 2018, Vol. 38
2018, Vol. 38文章信息
- 杨晓艳, 张世雄, 温静, 徐满厚.
- YANG Xiaoyan, ZHANG Shixiong, WEN Jing, XU Manhou.
- 吕梁山森林群落草本层植物物种多样性的空间格局及其对模拟增温的响应
- Spatial pattern of herbaceous plant species diversity and its changes due to simulated warming in the forest community of the Luliang Mountains
- 生态学报. 2018, 38(18): 6642-6654
- Acta Ecologica Sinica. 2018, 38(18): 6642-6654
- http://dx.doi.org/10.5846/stxb201803300700
-
文章历史
- 收稿日期: 2018-03-30
- 修订日期: 2018-07-02

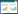




由温室效应不断累积所引起的全球气候变暖正在对陆地生态系统结构和功能等方面产生影响[1]。作为当今生态与环境科学领域研究的热点问题, 生物多样性是促进经济发展、维持生态和谐的基础[2], 而物种多样性是区域多样性的重要内容。因此, 研究气候变暖条件下物种多样性的空间分布具有重要生态意义。
物种的分布是多个生态过程的产物[3], 在不同研究尺度下, 植物物种多样性呈现出不同梯度分布格局[4]。对此, 众学者在国内外不同山地进行了大量研究, 主要以海拔梯度为主[5], 因为海拔梯度能够在相对有限的空间内更好地表征温度、湿度和土壤等诸多生境要素的差异性[6]。在海拔方面, 一般研究表明, 山地植物群落的多样性随海拔梯度的升高呈现“中间高度膨胀”[7-11]和负相关变化[12-14]两种, 而无显著性的分布情况较少[4, 15], 且大部分处于海洋性气候的山地。在纬度方面, 山地植物群落多样性随纬度梯度变化的研究相对较少且存在一定困难, 主要原因在于植被物种多样性指数计算依赖于样方的大小, 而对这方面的系统研究又取决于一定可比较的样方积累[4], 同时, 多样性随纬度的变化会受地形、气候等因素制约。因此, 自然条件下物种多样性的分布格局尚存在较大不确定性, 急需对物种多样性随海拔、纬度的空间变化进行深入探讨。
温度升高可促进植物生长和发育, 从而对植物群落的组成及演替产生一定影响。由于不同物种对温度升高的响应不同, 全球变暖将打破原有种间竞争[16], 并使该群落中优势种产生变化[17-18], 从而使物种趋于单一化发展。目前, 有关植物物种多样性对气候变暖响应的研究多集中于高寒草甸, 且研究结果表明物种多样性在不同增温幅度和增温时间下变化不同[19], 但对于其他区域研究较少。因此, 有必要在不同空间尺度开展物种多样性对气候变暖响应的深入研究。
吕梁山位于黄土高原东部, 呈南北走向, 其海拔、纬度梯度差异较大, 造成植被空间分布格局变化明显。对于吕梁山的研究, 多采用双向指示种分析法、除趋势对应分析等分析方法, 对山地植被类型进行分类和划分[20]以及对山地植物区系地理成分进行分析[21], 并逐渐将海拔梯度的思想引入植物群落的研究[22-23], 但对其植被多样性垂直格局的研究主要针对于单个山体[24-26], 运用系统论的观点和方法对吕梁山植被空间分异特征的分析仅见徐满厚等[27]的研究, 关于吕梁山森林植被物种多样性对增温响应的研究更是尚未见报道[27]。因此, 本研究以吕梁山森林群落草本层植物为研究对象, 进行模拟增温试验, 分析草本层植物物种多样性的空间分布格局及其对模拟增温的响应, 以期为黄土高原物种多样性的保护提供科学依据。
1 研究地区与研究方法 1.1 研究区概况吕梁山, 位于山西省西部(图 1), 南北约400 km, 属半干旱大陆性季风气候, 年均降水量150—400 mm, 且降水多集中于7—9月份, 植物生长期短, 无霜期仅140 d左右[28]。吕梁山纬度跨度较大, 海拔较高, 气候垂直变化及山脉纬度地带特征较明显。独特的气候条件与复杂的地形给物种提供了良好的生境, 使该区动植物资源丰富, 几乎包括了华北地区所有植物品种。

|
| 图 1 样地区位图 Fig. 1 Sample area bitmap |
五鹿山(36.38°—36.63°N、111.03°—111.3°E), 海拔1135—1950 m, 属吕梁山南段、黄土高原东部边缘, 其腹地位于山西省蒲县, 属于温带季风气候, 平均气温8.7℃, 年均降水500—560 mm, 森林覆盖率达60%以上, 水土保持良好, 主要树种为油松(Pinus tabuliformis)和辽东栎(Quercus liaotungensis), 植被类型具有明显的过渡性[25]。在实验地(1318—1602 m), 从低海拔到高海拔草本植物优势种依次为:铁杆蒿(Artemisia gmelinii)和粗壮苔草(Carex heterostachya)。
