 2018, Vol. 38
2018, Vol. 38文章信息
- 张世雄, 杨晓艳, 温静, 徐满厚.
- ZHANG Shixiong, YANG Xiaoyan, WEN Jing, XU Manhou.
- 山西吕梁山亚高山草甸物种多样性的时空变化格局
- Spatial and temporal patterns of species diversity of subalpine meadows in the Lvliang Mountains, Shanxi Province
- 生态学报. 2018, 38(18): 6685-6693
- Acta Ecologica Sinica. 2018, 38(18): 6685-6693
- http://dx.doi.org/10.5846/stxb201803300667
-
文章历史
- 收稿日期: 2018-03-30
- 修订日期: 2018-07-02

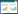




生物多样性研究一直是生态学和全球变化研究的热点问题, 其中物种多样性是最基本的层次, 要解决生物多样性保护、恢复和持续利用问题还须与物种多样性密切结合[1]。物种多样性既体现了生物之间及其与环境之间的复杂关系, 又体现了生物资源的丰富性[2-3]。物种多样性的变化研究一直是生态学的重点问题, 一方面主要集中在物种多样性的纬度、海拔等梯度的空间变化研究, 如贺金生等[4-5]对陆地植物群落物种多样性的纬度梯度变化总结后得出:随着纬度的降低, 群落物种多样性逐渐增加; 对东灵山亚高山草甸物种多样性研究发现, 物种多样性指数会随着海拔的升高发生明显分异[6]; 对云顶山和五台山亚高山草甸的研究显示, 坡向对该地区草甸物种多样性的影响较大[7-8]。另一方面是对物种多样性的时间变化影响因素的分析研究, 如沙威等[9]研究发现植物群落物种多样性指数的变化与土壤含水量呈极显著正相关; 白晓航等[10-12]对河北小五台山的研究发现降水、光照和温度的复合作用以及海拔、坡位、土壤理化性质等环境因子对植物的多样性和植被群落的分布格局有着较强的影响作用; 周红章等[13]分析了物种多样性在不同时间尺度的变化格局, 发现在较短的时间范围内物种多样性变化呈现特殊的模式。可见, 植物群落不同尺度的物种多样性变化对环境的响应不尽一致, 不同物种由于受到各种因子的制约也呈现出不同的变化格局。
亚高山草甸是我国分布范围广泛且以多年生草本植物为主的草甸类型, 由于其生态环境脆弱, 极易受到气候变化及人类活动的影响, 近年来一些地区亚高山草甸已经出现物种多样性下降的情况[14]。吕梁山位于黄土高原东部地区, 为山西省西部山地, 其自然环境恶劣, 植被稀少, 沟壑纵横, 水土流失严重, 其森林覆盖率仅为18.5%, 是重要的生态功能区。在该山系的较高海拔山地分布有大面积的亚高山草甸, 其种类组成与周围低海拔地区的相似性较低, 遭到破坏后极易产生退化, 物种多样性在短时期内将难以恢复。
本研究通过对α多样性指数以及物种重要值的分析, 按照纬度梯度从北向南依次选取吕梁山系的马伦草原、荷叶坪和云顶山作为研究区域, 通过野外实地取样获取的第一手数据资料, 对吕梁山亚高山草甸物种多样性时空变化格局进行研究, 以期能够在一定程度上揭示吕梁山亚高山草甸物种多样性的随时间和纬度的动态变化规律。因此, 研究吕梁山亚高山草甸物种多样性的时空变化格局对该地区草甸植被的有效保护、治理恢复以及合理开发, 促进地区经济社会发展与人口资源环境相协调具有积极作用和重要意义[15-16]。
