 2018, Vol. 38
2018, Vol. 38文章信息
- 罗渡, 汪学杰, 徐猛, 顾党恩, 牟希东, 韦慧, 杨叶欣, Cesar G. Demayo, 胡隐昌.
- LUO Du, WANG Xuejie, XU Meng, GU Dang'en, MU Xidong, WEI Hui, YANG Yexin, Cesar G. Demayo, HU Yinchang.
- 贝类壳-体质量比和静水沉降特性的相关性
- Correlation between shell-body mass ratio and hydrostatic settling characteristics of mollusc species
- 生态学报. 2018, 38(18): 6778-6785
- Acta Ecologica Sinica. 2018, 38(18): 6778-6785
- http://dx.doi.org/10.5846/stxb201803250589
-
文章历史
- 收稿日期: 2018-03-25
- 修订日期: 2018-07-05





2. Department of Biological Sciences, College of Science and Mathematics, Mindanao State University-Iligan Institute of Technology, Iligan City, Philippines
2. Department of Biological Sciences, College of Science and Mathematics, Mindanao State University-Iligan Institute of Technology, Iligan City, Philippines
贝类物种丰富、利害兼有, 其数量、分布与人类生产、生活密切相关[1]。由于贝类的主动运动主要依靠肉足驱动, 其种群扩散在行为上更多地表现为被动扩散。物种扩散的媒介或障碍往往能对种群、进化或生态系统产生广泛的效应[2]。不仅是水生贝类, 即使陆生螺类, 水依然是其存活的重要条件, 水流是其远距离迁移的主要媒介之一[3]。研究表明水流介导了钉螺的迁移, 并最终导致血吸虫病的蔓延[4]。福寿螺、非洲大蜗牛等恶性入侵生物的渗透式扩散和爆发成灾也往往随洪水而发生。因此水介质在贝类的物种传播上起着重要的作用。贝类在水下的沉降规律是研究其运动规律的重要内容之一[5]。
在贝类学的研究初期, 贝壳特征几乎成了分类特征的唯一来源[6]。长期以来, 贝壳形态特征都是分类的主要依据[7], 但是贝壳形态也会随着环境变化而变化, 贝壳的细微结构在种的水平上也具有差异[8]。贝类具有较大的表型可塑性, 即使是不同地理种群, 生活的环境差异以及长期的适应性进化在形态上都可能产生明显分化[9]。尽管贝壳几何学引起学者的注意已经有200多年了, 然而对构造规则如何发挥作用以及形态进化是如何发生的还知之甚少。运动需要能量, 贝壳在发挥着保护和支持作用的同时也是一种运动负担。有壳动物在爬行、游泳或掘穴时, 肌肉所施加的力量都必须足以克服贝壳的重量。碳酸钙的密度比水的大三倍左右, 减少贝壳重量将增加其个体的机动性能[10]。
贝类在贝壳厚度、生活史策略等特征上都存在快速进化的能力[11]。运动行为在软体动物的生态和进化上具有重要作用, 长期的适应性选择往往可以使其运动扩散能力出现特化[12]。贝壳是软体动物的支撑和保护器官, 但是贝壳质量的增加不利于其捕食和迁徙。此外, 在进化过程中, 为了适应不同的生境, 贝壳会通过异速生长产生差异, 但是此种差异在“壳-体质量比”上的体现以及此种变化对于运动扩散功能的影响未见相关报道。生态水力学是研究生命体在水中的扩散转移规律及流场控制技术的新兴交叉型学科[4]。因此本研究选取9种具代表性的常见贝类作为研究对象, 运用形态学与生态水力学相结合的交叉学科方法, 通过分析壳-体质量比和静水沉降特性的种间差异, 及壳-体质量比与静水沉降速度、静水沉降加速度之间的相关关系, 以期探讨贝壳形态的适应性进化与水环境介导的运动扩散能力进化的关系, 为阐明贝类种群扩散的生态策略提供理论基础。
1 材料与方法 1.1 实验材料实验于珠江水产研究所外来水生生物入侵风险评估中心进行, 所用贝类为南方地区常见贝类, 样品主要采集于广东省境内, 每个物种的样本量≥30。所有实验贝类样品皆为成体, 9种贝类样品分属于9个属, 其物种分类信息如表 1所示。