管涔山(38.25°—39.05°N、111.76°—112.9°E), 海拔1310—2770 m, 属吕梁山北段、鄂尔多斯台地东缘, 其腹地位于山西省宁武县, 属于温带大陆性季风气候[26], 昼夜温差大, 年均降水470—500 mm。植被位于暖温带落叶阔叶林与温带草原交界处, 主要植被为云杉(Picea asperata)、华北落叶松(Larix principis-rupprechtii), 且植被垂直变化明显, 在海拔>2000 m的位置出现以矮嵩草(Kobresia humilis)为建群种的亚高山草甸。在实验地(2222—2571 m), 从低海拔到高海拔草本植物优势种依次为:野草莓( Fragaria orientalis)和粗壮苔草(Carex heterostachya)。
1.2 样地设置试验样地位于吕梁山系的五鹿山(36.50°—36.52°N、111.15°—111.18°E)和管涔山(38.76°—38.79°N、111.94°—111.97°E), 对两座山均设置3个海拔梯度, 即3块试验样地。其中, 在五鹿山选取的样地平均海拔依次为:1318、1459 m和1602 m; 在管涔山选取的样地平均海拔依次为:2222、2395 m和2571 m。选取植被分布均匀、物种较多的林下草本层植物进行样地布设, 各样地面积为35—40 m2, 共计6个样地。分别对每个样地设置对照(CK)、低度增温(OTC1)和高度增温(OTC2)3种处理, 每种处理3个重复, 故各海拔梯度共设9个增温样方, 各样方随机布设, 间距为1—3 m。
增温样地采用无需电力供应的开顶室增温箱(OTC, 图 2)进行模拟增温处理, 箱体由透明有机玻璃构建, 厚度为4 mm, 增温小室为正六边形, 高度分别为40、80 cm, 顶面边长均为25 cm, 底面边长分别为50、75 cm。(1)CK样方设置:不进行增温处理, 使植被处于自然生长状态, 取样面积为1 m×1 m; (2)OTC1样方设置:装置高度为40 cm, 顶面边长25 cm, 底面边长50 cm, 各斜边与地面夹角均为60°, 升温约(1.7±0.3)℃; (3)OTC2样方设置:装置高度为80 cm, 顶面边长25 cm, 底面边长75 cm, 各斜边与地面夹角均为60°, 升温约(3.5±0.5)℃。

|
| 图 2 两种增温梯度装置 Fig. 2 Two temperature increasing gradient devices |
于2016年8月进行吕梁山森林群落考察, 选取林下草本层植物作为试验对象。其中, 五鹿山选取的样地以落叶阔叶树种为主, 从低海拔到高海拔依次分布着灌草丛、人工针叶林和针阔叶混交林; 管涔山选取的样地以针叶树种为主, 从低海拔到高海拔均分布着纯针叶林, 以云杉和华北落叶松为建群种。故本研究所选取的草本层植物均发育在乔木林或灌木林底层。于当年9月份进行样地的设置和增温装置架设。
1.3 植被群落特征测定于2017年8月(此时已进行1年的增温实验)对管涔山、五鹿山各样地森林群落进行植物物种多样性调查, 统计林下草本层植物的种类、多度、频度、盖度、高度, 计算每个物种的重要值, 将各样地不同处理下各物种重要值统计于表 1中。采用0.5 m×0.5 m网格样方框(图 3)进行测定, 根据样方框大小定制网格尺寸为0.1 m×0.1 m的铁丝网, 共计25个网格, 测量时将铁丝网平铺于样方框内, 统计每个物种的名称、数量、频度、盖度和高度。其中, 频度为各物种在网格中出现的次数, 盖度为各物种在网格中所占面积, 高度为各物种处于自然状态下卷尺所测最高值。此外, 本研究将乔木、灌木、藤本植物的幼苗均归为草本层, 各物种名由《中国植物志》得出。
| 山体名称 Mountain name |
海拔梯度 Altitude gradient |
植物种类 Plant species |
重要值Important value/% | ||
| 对照 CK |
低度增温 OTC1 |
高度增温 OTC2 |
|||
| 五鹿山 | 低海拔 | 铁杆蒿Artemisia gmelinii | 33.41 | 28.04 | 51.54 |
| 粗壮苔草Carex heterostachya | 11.27 | 0.00 | 0.00 | ||
| 铁扫帚Lespedeza cuneata | 10.25 | 7.79 | 0.00 | ||
| 野豌豆Vicia sepium | 8.89 | 2.44 | 6.79 | ||
| 苦马豆Sphaerophysa salsula | 8.85 | 10.56 | 11.55 | ||
| 腺毛萎陵菜Potentilla longifolia | 4.85 | 6.74 | 0.00 | ||
| 阴行草Siphonostegia chinensis | 4.78 | 1.96 | 0.00 | ||
| 针茅Stipa capillata | 4.71 | 15.30 | 9.35 | ||
| 堇菜Viola verecumda | 3.33 | 2.55 | 2.15 | ||
| 黄花铁线莲(幼苗)Clematis intricata | 1.68 | 2.00 | 0.00 | ||
| 地构叶Speranskia tuberculata | 1.62 | 0.00 | 0.00 | ||
| 山韭Allium thunbergii | 1.46 | 1.20 | 0.00 | ||
| 狗娃花Heteropappus hispidus | 1.45 | 0.00 | 9.61 | ||