1 材料与方法 1.1 研究区概况马伦草原位于山西省忻州市宁武县的管涔山顶, 年均气温为6—7℃, 年均降水为400—450 mm, 属温带大陆性气候; 此处植被垂直分布显著:海拔1300—1600 m为灌木丛及农垦带, 海拔1500—1800 m为针阔叶混交林带, 海拔1700—2600 m为针叶林带, 海拔2450—2710 m植被为亚高山草甸带; 该山地的取样点(111.92°E, 38.75°N)位于海拔2710 m, 植被是典型的亚高山草甸。荷叶坪位于山西省忻州市五寨县, 是华北最大的亚高山草甸区, 年均气温为6—10℃, 年均降水为380—680 mm, 属于温带大陆性季风气候; 植被类型具有明显的垂直分布, 与马伦草原同属于管涔山; 该山地取样点(111.54°E, 38.71°N)位于海拔2745 m, 为典型的亚高山草甸带。云顶山位于山西省吕梁市交城县、方山县、娄烦县的交界处, 年均气温为3—4℃, 年均降水量为600—800 mm, 属于温带大陆性季风气候; 植被类型从山麓到山顶依次为, 海拔200—1750 m为落叶阔叶林带, 海拔1750—2200 m为针阔叶混交林带, 海拔2200—2600 m为寒温性针叶林带, 海拔2600—2720 m为亚高山灌丛草甸带; 该山地取样点(111.54°E, 37.88°N)位于海拔2690 m, 为典型的亚高山草甸。本研究区一共3个取样点, 其样地分布如图 1所示。

|
| 图 1 实验样地布局图 Fig. 1 Layout of the sample plots |
于2015年6月中旬在吕梁山对亚高山草甸实验样地进行踏查和选取, 最终选取实验样地由北向南依次为较高纬度的马伦草原、中间纬度的荷叶坪、较低纬度的云顶山, 分别为吕梁山系的偏北部山地、中部山地、偏南部山地, 选择地势平坦、人为干扰少、植被分布均匀的典型亚高山草甸作为实验样地。于2015、2016、2017年每年的植被生长旺季(7—8月)进行一次植被调查, 且样方非固定, 每年的调查样方在同一调查区域重新进行, 随机布设5—6个面积为1 m2的样方进行群落多样性调查, 样方设置采用样方法, 即大小为1 m×1 m的样方框(面积为1 m2)。具体采样如下:2015年对马伦草原和云顶山进行采样, 每个样地分别布设5个样方; 2016年对马伦草原、云顶山、荷叶坪进行采样, 每个样地分别布设6个样方; 2017年对马伦草原、云顶山、荷叶坪进行采样, 每个样地分别布设5个样方, 共布设43个样方。样地设置如表 1所示。
| 样地 Sample plots |
纬度 Latitude |
经度 Longitude |
海拔 Altitude/m |
取样时间 Sampling time |
取样总次数 Total numbers |
| 马伦草原 | 38.75°N | 111.92°E | 2710 | 2015、2016、2017 | 16 |
| 荷叶坪 | 38.71°N | 111.84°E | 2745 | 2016、2017 | 11 |
| 云顶山 | 37.88°N | 111.54°E | 2690 | 2015、2016、2017 | 16 |
分别于2015、2016、2017年的7月底到8月上旬对吕梁山的马伦草原、荷叶坪和云顶山3个实验样地进行亚高山草甸的植被生长特征调查, 使用GPS记录下调查位置的地理坐标和海拔高度。对亚高山草甸按种进行调查, 测量时每个物种视其数量取其平均值作为记录数据, 调查的主要指标有种类、数量、高度、频度和盖度, 其中高度的测量工具为3 m长的钢卷尺, 频度和盖度的测量工具为自制的1 m×1 m铝制样方框图 2 [17]所示(内部分100个10 cm×10 cm小格)。调查过程中如果遇到陌生物种, 用标本夹采样带回实验室, 查阅植物检索表或由从事植物分类学的专家进行鉴定, 亚高山草甸的不同物种的生活型(一年生、两年生、多年生草本)和所属科属的辨认参考中国在线植物志(http://www.eflora.cn/)。

|
| 图 2 实验样地中的样方框及硬铁丝网 Fig. 2 Quadrat frame and hard wire net in experimental plots |