| 编号 Number |
物种 Species |
纲 Class |
目 Order |
科 Family |
属 Genus |
| 1 | 福寿螺 Pomacea canaliculata |
腹足纲 Gastropoda |
中腹足目 Mesogastropoda |
瓶螺科 Ampullariidae |
瓶螺属 Pomacea |
| 2 | 中华圆田螺 Cipangopaludina cathayensis |
腹足纲 Gastropoda |
中腹足目 Mesogastropoda |
田螺科 Viviparidae |
圆田螺属 Cipangopaludina |
| 3 | 铜锈环棱螺 Bellamya aeruginosa |
腹足纲 Gastropoda |
中腹足目 Mesogastropoda |
田螺科 Viviparidae |
环棱螺属 Bellamya |
| 4 | 平角卷螺 Planorbarius corneus var. |
腹足纲 Gastropoda |
基眼目 Basommatophora |
扁卷螺科 Planorbidae |
扁卷螺属 Planorbarius |
| 5 | 椭圆萝卜螺 Radix swinhoei |
腹足纲 Gastropoda |
基眼目 Basommatophora |
椎实螺科 Lymnaeidae |
萝卜螺属 Radix |
| 6 | 非洲大蜗牛 Achatina fulica |
腹足纲 Gastropoda |
柄眼目 Stylommatophora |
非洲大蜗牛科 Achatinidae |
非洲大蜗牛属 Achatina |
| 7 | 同型巴蜗牛 Bradybaena similaris |
腹足纲 Gastropoda |
柄眼目 Stylommatophora |
巴蜗牛科 Bradybaenidae |
巴蜗牛属 Bradybaena |
| 8 | 背角无齿蚌 Sinanodonta woodiana |
双壳纲 Bivalvia |
蚌目 Unionoida |
蚌科 Unionidae |
无齿蚌属 Sinanodonta |
| 9 | 河蚬 Corbicula fluminea |
双壳纲 Bivalvia |
帘蛤目 Veneroida |
蚬科 Corbiculidae |
蚬属 Corbicula |
(1) 形态测量:贝类个体的形态参数用游标卡尺(精确到0.001 cm)进行测量, 所测指标包括:壳长, 壳顶沿中轴到壳底部的最大长度;壳宽, 垂直于壳长中轴的最大宽度[13-14];对于双壳贝类, 壳长为壳体前后方向最大距离;壳宽为壳体背部与腹部方向最大距离[15]。用吸水纸吸干表面水分后, 用电子天平称量体质量(精确到0.01 g)和壳质量(精确到0.01 g), 所有质量均为湿重[16]。用变异系数(Coefficient of Variation, C.V.)表征数据的离散程度大小, 其值为标准差与平均值之比。
(2) 壳-体质量比计算:按以下公式计算。
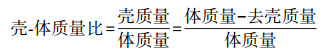
|
(3) 静水沉降特性的测定:采用静水沉降法, 设置沉降缸模拟自然条件下的沉降特征。沉降缸为圆柱体形, 高2.4 m、直径1.2 m, 以清澈的池塘养殖用水灌装。于水面中心点以下15 cm设置沉降起始点, 于起始点以下140 cm的垂直平面设置沉降计时终点, 终点距离缸底垂直距离大于25 cm, 缸底设置“缓冲网”以使样品免受机械刺激和损伤。所有受试个体皆以体长轴平行于水面的姿态进入自然沉降。以秒表记录起点至终点的运动时间, 测量重复3次(图 1)。依据公式v=S/t计算平均沉降速度(其中, v为沉降速度, S为沉降距离、t为沉降时间);假设样品为匀加速运动, 依据公式a=2S/t2计算加速度(其中a为沉降加速度, S为沉降距离、t为沉降时间)。

|
| 图 1 静水沉降示意图 Fig. 1 Settlement process of the molluscs in this study |
采用Microsoft office 2010软件、OriginPro 8.6软件和IBM SPSS Statistics 20软件等进行数据统计、差异性比较、回归分析和作图。依据前述公式计算壳-体质量比和沉降速度、沉降加速度。结果表示为:平均值±标准差。以“壳长”、“壳宽”、“体质量”、“壳-体质量比”等作为因变量, 采用单因素方差分析法(ANOVA)分析种间差异显著性(P<0.01)。