| 远志Polygala tenuifolia | 1.44 | 0.00 | 0.00 | ||
| 黄芩Scutellaria baicalensis | 1.28 | 1.43 | 0.00 | ||
| 刺儿菜Cirsium setosum | 0.70 | 1.16 | 4.25 | ||
| 藠头Allium chinense | 0.00 | 10.18 | 0.00 | ||
| 苜蓿Medicago Sativa | 0.00 | 2.50 | 0.00 | ||
| 芹叶铁线莲(幼苗)Clematis aethusifolia | 0.00 | 2.04 | 0.00 | ||
| 黄刺玫(幼苗)Rosa xanthina | 0.00 | 1.98 | 0.00 | ||
| 山野扁豆Cassia nomame | 0.00 | 1.47 | 0.00 | ||
| 齿苞风毛菊Saussurea odontolepis | 0.00 | 0.66 | 1.45 | ||
| 白头翁Pulsatilla chinensis | 0.00 | 0.00 | 3.31 | ||
| 中海拔 | 粗壮苔草Carex heterostachya | 78.74 | 77.30 | 75.99 | |
| 祁州漏芦Rhaponticum uniflorum | 5.70 | 3.10 | 0.95 | ||
| 艾蒿Artemisia argyi | 4.13 | 0.00 | 0.00 | ||
| 蹄叶橐吾Ligularia fischeri | 4.10 | 0.00 | 0.00 | ||
| 芹叶铁线莲(幼苗)Clematis aethusifolia | 5.41 | 0.00 | 0.00 | ||
| 南牡蒿Artemisia eriopoda | 1.93 | 0.00 | 0.00 | ||
| 石沙参Adenophora polyantha | 0.00 | 10.61 | 8.58 | ||
| 黄刺玫(幼苗)Rosa xanthina | 0.00 | 5.34 | 1.01 | ||
| 草木犀Melilotus officinalis | 0.00 | 2.48 | 0.00 | ||
| 三角枫(幼苗)Acer buergerianum | 0.00 | 1.17 | 0.00 | ||
| 灰条菜Chenopodium album | 0.00 | 0.00 | 7.52 | ||
| 小叶梣(幼苗)Fraxinus bungeana | 0.00 | 0.00 | 5.95 | ||
| 高海拔 | 粗壮苔草Carex heterostachya | 81.16 | 95.62 | 100.00 | |
| 知风草Eragrostis ferruginea | 13.28 | 0.00 | 0.00 | ||
| 龙芽草Agrimonia pilosa | 5.56 | 0.00 | 0.00 | ||
| 辽东栎(幼苗)Quercus wutaishanica | 0.00 | 4.38 | 0.00 | ||
| 管涔山 | 低海拔 | 野草莓Fragaria vesca | 35.38 | 18.94 | 22.42 |
| 粗壮苔草Carex heterostachya | 18.75 | 19.45 | 18.64 | ||
| 路边青Geum aleppicum | 7.90 | 4.72 | 0.00 | ||
| 四叶葎Galium bungei | 7.46 | 12.89 | 11.11 | ||
| 披碱草Elymus dahuricus | 6.38 | 9.53 | 0.00 | ||
| 卷耳Cerastium arvense | 4.66 | 2.95 | 0.63 | ||
| 刺儿菜Cirsium setosum | 4.18 | 0.00 | 4.37 | ||
| 毛蕊老鹳草Geranium eriostemon | 3.32 | 0.00 | 1.04 | ||
| 乌头Aconitum carmichaelii | 2.89 | 7.00 | 3.67 | ||
| 歪头菜Vicia unijuga | 2.14 | 1.73 | 1.80 | ||
| 马先蒿Pedicularis resupinata | 1.90 | 5.32 | 5.77 | ||
| 甘露子Stachys sieboldii | 1.84 | 1.51 | 0.00 | ||
| 珠芽蓼Polygonum viviparum | 1.83 | 8.28 | 4.91 | ||
| 地榆Sanguisorba officinalis | 0.90 | 3.70 | 1.43 | ||
| 半夏Pinellia ternata | 0.47 | 0.00 | 2.33 | ||
| 柳叶芹Czernaevia laevigata | 0.00 | 3.97 | 4.81 | ||
| 路边青Geum aleppicum | 0.00 | 0.00 | 12.43 | ||
| 酸模Rumex acetosa | 0.00 | 0.00 | 2.07 | ||
| 早熟禾Poa annua | 0.00 | 0.00 | 1.99 | ||
