanurus)、白眉地鸫(Zoothera sibirica), 从对鸟类的监测结果看, 主要以林下活动的鸟类为主(表 1—表 3), 所以可以利用红外相机对林下鸟类进行监测研究, 但是想要对保护区内鸟类多样性进行深入研究需结合传统调查方法。
4 讨论2011年以来红外相机技术广泛应用于国内鸟兽多样性研究中。在生物多样性和群落调查中抽样量充分性是研究中值得重点关注的问题[17-18]。通过稀疏化曲线, 可发现保护区内兽类取样比鸟类取样更为充分, 其主要是相机的安装位置对鸟类监测的限制性。本次监测中对鸟兽物种丰富度估计在70%以上, 将本次监测结果与近年相关研究资料对比分析发现, 低纬度(22°—27°)地区监测到的兽类种数较多[19-22], 中纬度(28°—33°)地区次之[13, 23-29], 高纬度地区监测到的兽类种数最少[30-32];秦岭处于古北界和东洋界两大生物区的分界线上, 以秦岭为界, 秦岭以南[20-22, 24-27, 30]要比秦岭以北[30-32]监测到的鸟类种数多(新疆除外), 新疆卡拉麦里山属于荒漠沙漠生境, 与森林生态系统相比, 红外相机在其监测过程中所拍摄范围更广, 可监测到的鸟类数量更多, 对森林生态系统中鸟类进行监测过程中主要获得林下鸟类, 这也是与同纬度地区鸟类监测中产生较大差异的原因。通过在纬度上的对比发现, 鸟兽多样性在纬度上存在梯度变化, 这与张荣祖所研究的结果相似[33]。
物种多样性与生境间呈规律变化[34], 本次调查区域属于针阔混交林, 将已有研究区域根据不同生境类型可划分为针阔混交林[31-32]、落叶阔叶林[23, 29]、常绿落叶阔叶混交林[25]、常绿阔叶林[21-22]、热带雨林[20, 35]和荒漠/沙漠[30], 由于多地鸟类监测未达到饱和[29, 31, 35], 很难发现在不同生境中监测到鸟类种数的变化;在荒漠/沙漠[30]和针阔混交林中[31-32]监测到的兽类种数相较于其他生境少, 这些地区年平均降雨量、年平均气温小于其他地区, 是导致鸟兽多样性相较于其他地区低的主要原因[33]。本次监测与同种生境类型[31-32]下监测到的兽类物种数相近, 但本次调查与北京松山保护区监测到的兽类差距较大, 刘芳等人利用公里网格法对保护区内兽类进行监测, 相机覆盖面积广, 而本研究相机以公顷网格布设, 是产生差距侧主要原因[36]。
凉水、丰林保护区内物种丰富度最高的为松鼠和花鼠, 主要原因为松鼠家域较小, 其次是由于相对于其他啮齿类即使在夜间活动也能被准确的辨认出来。除松鼠和花鼠外很多啮齿类无法准确识别[37-38], 针对这一类群的动物传统的笼捕法更为有效[39-40]。丰富度第二高的兽类为西伯利亚狍, 其主要原因是在凉水、丰林保护区内都有相当可观的西伯利亚狍种群存在。
丰富度较高的啮齿松鼠、花鼠以及普通在森林生态系统中是重要的种子扩散者, 通过对种子的分散贮藏来促进松树的种群更新[41-45]。由于保护区布设于典型红松林中, 所以也是导致这些物种在监测保护区内丰富度较高的原因。
捕获率可以作为野生动物相对丰度的一个指标[12], 研究发现除松鼠和花鼠外其他物种捕获率均不高, 影响捕获率的原因通常有种群数量多少、动物自身家域大小以及最大活动距离。相机镜头的拍摄视角约为40°, 动物个体经过相机视角是随机的, 动物种群越大, 经过相机拍摄视角的概率越高, 拍摄率越高。动物个体家域和最大活动距离越大经过相机视角的概率越小, 反之则拍摄率越大。
致谢: 感谢中国科学院动物研究所肖治术研究员、东北林业大学金光泽教授对工作的支持,感谢丰林保护区宋国华、黄华及东北林业大学张鸣天、宋琪、杨洋、田鹏对野外调查帮助。| [1] | 曹长雷, 高玮, 由玉岩, 王继龙, 周彤, 邓秋香. 次生林斑块的边缘效应对鸟类分布格局的影响. 四川师范大学学报:自然科学版, 2010, 33(2): 247–250. |
| [2] | Parsons A W, Bland C, Forrester T, Baker-Whatton M C, Schuttler S G, McShea W J, Costello R, Kays R. The ecological impact of humans and dogs on wildlife in protected areas in eastern North America. Biological Conservation, 2016, 203: 75–88. DOI:10.1016/j.biocon.2016.09.001 |
| [3] | Brodie J F, Giordano A J, Ambu L. Differential responses of large mammals to logging and edge effects. Mammalian Biology-Zeitschrift für Säugetierkunde, 2015, 80(1): 7–13. DOI:10.1016/j.mambio.2014.06.001 |
| [4] | Morrison J C, SechrestW, Dinerstein E, Wilcove D S, Lamoreux J F. Persistence of large mammal faunas as indicators of global human impacts. Journal of Mammalogy, 2007, 88(6): 1363–1380. DOI:10.1644/06-MAMM-A-124R2.1 |
| [5] | Muldoon K M, Goodman S M. Primates as predictors of mammal community diversity in the forest ecosystems of Madagascar. PLoS One, 2015, 10(9): e0136787. DOI:10.1371/journal.pone.0136787 |
| [6] | 王斌, 彭波涌, 李晶晶, 普穷, 胡慧建, 马建章. 西藏珠穆朗玛峰国家级自然保护区鸟类群落结构与多样性. 生态学报, 2013, 33(10): 3056–3064. |