植被调查数据采用α多样性指数主要包括Simpson指数、Shannon-Wiener指数和Pielou均匀度指数, 同时结合Patrick丰富度指数, 对马伦草原、荷叶坪、云顶山连续3年的植被调查数据进行统计分析。在基础数据处理中, 将每个山地的多个亚高山草甸样方进行合并, 同一物种在不同样方中取平均后作为每个山地植被物种的统计数据。在物种多样性指数计算中, 3个山地的亚高山草甸各只有一组相应的群落多样性指数数据。对各样方的数据进行合并处理, 是为了减小在植被调查时的人工取样误差, 防止物种丢失, 从而保证实验数据和结果的准确性[18]。其计算过程为:根据群落调查指标(高度、多度、盖度、频度), 先计算每个物种相应指标的相对值(相对高度、相对多度、相对盖度、相对频度), 由相对值计算每个物种的重要值, 再由重要值得到群落的物种多样性指数。气象数据采用各研究各样地5—8月(生长季)的月均温和降水天数, 从由山西省气象局(http://www.sxsqxj.gov.cn/)以及各样地所在地区的气象局(忻州市和吕梁市)获取相关气象数据并进行相应的计算处理使用。计算公式如下:
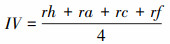
|
(1) |
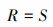
|
(2) |
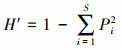
|
(3) |
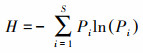
|
(4) |
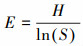
|
(5) |
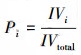
|
(6) |
式中, IV为重要值, rh为相对高度, ra为相对多度, rc为相对盖度, rf为相对频度; R为Patrick丰富度指数; H'为Simpson指数; H为Shonnon-Wiener指数; E为Pielou均匀度指数; i为样方框内的植物物种i, S为样地内所有植物种类的总和, Pi为相对重要值。
使用Excel进行数据计算, 使用Origin 8.5软件对物种多样性进行时间和空间变化的分析和相关图形的绘制。
2 结果与分析 2.1 吕梁山亚高山草甸植物的重要值在吕梁山亚高山草甸群落中, 菊科、蔷薇科、莎草科、禾本科的植物占总植物数的56%, 并且这4科植物的生活型均为多年生草本。在马伦草原亚高山草甸样点共调查到植物23种, 隶属于9科18属。在科水平上, 植物主要集中在菊科、莎草科、蓼科(Polygonaceae)、毛茛科(Ranunculaceae); 在属水平上, 植物主要集中在马先蒿属(Pedicularis)、唐松草属(Thalictrum)、薹草属(Carex)和委陵菜属(Potentilla)。在荷叶坪亚高山草甸样点共调查到植物28种, 隶属于10科20属。在科水平上, 植物主要集中在菊科、蔷薇科、禾本科、莎草科; 在属水平上, 植物主要集中在委陵菜属、马先蒿属、薹草属和蒿属(Artemisia)。在云顶山亚高山草甸样点共调查到植物40种, 隶属于16科30属。在科水平上, 植物主要集中在菊科、蔷薇科、莎草科; 在属水平上, 植物主要集中在委陵菜属、薹草属、蒿属、风毛菊属(Saussurea)。
由表 2的重要值可知, 在马伦草原亚高山草甸群落中, 矮嵩草和珠芽蓼的重要值达到20%以上, 且两个物种的重要值之和>30%, 其相对高度、相对盖度、相对多度和相对频度均高于群落内的其他物种, 表明矮嵩草和珠芽蓼为马伦草原亚高山草甸植物群落的优势种。苔草(Carex montis-wutaii)和披碱草的重要值达10%以上, 为马伦草原草甸群落的次优势种。在荷叶坪亚高山草甸群落中, 线叶嵩草和珠芽蓼为优势种, 唐松草(Thalictrum)和苔草为次优势种。在云顶山亚高山草甸植物群落中, 车前草和披碱草为优势种, 矮嵩草和苔草为次优势种。
| 编号 Number |
科 Family |
物种 Species name |
生活型 Life form |
重要值 Importance value/% |
| 马伦草原 | ||||
| 1 | 莎草科 | 矮嵩草Kobresia humilis | 多年生 | 23 |
| 2 | 蓼科 | 珠芽蓼Polygonum viviparum | 多年生 | 20 |
| 3 | 莎草科 | 苔草Carex montis\\wutaii | 多年生 | 12 |