| 腺毛委陵菜Potentilla longifolia | 0.00 | 0.00 | 0.58 | ||
| 中海拔 | 粗壮苔草Carex heterostachya | 69.95 | 55.84 | 65.79 | |
| 半夏Pinellia ternata | 10.40 | 13.17 | 7.70 | ||
| 金线草Rubia membranacea | 9.16 | 12.80 | 13.02 | ||
| 毛蕊老鹳草Geranium eriostemon | 6.14 | 9.97 | 7.19 | ||
| 野草莓Fragaria vesca | 3.23 | 0.00 | 0.00 | ||
| 花荵Polemonium coeruleum | 1.13 | 5.84 | 0.00 | ||
| 酸模Rumex acetosa | 0.00 | 2.39 | 0.00 | ||
| 柳叶芹Czernaevia laeviealta | 0.00 | 0.00 | 4.09 | ||
| 四叶葎Galium bungei | 0.00 | 0.00 | 0.96 | ||
| 华北落叶松(幼苗)Larix principis-rupprechtii | 0.00 | 0.00 | 1.25 | ||
| 高海拔 | 粗壮苔草Carex heterostachya | 33.51 | 25.13 | 16.31 | |
| 火焰草Sedum stellariifolium | 26.84 | 25.63 | 31.05 | ||
| 野草莓Fragaria vesca | 16.67 | 2.37 | 22.96 | ||
| 卷耳Cerastium arvense | 7.63 | 22.25 | 15.23 | ||
| 路边青Geum aleppicum | 4.50 | 0.00 | 0.00 | ||
| 早熟禾Poa annua | 3.85 | 9.27 | 9.09 | ||
| 堇菜Viola verecumda | 2.70 | 0.00 | 1.37 | ||
| 珠芽蓼Polygonum viviparum | 2.42 | 6.45 | 1.30 | ||
| 腺毛委陵菜Potentilla longifolia | 1.88 | 3.27 | 0.00 | ||
| 柳叶芹Czernaevia laeviealta | 0.00 | 4.54 | 0.00 | ||
| 唐松草Thalictrum aquilegifolium | 0.00 | 1.09 | 0.00 | ||
| 毛蕊老鹳草Geranium eriostemon | 0.00 | 0.00 | 2.68 | ||

|
| 图 3 试验样地中的样方框及硬铁丝网 Fig. 3 Quadrat frame and hard wire net in experimental plots |
于2017年8月, 在距地面20 cm高度处架设带有防辐射罩(RS3-B)的空气温度探头(S-THB-M008)对空气温度进行测定, 并采用数据采集器(H21-USB)以30 s为时间间隔存储数据。所有仪器均购置于美国ONSET公司生产的HOBO系列。
1.5 数据处理 1.5.1 物种重要值及多样性指数计算采用α多样性指数对吕梁山森林群落草本层植被物种多样性垂直空间格局进行分析, 其指标包括:Simpson指数、Shannon指数、Pielou指数、Patrick指数。计算公式如下:
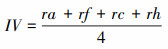
|
(1) |
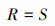
|
(2) |
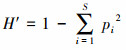
|
(3) |
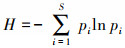
|
(4) |
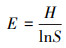
|
(5) |
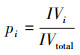
|
(6) |
式中, IV为重要值, ra为相对多度, rf为相对频度, rc为相对盖度, rh为相对高度; H'为Simpson指数; H为Shannon指数; E为Pielou指数; R为Patrick指数; i为样方框内的植物物种i, Pi为第i个物种的相对重要值, S为样方框内所有植物种类的总和。
1.5.2 方差分析对森林群落草本层植物各物种多样性指数(Simpson指数、Shannon指数、Pielou指数、Patrick指数)做不同海拔梯度和不同实验处理的多因素方差分析, 并分别对五鹿山和管涔山森林群落草本层植物的各物种多样性指数在不同增温处理下的变化进行多重比较。
1.5.3 回归分析采用回归分析方法拟合物种多样性随海拔、纬度和温度的回归关系, 依据R2值的大小, 确定拟合程度, 分析海拔梯度、纬度梯度及温度对物种多样性的影响。
1.5.4 分析软件数据初步处理在Microsoft Excel 2013上完成, 用SPSS 19.0软件进行多因素方差分析和多重比较, 用Origin 9.1软件绘制各样地物种多样性指数在不同处理下的变化情况图和多样性指数随海拔、纬度和温度变化的拟合曲线图。