2 结果与分析 2.1 供试贝类的主要形态测量分别测量9种贝类的壳长、壳宽和体质量三个主要的形态指标, 其测量结果如表 2所示。由表可知, 背角无齿蚌(Sinanodonta woodiana Lea, 1834)的壳长、壳宽、体质量最大, 平角卷螺(Planorbarius corneus var.)的壳长最小, 椭圆萝卜螺(Radix swinhoei H.Adams)的壳宽和体质量最小。9种贝类的壳长、壳宽、体质量的总体变异系数(C.V.)分别为0.67, 0.54, 1.11, 因此体质量的变异>壳长的变异>壳宽的变异。
| 编号 Number |
物种 Species |
壳长 Shell length/cm |
壳宽 Shell width/cm |
体质量 Body mass/g |
| 1 | 平角卷螺Planorbarius corneus var. | 1.03±0.09A | 1.81±0.11BC | 1.15±0.19A |
| 2 | 同型巴蜗牛Bradybaena similaris | 1.35±0.24AB | 1.46±0.21AB | 1.13±0.44A |
| 3 | 椭圆萝卜螺Radix swinhoei | 1.60±0.18B | 1.02±0.11A | 0.51±0.13A |
| 4 | 铜锈环棱螺Bellamya aeruginosa | 2.34±0.33C | 1.69±0.18B | 2.70±1.08A |
| 5 | 河蚬Corbicula fluminea | 2.35±0.35C | 2.16±0.26C | 4.19±1.55A |
| 6 | 中华圆田螺Cipangopaludina cathayensis | 4.54±0.50D | 3.69±0.36D | 20.67±6.07B |
| 7 | 福寿螺Pomacea canaliculata | 5.59±1.29E | 4.72±0.75E | 32.58±14.87C |
| 8 | 非洲大蜗牛Achatina fulica | 6.23±1.45E | 3.45±0.70D | 31.36±17.13C |
| 9 | 背角无齿蚌Sinanodonta woodiana | 7.75±0.46F | 5.13±0.49E | 43.83±11.73D |
| 同列数据无相同字母表示在0.01水平上差异极显著 | ||||
研究结果显示(图 2):9种贝类的平均壳-体质量比为(0.32±0.13), 所有受试贝类壳-体质量比的种内C.V.<0.36, 平均C.V.=0.20, 背角无齿蚌变异最大(C.V.=0.35), 河蚬变异最小(C.V.=0.11), 因此贝类壳-体质量比的种属特异性强。不同种贝类之间存在明显的种间变异, 其中非洲大蜗牛的壳-体质量比最小(0.17±0.04), 极显著地低于其他贝类(P<0.01), 河蚬的壳-体质量比最大(0.54±0.06), 极显著高于其他贝类(P<0.01), 福寿螺的壳-体质量比接近0.50(0.50±0.06), 铜锈环棱螺次之(0.38±0.06)。中华园田螺(0.22±0.03)、背角无齿蚌(0.23±0.08)、同型巴蜗牛(0.26±0.08)、平角卷螺(0.26±0.04)和椭圆萝卜螺(0.28±0.04)的壳体质量比为0.25左右, 种间差异性较小(P<0.01), 只有中华园田螺和椭圆萝卜螺之间存在极显著差异(P<0.01), 其余差异不显著。陆生贝类的总体壳-体质量比(0.21)显著小于水生贝类(0.35)(P<0.01), 但并非所有陆生贝类的壳-体质量比都较水生贝类小。

|
| 图 2 贝类壳-体质量比的种间差异 Fig. 2 Interspecific difference of the ratio of shell-mass to body-mass |