| 4 | 禾本科 | 披碱草Elymus dahuricus | 多年生 | 11 |
| 5 | 毛茛科 | 毛茛Ranunculus japonicus | 多年生 | 4 |
| 6 | 毛茛科 | 唐松草Thalictrum | 多年生 | 3 |
| 7 | 菊科 | 风毛菊Saussurea nivea | 二年生 | 3 |
| 8 | 菊科 | 蒲公英Taraxacum mongolicum | 多年生 | 3 |
| 9 | 蔷薇科 | 委陵菜Potentilla chinensis | 多年生 | 3 |
| 10 | 菊科 | 火绒草Leontopodium alpinum | 多年生 | 3 |
| 11 | 菊科 | 紫菀Aster tataricus | 多年生 | 2 |
| 荷叶坪 | ||||
| 1 | 莎草科 | 线叶嵩草Kobresia capillifolia | 多年生 | 14 |
| 2 | 蓼科 | 珠芽蓼Polygonum viviparum | 多年生 | 14 |
| 3 | 毛茛科 | 唐松草Thalictrum | 多年生 | 10 |
| 4 | 莎草科 | 苔草Carex montis\\wutaii | 多年生 | 9 |
| 5 | 禾本科 | 披碱草Elymus dahuricus | 多年生 | 7 |
| 6 | 蔷薇科 | 莓叶委陵菜Potentilla fragarioides | 多年生 | 3 |
| 7 | 菊科 | 风毛菊Saussurea nivea | 二年生 | 3 |
| 8 | 菊科 | 火绒草Leontopodium alpinum | 多年生 | 2 |
| 9 | 蔷薇科 | 地榆Sanguisorba officinalis | 多年生 | 2 |
| 10 | 玄参科 | 马先蒿Pedicularis | 多年生 | 2 |
| 11 | 鸭跖草科 | 节节草Commelina diffusa | 一年生 | 2 |
| 12 | 菊科 | 紫菀Aster tataricus | 多年生 | 2 |
| 13 | 车前科 | 车前草Plantago asiatica | 二年或多年生 | 2 |
| 云顶山 | ||||
| 1 | 车前科 | 车前草Plantago asiatica | 二年或多年生 | 15 |
| 2 | 禾本科 | 披碱草Elymus dahuricus | 多年生 | 13 |
| 3 | 莎草科 | 矮嵩草Kobresia humilis | 多年生 | 12 |
| 4 | 莎草科 | 苔草Carex montis\\wutaii | 多年生 | 12 |
| 5 | 伞形科 | 葛缕子Carum carvi | 多年生 | 11 |
| 6 | 蓼科 | 珠芽蓼Polygonum viviparum | 多年生 | 11 |
| 7 | 菊科 | 蒲公英Taraxacum mongolicum | 多年生 | 7 |
| 8 | 菊科 | 火绒草Leontopodium japonicum | 多年生 | 7 |
| 9 | 菊科 | 蒿属Artemisia | 多年生 | 5 |
| 10 | 蔷薇科 | 叉叶委陵菜Potentilla bifurca | 多年生 | 3 |
| 11 | 菊科 | 紫菀Aster tataricus | 多年生 | 2 |
| 12 | 蔷薇科 | 地榆Sanguisorba officinalis | 多年生 | 2 |
| 13 | 蔷薇科 | 腺粒委陵菜Potentilla granulosa | 多年生 | 2 |
因此, 在吕梁山亚高山草甸群落中, 不同山地草甸植物的优势种和次优势种各不相同, 但其均以菊科、蔷薇科、莎草科、禾本科植物为主, 且其生活型均以多年生草本为主。