2 结果与分析 2.1 物种多样性对不同海拔梯度的响应物种多样性随海拔呈二次多项式函数变化(图 4), 其中Simpson指数拟合度最高(R2=0.1711), 然后依次为:Pielou指数(R2=0.1303)、Shannon指数(R2=0.1046)、Patrick指数(R2= 0.0515)。总体来看, 各多样性指数随海拔梯度的变化规律一致, 都随海拔的升高多样性先降低后上升, 即在中海拔最低。

|
| 图 4 植被各物种多样性指数随海拔梯度的变化趋势 Fig. 4 Trends of vegetation diversity index with altitude gradient |
物种多样性随纬度升高呈递增趋势, 其中Simpson指数与纬度的拟合程度较高(R2=0.1221, P > 0.05), 依次为Shannon指数(R2=0.0807)、Pielou指数(R2=0.0785)、Patrick指数(R2=0.0382)(图 5)。总体来看, 在较高纬度山地具有较高的物种多样性, 但变化趋势并不明显。

|
| 图 5 植被各物种多样性指数随纬度梯度的变化趋势 Fig. 5 Trends of species diversity index with latitudinal gradient in vegetation |
物种多样性随温度升高呈二次多项式函数变化, 其中Simpson指数的拟合程度最高(R2=0.3374, P > 0.05), 依次为Shannon指数(R2=0.3214)、Pielou指数(R2=0.2684)和Patrick指数(R2=0.2425)(图 6)。总体来看, 随温度升高, 各多样性指数均呈先下降后上升的变化, 整体呈下降趋势(P > 0.05), 其中Simpson指数、Shannon指数和Patrick指数的最低值出现在30℃附近, Pielou指数的最低值出现在25℃附近。

|
| 图 6 植被各物种多样性指数随温度的变化趋势 Fig. 6 Trends of species diversity index with temperature in vegetation |
海拔梯度对林下草本层植物物种多样性的影响呈极显著相关(P < 0.01)(表 2)。在管涔山不同处理对Pielou均匀度指数产生显著影响(P < 0.05), 而在其他样地不同处理对各植物物种多样性的影响均不显著。海拔梯度与不同处理共同作用下对植物物种多样性的影响效果最差。
| 山体名称 Mountain name |
多因素处理 Multi-factor processing |
Simpson多样性指数 Simpson index |
Shannon多样性指数 Shannon index |
Pielou均匀度指数 Pielou index |
Patrick丰富度指数 Patrick index |
| 五鹿山 | 海拔梯度 | 0.000** | 0.000** | 0.000** | 0.000** |
| 不同处理 | 0.271 | 0.146 | 0.197 | 0.082 | |
| 海拔梯度×不同处理 | 0.328 | 0.339 | 0.312 | 0.238 | |
| 管涔山 | 海拔梯度 | 0.000** | 0.000** | 0.000** | 0.000** |
| 不同处理 | 0.089 | 0.131 | 0.012* | 0.504 | |
| 海拔梯度×不同处理 | 0.897 | 0.929 | 0.766 | 0.758 | |
| 表中数据为P值, 若P < 0.05则达到显著水平*, P < 0.01则达到极显著水平** | |||||
在较低纬度的五鹿山, 物种多样性指数在不同处理下均随海拔梯度的升高而下降, 即与海拔呈负相关变化, 且随温度升高多样性受到抑制(图 7)。在低海拔, 持续增温会使物种多样性降低, 且Simpson指数、Shannon指数和Patrick指数在0.05置信水平下显著相关; 在中、高海拔, Simpson指数、Shannon指数、Pielou指数和Patrick指数均随温度升高呈先下降后上升的变化, 但整体呈下降趋势, 其中在高海拔低度增温条件下, Simpson指数、Shannon指数和Pielou指数为0。

|
| 图 7 植被各物种多样性指数在不同处理下各海拔梯度的变化 Fig. 7 Changes of individual altitude gradient in different species of vegetation diversity in different vegetation |
在较高纬度的管涔山, 物种多样性指数在不同处理下均随海拔升高呈现先降低后上升的分布格局, 且适度增温可提高物种多样性, 持续增温多样性则会受到抑制, 但变化趋势并不明显(图 7)。其中, Simpson指数和Pielou均匀度指数在不同海拔梯度下均随温度升高呈现先上升后下降的变化; Shannon指数在低海拔地区与温度呈正相关趋势, 在中、高海拔地区随温度升高多样性指数先升高后降低; Patrick指数在低、中海拔随温度升高呈递增趋势, 在高海拔低度增温处理下达到峰值。