| [7] | Stephens P A, Zaumyslova O Y, Miquelle D G, Myslenkov A I, Hayward G D. Estimating population density from indirect sign:track counts and the Formozov-Malyshev-Pereleshin formula. Animal Conservation, 2006, 9(3): 339–348. DOI:10.1111/acv.2006.9.issue-3 |
| [8] | 梁虎. 长白山保护区地区东北虎及其猎物群落生境适宜性评价[D].北京:清华大学, 2013. http://cdmd.cnki.com.cn/Article/CDMD-10003-1016712472.htm |
| [9] | 张常智, 张明海. 黑龙江省东完达山地区东北虎猎物种群现状及动态趋势. 生态学报, 2011, 31(21): 6481–6487. |
| [10] | 张家银, 廖进平, 罗建峰, 陈卫新, 周襄武, 康华靖. 浙江九龙山陆生野生脊椎动物资源现状及保护对策. 温州大学学报:自然科学版, 2009, 30(4): 1–6. |
| [11] | Cutler T L, Swann D E. Using remote photography in wildlife ecology:a review. Wildlife Society Bulletin, 1999, 27(3): 571–581. |
| [12] | Kinnaird M F, O'Brien T G. Effects of private-land use, livestock management, and human tolerance on diversity, distribution, and abundance of large african mammals. Conservation Biology the Journal of the Society for Conservation Biology, 2012, 26(6): 1026. DOI:10.1111/j.1523-1739.2012.01942.x |
| [13] | Ahumada J A, Silva C E F, Gajapersad K. Community structure and diversity of tropical forest mammals:data from a global camera trap network. Philosophical Transactions of the Royal Society B:Biological Sciences, 2011, 366(1578): 2703–2711. DOI:10.1098/rstb.2011.0115 |
| [14] | Liu X H, Wu P F, Songer M, Cai Q, He X B, Zhu Y, Shao X M. Monitoring wildlife abundance and diversity with infra-red camera traps in Guanyinshan Nature Reserve of Shaanxi Province, China. Ecological Indicators, 2013, 33: 121–12. DOI:10.1016/j.ecolind.2012.09.022 |
| [15] | 马鸣, 徐峰, Chundawat R, Jumabay K, 吴逸群, 艾则孜, 朱玛洪. 利用自动照相术获得天山雪豹拍摄率与个体数量. 动物学报, 2006, 52(4): 788–793. |
| [16] | 章书声, 鲍毅新, 王艳妮, 方平福, 叶彬. 基于红外相机技术的黑麂活动节律. 兽类学报, 2012, 32(4): 368–372. |
| [17] | 李巧, 陈又清, 徐正会. 蚂蚁群落研究方法. 生态学杂志, 2009, 28(9): 1862–1870. |
| [18] | 陈又清, 李巧, 陈彦林, 王思铭. 糖醋液和乙二醇对地表甲虫的诱集效率比较. 昆虫知识, 2010, 47(1): 129–133. DOI:10.7679/j.issn.2095-1353.2010.023 |
| [19] | 蔡玉生, 龚粤宁, 卢学理, 肖治术, 王新财. 南岭森林哺乳动物多样性的红外相机监测. 生态科学, 2016, 35(2): 57–61. |
| [20] | 施泽攀, 周岐海, 李先琨, 肖治术. 弄岗森林动态监测样地及周边鸟兽的红外相机初步监测. 生物多样性, 2014, 22(6): 826–829. |
| [21] | 吴建普, 罗红, 朱雪林, 李炳章, 刘务林, 次平. 西藏墨脱不同海拔区鸟兽红外相机监测. 生物多样性, 2016, 24(3): 351–354. DOI:10.17520/biods.2015106 |
| [22] | 温立嘉, 时坤, 黄建, 宋阳, 郭玉民. 西藏墨脱鸟兽红外相机监测初报. 生物多样性, 2014, 22(6): 798–799. |
| [23] | 武鹏峰, 刘雪华, 蔡琼, 何祥博, Songer M, 朱云, 邵小明. 红外相机技术在陕西观音山自然保护区兽类监测研究中的应用. 兽类学报, 2012, 32(1): 67–71. |
| [24] | 斯幸峰, 丁平. 古田山森林动态监测样地内鸟兽种群动态的红外相机监测. 生物多样性, 2014, 22(6): 819–822. |
| [25] | 刘芳, 宿秀江, 李迪强, 王本忠, 张自亮. 利用红外相机调查湖南高望界国家级自然保护区鸟兽多样性. 生物多样性, 2014, 22(6): 779–784. |
| [26] | 郑伟成, 章书声, 潘成椿, 刘菊莲, 季国华. 红外相机技术监测九龙山国家级自然保护区鸟兽多样性. 浙江林业科技, 2014, 34(1): 17–22. |