观察发现, 除中华园田螺、铜锈环棱螺和河蚬外, 其余贝类均具备在水面漂浮的能力。在静水沉降实验中, 壳形是影响沉降姿势的主要因素, 双壳贝类和螺类的沉降行为差异明显, 其中双壳类的沉降稳定性较差, 沉降终点距离理想降落点的距离较大, 而螺类多为螺旋形下沉或平稳形下沉, 壳形接近的物种沉降行为相似。不同贝类沉降速度的种间差异如图 3所示。由图可知, 贝类沉降速度的变异范围较大, 其中椭圆萝卜螺的沉降速度最低((4.13±0.96) cm/s), 只与平角卷螺((9.86±3.04) cm/s)差异不显著, 但极显著低于其他贝类(P<0.01);而河蚬的沉降速度最大((24.99±4.22) cm/s), 极显著高于其他贝类(P<0.01)。同为水生贝类, 河蚬的沉降速度是椭圆萝卜螺沉降速度的6倍;平角卷螺、同型巴蜗牛((12.73±3.93) cm/s)和背角无齿蚌((13.04±3.73) cm/s)3种贝类的种间沉降速度差异不显著, 但与其他贝类差异极显著(P<0.01);非洲大蜗牛((18.30±3.64) cm/s)、铜锈环棱螺((19.48±3.14) cm/s)、中华园田螺((21.44±3.92) cm/s)和福寿螺((21.77±5.23) cm/s)4种贝类的种间沉降速度差异不显著, 但是与其他贝类差异极显著(P<0.01)。

|
| 图 3 静水沉降速度的种间差异 Fig. 3 Interspecific difference of the settling velocity in still water |
沉降加速度的种间差异如图 4所示, 由图可知椭圆萝卜螺的沉降加速度最小(0.25±0.11 cm/s2), 平角卷螺、同型巴蜗牛和背角无齿蚌的沉降加速度较为接近, 处于较低水平。非洲大蜗牛、铜锈环棱螺、中华园田螺和福寿螺4种贝类的沉降加速度较为接近, 处于较高水平, 河蚬的沉降加速度最大(9.17±3.20 cm/s2), 极显著的高于其他贝类(P<0.01)。贝类的沉降速度和沉降加速度的种间差异不尽相同, 但是种间差异的趋势基本一致。

|
| 图 4 静水沉降加速度的种间差异 Fig. 4 Interspecific difference of the settling acceleration in still water |
9种受试贝类的沉降速度和沉降加速度与壳-体质量比的相关关系如图 5所示, 由图可知, 总体上, 沉降速度、沉降加速度与壳-体质量比的均值之间存在一定的相关性, 相关性系数R2分别为0.28和0.39, 但是没有明显的线性关系。而指数函数(0.15, 0.15)、幂函数(0.09, 0.09)和对数函数(0.19, 0.28)回归分析得到的相关性系数更低, 拟合度更小。

|
| 图 5 壳-体质量比与沉降速度、沉降加速度的相关关系 Fig. 5 The relationship between shell-mass: body-mass and the settling characteristics (velocity and acceleration) |
软体动物种类繁多且分布广泛, 现存种和化石种共有近15万种, 贝壳是最重要的保护器官, 其形成主要依靠外套膜表皮细胞的分泌物。瓣鳃纲(Lamellibranchia)贝类约2万种, 全部营水生生活, 大部分海产, 少部分淡水种。腹足纲(Gastropoda)贝类约10万种, 一般通称螺类, 包括前鳃亚纲(Prosobranchia)、后鳃亚纲(Opisthobranchia)及肺螺亚纲(Pulmonata)。广布于海洋、淡水和陆地。贝类的干重、湿重与壳长、壳宽、壳高等主要形态学参数存在明显的线性关系, 这些关系在河蚬[15]、环棱螺[16]、萝卜螺[16]、福寿螺[14]等大多数物种中都得到了验证。带壳湿重(体质量)是估算淡水螺类生物量和能量值的最有效指标[16]。因此, 壳-体质量比是形态对环境变化适应性的一种综合反应。
壳形的可塑性和异速生长在贝类中已被广泛研究, 环境因素可以诱导壳形的微结构产生变异, 受环境因子时空变异的选择作用, 螺类的生态表型可在壳的厚薄和大小之间产生进化权衡[17]。因此尽管本研究中由于不同贝类生境特征的差异性和多样性[18], 水文环境特征可能并非是贝类壳-体质量比种间差异形成的主要原因, 但是陆生贝类与水生贝类之间、双壳类与螺类之间受水力学作用的差异性是显而易见的, 只是这种具体选择作用的实现过程还有待进一步研究[19]。
有研究表明, 淡水贝类在壳厚、壳型、生长以及繁殖等特征上的表型可塑性是海水贝类的三倍以上, 因此淡水贝类的表型可塑性在种间以及种内的差异可能是极其显著的[20], 这与本研究发现淡水贝类的壳-体质量比呈现出极显著的种间差异是相一致的(P<0.01)。有研究表明生活在海岸带的贝类, 其形态和行为都会受到水流的影响并产生适应性变化[10]。一般而言, 贝壳的保护作用和贝类的运动速度的进化方向是不相容的[10]。但是, 某些贝壳较厚的螺类更能够在一定的时间内抵御水禽的消化作用, 从而实现远距离传播, 这可能是贝壳同时具有保护作用和促进扩散能力的另一种体现[21]。