3 讨论 3.1 物种多样性随海拔梯度的变化物种多样性随海拔梯度的变化大致分为5种[4], 即显著负相关、“中间高度膨胀”、中海拔最低、显著正相关、与海拔无关。整体看来, 在吕梁山物种多样性随海拔梯度的升高呈先下降后上升的变化趋势, 符合第3种情况, 即中海拔最低, 与Peet在落基山脉东坡物种多样性随海拔梯度变化的研究结果相似, 这是由两山相似的气候类型和森林郁闭度较低所致[29]。五鹿山物种多样性随海拔梯度升高呈现负相关变化, 符合第1种情况, 这与赵淑清等[13]对长白山植被垂直格局的研究相同, 此外, 在许多山地均呈现该变化特征, 如天山、伏牛山、贡嘎山[12, 14, 30], 该现象是由于随海拔升高温度降低所致; 管涔山物种多样性与随海拔梯度升高也呈现先下降后上升的变化格局, 符合第3种情况。而在青藏高原的研究中, 多样性随海拔梯度的变化以单峰分布格局为主, 符合第2种情况, 即“中间高度膨胀”[9, 31-32]。由于决定物种多样性的根本要素是特定海拔处各环境因子综合作用所形成的综合指标[33], 不同地区由于自然要素存在很大差异, 从而造成多样性随海拔梯度变化的差异性。
3.2 物种多样性随纬度梯度的变化系统地研究植物群落物种多样性随纬度的变化特征有很大难度, 原因主要包括样地的选取与积累[4]。根据欧美以及我国的研究显示, 植物物种多样性呈现明显的纬度变化, 即低纬度物种多样性最高, 随着纬度升高多样性递减[34-35]。但在本文研究中, 位于较高纬度的管涔山物种多样性大于较低纬度的五鹿山。首先是由于所选取的样地纬度跨度较小所致(吕梁山跨度2.67°, 而本研究区跨2.29°), 且在纬向上气温变化率显著低于垂直方向, 因此在小尺度下多样性变化不显著; 其次, 与五鹿山海拔较低, 受历史人为干扰(如耕作、放牧、采药等)有关。吕梁山中段与南段地区人类对其开发历史较久远, 对植被的生长造成较强的干预, 再加上吕梁山区气候干旱, 物种多样性较低, 是黄土高原的生态脆弱区, 人类的活动会严重影响该地区物种多样性, 降低优势种的种群数量与密度[27]。总体看来, 物种多样性随纬度的分布格局受水热条件及人为干扰的影响较大。
3.3 物种多样性随温度升高的变化对于增温幅度方面, 本研究表明, 在五鹿山, 植物物种多样性随温度升高呈递减趋势, 这是由于增温使局部小气候出现暖干化, 从而影响了植被的生长发育; 在管涔山, 适度增温可提高物种多样性, 持续增温多样性则会受到抑制, 这与徐满厚等[36]在青藏高原的研究结果相同, 即适当的增温可极显著促进高寒草甸植被生长, 这是由于在管涔山受地形和海拔等因素影响使该区较高海拔山地表现为严寒微湿的气候[28], 从而与青藏高原的气候类似所致。李娜等[1]的研究也表明, 在高寒草甸, 小幅度增温可使杂草分盖度显著增加(P < 0.05), 大幅增温则抑制增加趋势, 而在沼泽草甸, 小幅增温可使杂草分盖度显著降低, 但大幅增温反而促进这种减少趋势(P < 0.05)。这是由于不同草地类型的土壤条件和水热状况等不同, 导致其植被物种多样性对增温响应的表现不同[1]。
对于增温时间尺度方面, 不同研究区对模拟增温的响应不同。本研究结果表明在吕梁山低海拔地区, 增温1年可显著降低物种多样性, 但在中、高海拔地区, 增温对多样性的影响并不显著。对高寒草甸的研究表明, 短期内适度增温可提高植物物种多样性[37-38], 但这种响应并不敏感, 如徐满厚等[39]在北麓河的模拟增温试验表明, 物种多样性在生长旺季显著高于前期和末期, 且增温3年开始显著性得到加强, 但整体来看, 物种多样性对增温(3年)的响应并不显著。在亚高山草甸, 短期增温并未改变群落组成及物种多样性, 但使群落中优势种种类增加[19]。在东北羊草草甸的研究表明, 长期增温(4年)可提高物种多样性指数[40]。因此, 短期增温对多样性影响效果不显著, 但长期增温会对物种多样性产生显著影响, 主要包括群落组成和结构等方面。
在吕梁山较低海拔, 持续增温会使物种多样性降低(P < 0.05);在中等海拔, 物种多样性随温度升高呈现先下降后上升的变化趋势; 在较高海拔, 适度增温可提高林下草本层植物物种多样性, 持续增温多样性则受到抑制。对于吕梁山植物物种多样性对不同增温处理下空间分布格局的研究尚未见报道, 今后应注意该领域的研究, 更好地为吕梁山森林群落草本层植物物种多样性的保护提供科学依据。
4 结论从北向南按照纬度梯度选择吕梁山系的管涔山和五鹿山为研究区, 在每个山地的高、中、低海拔分别设置对照、低度增温、高度增温试验样地, 以此探讨吕梁山森林群落草本层植物物种多样性的空间格局及其对模拟增温的响应。吕梁山森林群落草本层植物物种多样性随海拔升高呈先下降后上升的变化趋势, 随纬度增加略有上升, 随温度升高整体呈下降趋势, 但后两者均未达到显著水平。在模拟增温条件下, 吕梁山林下草本植物物种多样性对增温的响应随海拔升高先增强后减弱, 但整体呈下降趋势。
| [1] | 李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895–905. |
| [2] | 刘世荣, 蒋有绪, 史作民. 中国暖温带森林生物多样性研究. 北京: 中国科学技术出版社, 1998: 1–2. |