2.2 吕梁山亚高山草甸物种多样性的时间变化在马伦草原亚高山草甸物种多样性的时间变化中(图 3), Pielou指数表现为随时间增加的趋势, 但变化幅度并不明显; Patrick指数、Simpson指数、Shannon-Wiener指数的变化均表现为先增后减的单峰变化趋势, 且2016年均为最高值。在荷叶坪亚高山草甸物种多样性的时间变化中(图 3), 除Pielou指数在2017年为最高值, Patrick指数、Simpson指数、Shannon-Wiener指数的变化均为递减趋势, 且2016年为最高值。在云顶山物种多样性的时间变化中(图 3), Shannon-Wiener指数和Patrick指数的变化均比较明显, 表现为先增后减的单峰变化趋势, 且2016年为最高值; Simpson指数和Pielou指数在连续3年内的变化趋势并明显变化。在吕梁山亚高山草甸物种多样性时间变化中, Pielou均匀度指数连续3年内无较大变化, 这表明吕梁山亚高山草甸群落的物种竞争性和相对稳定; Shannon-Wiener指数、Simpson指数和Patrick物种丰富度指数在2016年为最高值, 表明吕梁山亚高山草甸群落的物种分布的均匀度, 植被结构的稳定性和物种多样性在2016年最为稳定。

|
| 图 3 各样地不同时间草本植物的物种多样性 Fig. 3 Species diversity of different times in different sample plots |
由气象数据可知(表 3), 在植被生长旺季中, 吕梁山系3个山地的月平均温度在连续3年内的基本保持稳定, 且无极端气候出现, 表明稳定的气温变化并不是对吕梁山亚高山草甸物种多样性时间变化格局的主导因素。在3个山地连续3年的草甸生长季降水天数变化中, 2016年的降水天数明显高于其他两年, 这与吕梁山亚高山草甸物种多样性在2016年高于其他年份相一致, 这表明生长季降水天数是吕梁山物种多样性时间变化的主导因素。因此, 吕梁山亚高山草甸物种多样性在时间上呈现出受降水条件影响较大的变化格局。
| 年份 Year |
马伦草原Malun Grassland | 荷叶坪Heyeping | 云顶山Yunding Mountain | |||||
| 月均温/℃ | 降水天数/天 | 月均温/℃ | 降水天数/天 | 月均温/℃ | 降水天数/天 | |||
| 2015 | 17.5 | 54 | 22.5 | 46 | 17 | 52 | ||
| 2016 | 17.85 | 59 | 22.25 | 64 | 17.5 | 58 | ||
| 2017 | 18.125 | 53 | 23.25 | 53 | 18.5 | 55 | ||
由不同山地亚高山草甸群落物种多样性指数的变化可知(图 4), 在2016年, Patrick指数、Shannon-Wiener指数、Pielou指数的变化趋势均为云顶山>荷叶坪>马伦草原, 荷叶坪的Simpson指数在三地中为最高值。在2017年, 除荷叶坪和云顶山的Patrick指数相同, 在总体上3个山地的Patrick指数、Simpson指数、Shannon-Wiener指数、Pielou指数的变化趋势均为云顶山>荷叶坪>马伦草原。综合两年的变化趋势, 云顶山亚高山草甸群落在物种丰富度、均匀度以及多样性上均是3地中最高值, 这表明吕梁山系亚高山草甸物种多样性变化趋势表现为偏北山地<中部山地<偏南山地。

|
| 图 4 2016、2017年不同样地物种多样性的变化 Fig. 4 Changes in species diversity of different regions in 2016 and 2017 |
由样地纬度位置可知(表 1), 由北向南依次为较高纬度的马伦草原(38.75°N)、中间纬度的荷叶坪(38.71°N)、较低纬度的云顶山(37.88°N)分别为吕梁山系的偏北部山地、中部山地、偏南部山地。因此, 这种纬度梯度差异导致的吕梁山南北水热状况的差异, 使吕梁山系亚高山草甸群落物种多样性在空间上表现出由北向南逐渐升高的变化格局。