环境对贝壳形态不具备决定作用, 而只有调控作用[22]。古代湖泊中的软体动物贝壳形态与其捕食者的进化有关, 对于捕食者驱动的贝类贝壳形态进化, 淡水贝类不如海洋贝类那么明显[10]。具备最坚固贝壳的淡水贝类只在古湖泊和古河流中有发现, 然而这种普遍的反捕食特征在现代贝类中已经不复存在了[23]。大量研究表明, 淡水螺类的壳形态指数与体质量显著相关, 但是相关关系会随着生境条件的变化而不同[24]。除了本文所研究的成体贝类的壳-体质量比特征以外, 贝壳的强度、硬度和弹性模量等力学性能指标也会随着贝壳的生长而增大[25]。因此, 壳-体质量比的种内时空变异还需要进一步的研究。
3.2 贝类水力学沉降特性的种间差异贝类水力学沉降特征研究属于生态水力学的范畴。生态水力学作为一门新兴交叉学科, 关注水力学与生态过程之间的关联, 研究水力条件的改变对水生态系统平衡以及对生物多样性的影响是其最重要的研究内容之一[26-27]。贝类水力学沉降特性种间差异的研究是对水文与生物体之间互作研究的一个有益补充[28-29]。极端气候和人类活动影响下, 贝类水力学沉降特征的明确有助于对入侵、进化等相关生态过程的理解[30]。
有研究表明河蚬形态在流域间存在显著的区域变化, 区域间壳体形态差异是水流速度的差异所致, 而水体化学组成和温度变化与此种形态变化之间相关性不大[31]。沉降速度与沉降加速度是体质量与形态的综合作用结果。许多底栖无脊椎动物都没有对抗强烈水流的游泳能力[32], 因此只能作为近似颗粒在水流中被动运动, 随水漂流和吸附能力对于扩散、传播和定殖尤其重要[33]。在底栖动物生活史中利于种群扩散的时期可能不尽相同[34], 种群的扩散传播往往是通过整个生活史策略实现的。因此本研究中沉降速度较大的物种并不一定就是随水流扩散能力弱的物种, 不过其他生活史阶段的扩散能力还有待进一步研究。
3.3 贝类壳-体质量比与水力学沉降特性的相关关系本研究所涉及的贝类为我国南方地区常见的贝类, 相关性分析表明壳-体质量比和沉降特性之间的相关性总体上不显著, 并非壳-体质量比越大沉降速度或沉降加速度就越大。壳-体质量比是形态参数之间的一种关系, 沉降特性是形态、行为功能与水环境之间互作的一种反应。有研究表明螺类运动特征的进化通常是不稳定的, 但是体型大小在进化上则是相对较为保守的, 而体型大小是运动能力最主要的限制因子之一[10]。福寿螺和非洲大蜗牛的体型与同类型螺类相比优势极其显著, 都具有较强的运动、扩散能力, 此二种螺类极强的摄食、破坏能力也与体型大小相关[35], 并且成体阶段都具有水中漂浮的能力, 这可能部分解释了其成为恶性入侵生物的原因。这与水文格局变化、长距离运输和极端气候改变了腹足类在人类世的地理分布的研究结果也是一致的[36]。
尽管体型对于贝类在生态上具有无可争议的重要性, 但这并不意味着在宏观进化上就一定会有直接的体现[37]。形态上的特化将降低贝类的运动、扩散能力, 但是扩散能力是一个综合性状, 并不是由某一特征单独决定的。壳-体质量比和沉降特性之间的相关关系不显著, 可能预示着贝壳特征在很大程度上是综合生境适应的结果。沉降速度和沉降加速度是成体贝类随水流扩散能力的一种反应, 但生态水力学对于种群扩散的综合作用可能还需考虑整个生活史阶段的作用以及贝类的运动、繁殖行为特征[38]。例如, 福寿螺与中华园田螺和铜锈环棱螺的沉降速度差异并不显著, 但是前者在非应激状态下还具有在水中漂浮的能力, 使其具备更强的扩散能力。作为广布种, 这些贝类都具有较强的适应能力[39], 除了福寿螺和非洲大蜗牛等世界性入侵物种以外, 河蚬、背角无齿蚌、同型巴蜗牛等在某些地区也被当做入侵种, 种群扩散能力的研究是其入侵生态学研究的重要内容[40]。不过, 贝壳表型与沉降行为特征各自的进化路径与生态功能以及二者之间的相关关系对于种群扩散的作用力还有待深入研究[41]。
4 结论贝类的壳-体质量比存在极显著的种间差异, 种属特异性强, 水生贝类的壳-体质量比显著大于陆生贝类, “壳-体质量比”参数能较好的反应贝类在生境适应性上对运动与保护功能的权衡。贝类的静水沉降速度和沉降加速度存在极显著的种间差异, 成体贝类的壳-体质量比与沉降特性之间不存在普适性的显著相关关系, 其相关性的进一步揭示还需考虑水文对不同生活史时期贝类种群扩散的作用。同时, 贝类入侵种的危害性与其扩散能力密切相关, 而扩散能力是物种长期适应性进化的重要结果, 是包含物种个体形态特征、生理生态行为以及所处生境等多因素的综合策略。
| [1] | 刘月英, 张文珍. 淡水螺类与人们的关系. 动物学杂志, 1977, 12(2): 46–47. |