| [27] | 李广良. 神农架保护区川金丝猴活动区域动植物群落研究[D]. 北京:中国林业科学研究院, 2012. http://cdmd.cnki.com.cn/Article/CDMD-82201-1012423514.htm |
| [28] | 李晟, 张晓峰, 陈鹏, 王军, 向定乾, 董伟, 张希明, 何百锁, 孙瑞谦, 赵纳勋, 王大军. 秦岭南坡森林有蹄类群落组成与垂直分布特征. 动物学杂志, 2014, 49(5): 633–643. |
| [29] | 肖治术, 杜晓军, 王学志, 黄小群, 闫满玉. 利用红外相机对河南宝天曼森林动态监测样地鸟兽的初步调查. 生物多样性, 2014, 22(6): 813–815. |
| [30] | 董潭成, 初红军, 吴洪潘, 王渊, 葛炎, 布兰. 卡拉麦里山有蹄类自然保护区鸟兽的红外相机监测. 生物多样性, 2014, 22(6): 804–807. |
| [31] | 李欣海, 朴正吉, 武耀祥, 周海成, 张鹏, 刘丙万, 黄小群, 肖治术. 长白山森林动态监测样地鸟兽的红外相机初步监测. 生物多样性, 2014, 22(6): 810–812. |
| [32] | 张鹏, 姜广顺, 肖治术. 运用红外相机和样线法调查黑龙江双河自然保护区兽类资源. 生物多样性, 2014, 22(6): 773–775. |
| [33] | 张荣祖, 林永烈. 中国及其邻近地区兽类分布的趋势. 动物学报, 1985, 31(2): 187–197. |
| [34] | MacArthur R, Recher H, Cody M. On the relation between habitat selection and species diversity. The American Naturalist, 1966, 100(913): 319–332. DOI:10.1086/282425 |
| [35] | 张明霞, 曹林, 权锐昌, 肖治术, 杨小飞, 张文富, 王学志, 邓晓保. 利用红外相机监测西双版纳森林动态样地的野生动物多样性. 生物多样性, 2014, 22(6): 830–832. |
| [36] | 刘芳, 李迪强, 吴记贵. 利用红外相机调查北京松山国家级自然保护区的野生动物物种. 生态学报, 2012, 32(3): 730–739. |
| [37] | 李欣海, 于家捷, 张鹏, 朴正吉, 肖治术. 应用红外相机监测结果估计小型啮齿类物种的种群密度. 生态学报, 2016, 36(8): 2311–2318. |
| [38] | 章书声, 鲍毅新, 王艳妮, 方平福, 叶彬. 红外相机技术在鼠类密度估算中的应用. 生态学报, 2013, 33(10): 3241–3247. |
| [39] | Smith M H, Gardner R H, Gentry J B, Kaufman D W, O'Farrel M H. Density estimation of small mammal populations//Golley F B, Petrusewicz K, Ryszkowski L, eds. Small Mammals:Their Productivity and Population Dynamics. Cambridge:Cambridge University Press, 1975:25-53. |
| [40] | Krebs C J, Boonstra R, Gilbert S, Reid D, Kenney A J, Hofer E J. Density estimation for small mammals from livetrapping grids:rodents in northern Canada. Journal of Mammalogy, 2011, 92(5): 974–981. DOI:10.1644/10-MAMM-A-313.1 |
| [41] | 姚晓琳, 朴正吉, 李步杭, 张健, 王绪高, 叶吉, 郝占庆. 啮齿动物和鸟类对红松种子的消耗. 应用生态学报, 2008, 19(8): 1759–1763. |
| [42] | Hutchins H E, Hutchins S A, Liu B W. The role of birds and mammals in Korean pine (Pinus koraiensis) regeneration dynamics. Oecologia, 1996, 107(1): 120–130. DOI:10.1007/BF00582242 |
| [43] | Bobadilla S Y, Benitez V V, Guichón M L. Asiatic Callosciurus squirrels as seed dispersers of exotic plants in the Pampas. Current Zoology, 2016, 62(3): 215–219. DOI:10.1093/cz/zow009 |
| [44] | Yi X F, Li J, Zhang M M, Zhang D Y, Wang Z Y. Short-term acute nitrogen deposition alters the interaction between Korean pine seeds and food hoarding rodents. Forest Ecology and Management, 2016, 367: 80–85. DOI:10.1016/j.foreco.2016.02.025 |
| [45] | Yang Y Q, Zhang M M, Yi X F. Small rodents trading off forest gaps for scatter-hoarding differs between seed species. Forest Ecology and Management, 2016, 379: 226–231. DOI:10.1016/j.foreco.2016.08.005 |