3 讨论 3.1 吕梁山亚高山草甸重要值的变化植物群落的物种多样性是研究群落组成的重要指标, 而重要值作为反映物种在群落中地位和作用的综合指标, 被广泛应用于群落的物种比较[19]。通过重要值的计算, 可以反映群落的结构组成、群落动态以及各物种在构成群落时所起的不同作用[20]。本研究中, 吕梁山亚高山草甸植物群落中物种多样性的分布比较均匀, 矮嵩草和珠芽蓼、线叶嵩草和珠芽蓼、车前草和披碱草分别为马伦草原、荷叶坪、云顶山亚高山草甸群落的优势种, 优势种和次优势种均以菊科、蔷薇科、莎草科、禾本科植物为主。草本植物不同生活型主要受到降水条件的影响, 还需考虑水热状况与光照条件的影响[21]。吕梁山亚高山草甸群落的优势种和次优势种生活型均以多年生草本为主, 徐满厚等[22]研究也表明吕梁山不同纬度处的物种多样性指数都是多年生草本高于一年生草本。就整个吕梁山脉而言, 吕梁山属于暖温带大陆性季风气候, 四季分明, 春季干旱多风, 气温回升快, 昼夜温差较大, 符合多年生草本的生境特征。因此, 吕梁山系的水热状况格局决定了不同山地亚高山草甸群落植物生活型的变化。
3.2 吕梁山亚高山草甸物种多样性的时间变化格局植物的物种多样性是测定群落和生态系统特征的重要指标, 可以直接或间接的反映植物群落和生态系统的结构类型、稳定程度及生境差异等[23-24]。杨崇曜等[25]研究结果表明, 气候因子中的年均温、年降水量、潜在蒸散量和最冷月均温对群落物种多样性均具有显著影响, 但各气候因子对Pielou均匀度指数的影响均不显著。白家烨等[14]以及雷淑慧等[26]的研究发现荷叶坪亚高山草甸物种多样性分布较均匀。本研究得到的吕梁山亚高山草甸连续3年内的Pielou均匀度指数无明显变化, 群落的生态系统和物种竞争性相对稳定。杜京旗等[27]的研究发现降水条件是影响云顶山亚高山草甸植被分布的重要因素。李晋鹏等[28]的研究发现, 气候条件、人类干扰、演替阶段、生境条件的差异都可能是影响群落物种多样性的因素。本研究中, 吕梁山系不同山地亚高山草甸的物种多样性时间变化为先增后减的单峰变化趋势, 且在2016年为最高值, 造成这种变化格局的主要因素是不同年份降水量的差异, 结果与上述的影响因素相一致。此外造成亚高山草甸物种多样性时间变化的因素很多, 还需要考虑放牧和旅游干扰、热量、光照等条件[6]。马伦草原、荷叶坪、云顶山都是吕梁山著名的生态旅游地和优良的天然牧场, 旅游开发与放牧也是影响物种多样性时间变化的重要因素。在研究物种多样性的时间变化格局时, 还要根据物种特定生长环境和具体环境因子来确定时间变格局的影响因素。因此, 在稳定的自然因素影响下, 限制人类活动干扰是保护吕梁山亚高山草甸生态系统的有效措施。
3.3 吕梁山亚高山草甸物种多样性的空间变化格局不同植物对环境因子的响应程度具有差异性, 导致不同植物物种多样性的空间分布格局会随着环境梯度的变化产生不同的分异格局[29]。孙菊等[30]研究表明, 随着纬度降低草本的物种多样性逐渐增大。巴图娜存[31]等的研究也发现, 草本植物的物种多样性呈现由北向南逐渐升高的趋势。本研究得到吕梁山亚高山草甸物种多样性变化, 最高值出在偏南部的云顶山样地, 最低值在偏北部的马伦草原样地。在海拔高度大致相同的情况下, 这是由于吕梁山偏南部的云顶山与偏北部的马伦草原、荷叶坪相比, 降水条件、热量状况明显具有优势, 其水热条件的配合与北部山地相比更有利于亚高山草甸的生长, 南部云顶山的物种丰富度、均匀度以及多样性相比北部的马伦草原和荷叶坪更具有优势。马斌等[32]的研究发现, 随纬度的增加, 草本植物物种丰富度呈现下降趋势, 地理因素对于物种多样性有显著的影响, 而且经度的影响大于海拔的影响。因此, 受到纬度梯度影响的水热分配状况决定了吕梁山亚高山草甸物种多样性由北向南逐渐增加的空间变化格局。
4 结论吕梁山亚高山草甸生态系统比较稳定, 不同山地植被主要以菊科、蔷薇科、莎草科、禾本科植物为主, 其物种多样性由于受到降水条件和纬度梯度的影响, 在2016年明显高于其他年份, 而且从偏北部的马伦草原到中部的荷叶坪再到偏南部的云顶山, 物种多样性在空间上呈现出由北向南逐渐升高的变化格局。因此, 降水条件和纬度梯度导致的环境差异对吕梁山亚高山草甸物种多样性的时空变化格局产生了重要影响。
本研究涉及研究范围及研究时限有限, 且仅调查了吕梁山部分亚高山草甸夏季草本植物, 对全面反映吕梁山亚高山草甸生物多样性的变化具有一定局限性, 今后可以此为基础, 融入其他环境因子, 延长调查时间, 扩大研究范围, 使之具有完整时空梯度, 深入研究亚高山草甸物种多样性与环境关系的形成机制, 进一步对亚高山草甸的生态保护和治理提供理论依据。