| [3] | 马晓娟.高寒草甸植物种子萌发与幼苗建植机制研究[D].兰州: 兰州大学, 2008. http://cdmd.cnki.com.cn/Article/CDMD-10730-2008163399.htm |
| [4] | 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 1997, 17(1): 91–99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
| [5] | 唐志尧, 方精云. 植物物种多样性的垂直分布格局. 生物多样性, 2004, 12(1): 20–28. DOI:10.3321/j.issn:1005-0094.2004.01.004 |
| [6] | 赵鸣飞, 邢开雄, 王宇航, 王国义, 薛峰, 左婉怡, 康慕谊. 芦芽山寒温性针叶林冠层下植被beta多样性格局及其成因. 生态学报, 2017, 37(10): 3327–3334. |
| [7] | 刘增力, 郑成洋, 方精云. 河北小五台山北坡植物物种多样性的垂直梯度变化. 生物多样性, 2004, 12(1): 137–145. DOI:10.3321/j.issn:1005-0094.2004.01.017 |
| [8] | 茹文明, 张金屯, 张峰, 张贵萍, 刘瑞祥. 历山森林群落物种多样性与群落结构研究. 应用生态学报, 2006, 17(4): 561–566. DOI:10.3321/j.issn:1001-9332.2006.04.001 |
| [9] | 段敏杰, 高清竹, 郭亚奇, 万运帆, 李玉娥, 干珠扎布, 旦久罗布, 韦兰亭, 西饶卓玛. 藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局. 草业科学, 2011, 28(10): 1845–1850. |
| [10] | 井学辉, 臧润国, 丁易, 张炜银, 张新平, 白志强, 郭仲军. 新疆阿尔泰山小东沟北坡植物多样性沿海拔梯度分布格局. 林业科学, 2010, 46(1): 23–28. |
| [11] | 黄甫昭, 丁涛, 李先琨, 郭屹立, 王斌, 向悟生, 文淑均, 李冬兴, 何运林. 弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化. 生态学报, 2016, 36(14): 4509–4517. |
| [12] | 贾宝全, 闫顺, 李国旗, 许英勤. 天山乌鲁木齐河源区高山带植被及其生物多样性初步研究. 干旱区研究, 2002, 19(2): 17–20. |
| [13] | 赵淑清, 方精云, 宗占江, 朱彪, 沈海花. 长白山北坡植物群落组成、结构及物种多样性的垂直分布. 生物多样性, 2004, 12(1): 164–173. DOI:10.3321/j.issn:1005-0094.2004.01.020 |
| [14] | 卢训令, 胡楠, 丁圣彦, 范玉龙, 廖秉华, 翟元杰, 柳静. 伏牛山自然保护区物种多样性分布格局. 生态学报, 2010, 30(21): 5790–5798. |
| [15] | Rey Benayas J M. Patterns of diversity in the strata of boreal montane forest in British Columbia. Journal of Vegetation Science, 1995, 6(1): 95–98. DOI:10.2307/3236260 |
| [16] | 石福孙, 吴宁, 罗鹏. 川西北亚高山草甸植物群落结构及生物量对温度升高的响应. 生态学报, 2008, 28(11): 5286–5293. DOI:10.3321/j.issn:1000-0933.2008.11.010 |
| [17] | Alward R D, Detling J K, Milchunas D G. Grassland vegetation changes and nocturnal global warming. Science, 1999, 283(5399): 229–231. DOI:10.1126/science.283.5399.229 |
| [18] | Pauli H, Gottfried M, Grabherr G. High summits of the Alps in a changing climate: the oldest observation series on high mountain plant diversity in Europe//Walther G R, Burga C A, Edwards P J, eds. "Fingerprints" of Climate Change. Boston, MA: Springer, 2001. |
| [19] | 马丽, 徐满厚, 翟大彤, 贾燕燕. 高寒草甸植被-土壤系统对气候变暖响应的研究进展. 生态学杂志, 2017, 36(6): 1708–1717. |
| [20] | 上官铁梁. 山西主要植被类型及其分布的初步研究. 山西大学学报, 1985(1): 74–84. |