| [1] | 陈灵芝, 钱迎倩. 生物多样性科学前沿. 生态学报, 1997, 17(6): 565–572. |
| [2] | Tilman D, Reich P B, Knops J, Wedin D, Mielke T, Lehman C. Diversity and productivity in a long-term grassland experiment. Science, 2001, 294(5543): 843–845. DOI:10.1126/science.1060391 |
| [3] | Grabherr G, Gottfried M, Paull H. Climate effects on mountain plants. Nature, 1994, 369(6480): 448. |
| [4] | 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 1997, 17(1): 91–99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
| [5] | 贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征. 植物生态学报, 1998, 22(4): 303–311. DOI:10.3321/j.issn:1005-264X.1998.04.003 |
| [6] | 向春玲, 张金屯. 东灵山亚高山草甸物种多样性变化及其影响因子. 北京师范大学学报:自然科学版, 2009, 45(3): 275–278. |
| [7] | 王世昌. 云顶山亚高山草甸物种多样性研究. 山西林业科技, 2011, 40(3): 17-19, 23-23. DOI:10.3969/j.issn.1007-726X.2011.03.007 |
| [8] | 江源, 黄晓霞, 刘全儒, 和克俭, 杨艳刚. 五台山高山、亚高山草甸植物多样性格局分析. 北京师范大学学报:自然科学版, 2009, 45(1): 91–95. |
| [9] | 沙威, 董世魁, 刘世梁, 刘全儒, 石建斌, 李晓文, 苏旭坤, 吴娱. 阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素. 生态学杂志, 2016, 35(2): 330–337. |
| [10] | 白晓航, 张金屯, 曹科, 王云泉, Sadia S, 曹格. 小五台山亚高山草甸的群落特征及物种多样性. 草业科学, 2016, 33(12): 2533–2543. DOI:10.11829/j.issn.1001-0629.2016-0281 |
| [11] | 白晓航, 张金屯. 小五台山森林群落优势种的生态位分析. 应用生态学报, 2017, 28(12): 3815–3826. |
| [12] | 白晓航, 张金屯, 曹科, 王云泉, Sadia S, 曹格. 河北小五台山国家级自然保护区森林群落与环境的关系. 生态学报, 2017, 37(11): 3683–3696. |
| [13] | 周红章, 于晓东, 罗天宏, 何君舰. 物种多样性变化格局与时空尺度. 生物多样性, 2000, 8(3): 325–336. DOI:10.3321/j.issn:1005-0094.2000.03.015 |
| [14] | 白家烨, 刘卫华, 赵冰清, 张青, 郭东罡. 芦芽山荷叶坪亚高山草甸生物多样性. 应用生态学报, 2018, 29(2): 389–396. |
| [15] | 李素清, 张金屯, 上官铁梁. 芦芽山亚高山草甸的数量分类与排序研究. 西北植物学报, 2005, 25(10): 2062–2067. DOI:10.3321/j.issn:1000-4025.2005.10.022 |