| [21] | 上官铁梁, 张峰, 邱富财. 芦芽山自然保护区种子植物区系地理成分分析. 武汉植物学研究, 1999, 17(4): 323–331. DOI:10.3969/j.issn.2095-0837.1999.04.007 |
| [22] | 赵鸣飞, 薛峰, 王宇航, 王国义, 邢开雄, 康慕谊, 王菁兰. 山西芦芽山针叶林草本层群落谱系结构与多样性的海拔格局. 植物生态学报, 2017, 41(7): 707–715. |
| [23] | 张文涛, 江源, 王明昌, 张凌楠, 董满宇. 芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应. 生态学报, 2015, 35(19): 6481–6488. |
| [24] | 陈廷贵, 张金屯, 上官铁梁, 张峰, 王巧仙. 山西关帝山神尾沟植物群落多样性研究. 西北植物学报, 2000, 20(4): 638–646. DOI:10.3321/j.issn:1000-4025.2000.04.023 |
| [25] | 曲波, 苗艳明, 张钦弟, 毕润成. 山西五鹿山植物物种多样性及其海拔梯度格局. 植物分类与资源学报, 2012, 34(4): 376–382. |
| [26] | 马丽. 管涔山生物多样性特征研究. 山西林业科技, 2014, 43(3): 19–21. DOI:10.3969/j.issn.1007-726X.2014.03.006 |
| [27] | 徐满厚, 马丽, 白皓宇, 冀钦, 李晓丽. 山西吕梁山植被群落多样性的垂直空间分异. 江苏农业科学, 2017, 45(12): 256–260. |
| [28] | 张金屯, 柴宝峰, 邱扬, 陈廷贵. 晋西吕梁山严村流域撂荒地植物群落演替中的物种多样性变化. 生物多样性, 2000, 8(4): 378–384. DOI:10.3321/j.issn:1005-0094.2000.04.004 |
| [29] | Peet R K. Forest vegetation of the Colorado Front Range:patterns of species diversity. Vegetatio, 1978, 37(2): 65–78. DOI:10.1007/BF00126830 |
| [30] | 沈泽昊, 刘增力, 伍杰. 贡嘎山东坡植物区系的垂直分布格局. 生物多样性, 2004, 12(1): 89–98. DOI:10.3321/j.issn:1005-0094.2004.01.011 |
| [31] | 刘哲, 李奇, 陈懂懂, 翟文婷, 赵亮, 徐世晓, 赵新全. 青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响. 生物多样性, 2015, 23(4): 451–462. |
| [32] | 周华坤, 赵新全, 唐艳鸿, 周立, 刘伟, 于龙. 长期放牧对青藏高原高寒灌丛植被的影响. 中国草地, 2004, 26(6): 1–11. |
| [33] | 王长庭, 龙瑞军, 曹广民, 王启兰, 景增春, 施建军. 高寒草甸不同类型草地土壤养分与物种多样性——生产力关系. 土壤通报, 2008, 39(1): 1–8. DOI:10.3321/j.issn:0564-3945.2008.01.001 |
| [34] | Malyshev L, Nimis P L, Bolognini G. Essays on the modelling of spatial floristic diversity in Europe:British Isles, West Germany, and East Europe. Flora, 1994, 189(1): 79–88. DOI:10.1016/S0367-2530(17)30571-6 |
| [35] | 谢晋阳, 陈灵芝. 暖温带落叶阔叶林的物种多样性特征. 生态学报, 1994, 14(4): 337–344. DOI:10.3321/j.issn:1000-0933.1994.04.012 |
| [36] | 陈磊, 顾润源, 姜学恭, 周伟灿, 宋桂英. 内蒙古中西部一次强沙尘暴天气过程的诊断分析Ⅰ——背景场分析. 干旱区资源与环境, 2013, 27(3): 137–141. |
| [37] | 赵新全, 陈世龙, 曹广民, 师生波, 徐世晓, 王启兰. 青藏高原高寒草甸生态系统与全球气候变化的相互作用机理研究. 科技和产业, 2003, 3(8): 51–59. DOI:10.3969/j.issn.1671-1807.2003.08.004 |
| [38] | 权国玲, 尚占环. 中国草地生态系统模拟增温实验的综合比较. 生态学杂志, 2015, 34(4): 1166–1173. |
| [39] | 徐满厚, 刘敏, 薛娴, 翟大彤, 彭飞, 尤全刚, 刘忠权. 增温、刈割对高寒草甸植被物种多样性和地下生物量的影响. 生态学杂志, 2015, 34(9): 2432–2439. |
| [40] | 高嵩.增温和氮素添加对松嫩草原羊草群落结构和功能的影响[D].长春: 东北师范大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10200-1013142140.htm |