| [16] | 李静怡, 王艳慧. 吕梁地区生态环境质量与经济贫困的空间耦合特征. 应用生态学报, 2014, 25(6): 1715–1724. |
| [17] | 徐满厚, 刘敏, 薛娴, 翟大彤, 彭飞, 尤全刚, 刘忠权. 增温、刈割对高寒草甸植被物种多样性和地下生物量的影响. 生态学杂志, 2015, 34(9): 2432–2439. |
| [18] | 徐满厚, 马丽, 白皓宇, 冀钦, 李晓丽. 山西吕梁山植被群落多样性的垂直空间分异. 江苏农业科学, 2017, 45(12): 256–260. |
| [19] | 牛钰杰, 杨思维, 王贵珍, 刘丽, 花立民. 放牧干扰下高寒草甸物种多样性指数评价与选择. 应用生态学报, 2017, 28(6): 1824–1832. |
| [20] | 卢爱英, 张先平, 王世裕, 王孟本. 干扰对云顶山亚高山草甸群落物种多样性的影响. 植物研究, 2011, 31(1): 73–78. |
| [21] | 何明珠. 阿拉善高原荒漠植被组成分布特征及其环境解释Ⅴ.一年生植物层片物种多样性及其分布特征. 中国沙漠, 2010, 30(3): 528–533. |
| [22] | 徐满厚, 白皓宇, 冀钦, 李明明, 马晨阳, 李晓丽, 杨朝. 山西吕梁山植被群落多样性的纬度变化. 价值工程, 2016, 35(20): 198–202. |
| [23] | 张丽霞, 张峰, 上官铁梁. 芦芽山植物群落的多样性研究. 生物多样性, 2000, 8(4): 361–369. DOI:10.3321/j.issn:1005-0094.2000.04.002 |
| [24] | 郑元润. 大青沟森林植物群落物种多样性研究. 生物多样性, 1998, 6(3): 191–196. DOI:10.3321/j.issn:1005-0094.1998.03.006 |
| [25] | 杨崇曜, 李恩贵, 陈慧颖, 张景慧, 黄永梅. 内蒙古西部自然植被的物种多样性及其影响因素. 生物多样性, 2017, 25(12): 1303–1312. DOI:10.17520/biods.2017140 |
| [26] | 雷淑慧, 卢爱英, 张先平, 王孟本. 庞泉沟自然保护区森林群落物种多样性. 生态学杂志, 2009, 28(12): 2431–2436. |
| [27] | 杜京旗, 张巧仙, 田晓东, 王悦, 许馨云, 宋丽, 袁自强. 云顶山亚高山草甸植被分布、物种多样性与土壤化学因子的相关性. 植物研究, 2016, 36(3): 444–451. |
| [28] | 李晋鹏, 上官铁梁, 郭东罡, 孟东平. 山西吕梁山南段植物群落物种多样性与环境的关系. 山地学报, 2008, 26(5): 612–619. DOI:10.3969/j.issn.1008-2786.2008.05.016 |
| [29] | Rey Benayas J M. Patterns of diversity in the strata of boreal montane forest in British Columbia. Journal of Vegetation Science, 1995, 6(1): 95–98. DOI:10.2307/3236260 |
| [30] | 孙菊, 李秀珍, 胡远满, 王宪伟, 吕久俊, 李宗梅, 陈宏伟. 大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度. 应用生态学报, 2009, 20(9): 2049–2056. |
| [31] | 巴图娜存, 胡云锋, 毕力格吉夫, 刘纪远, 甄霖. 蒙古高原乌兰巴托-锡林浩特草地样带植物物种的空间分布. 自然资源学报, 2015, 30(1): 24–36. |
| [32] | 马斌, 周志宇, 张莉丽, 高文星, 陈善科, 张宝林. 阿拉善左旗植物物种多样性空间分布特征. 生态学报, 2008, 28(12): 6099–6106. DOI:10.3321/j.issn:1000-0933.2008.12.038 |