| [2] | Caplat P, Edelaar P, Dudaniec R Y, Green A J, Okamura B, Cote J, Ekroos J, Jonsson P R, Löndahl J, Tesson S V M, Petit E J. Looking beyond the mountain:dispersal barriers in a changing world. Frontiers in Ecology and the Environment, 2016, 14(5): 261–268. DOI:10.1002/fee.1280 |
| [3] | Dörge N, Walther C, Beinlich B, Plachter H. The significance of passive transport for dispersal in terrestrial snails (Gastropoda, Pulmonata). Zeitschrift für Ökologie und Naturschutz, 1999, 8: 1–10. |
| [4] | 李大美. 生态水力学. 北京: 科学出版社, 2006. |
| [5] | 潘昭汉, 叶廉, 陈和春, 何昌浩, 常汉斌. 钉螺静水和动水沉降试验研究. 武汉水利电力大学(宜昌)学报, 1998, 20(1): 21–27. |
| [6] | 王忠明, 陈德牛, 吴小平. 腹足类贝壳特征的测量及分析方法. 动物学杂志, 2003, 38(4): 62–65. DOI:10.3969/j.issn.0250-3263.2003.04.014 |
| [7] | Lu H F, Du L N, Li Z Q, Chen X Y, Yang J X. Morphological analysis of the Chinese Cipangopaludina species (Gastropoda; Caenogastropoda:Viviparidae). Zoological Research, 2014, 35(6): 510–527. |
| [8] | 欧阳珊, 吴小平. 蚌科贝壳细微结构及其在分类上的意义//中国动物学会全国显微与亚显微形态科学分会第十次形态科学与显微注射技术学术研讨会论文摘要集. 海口: 中国动物学会全国显微与亚显微形态科学分会, 2000. |
| [9] | Hynd J S. A revision of the Australian pearl-shells, genus Pinctada (Lamelli-branchia). Australian Journal of Marine and Freshwater Research, 1955, 6(1): 98–138. DOI:10.1071/MF9550098 |
| [10] | Vermeij G J. A Natural History of Shells. Princeton: Princeton University Press, 1995. |
| [11] | Chapuis E, Lamy T, Pointier J P, Juillet N, Ségard A, Jarne P, David P. Bioinvasion triggers rapid evolution of life histories in freshwater snails. American Naturalist, 2017, 190(5): 694–706. DOI:10.1086/693854 |
| [12] | Dahirel M, Olivier E, Guiller A, Martin M C, Madec L, Ansart A. Movement propensity and ability correlate with ecological specialization in European land snails:comparative analysis of a dispersal syndrome. Journal of Animal Ecology, 2015, 84(1): 228–238. DOI:10.1111/jane.2014.84.issue-1 |
| [13] | 蔡英亚, 张英, 魏若飞. 贝类学概论. 上海: 上海科学技术出版社, 1995. |
| [14] | 李玺洋, 罗渡, 顾党恩, 徐猛, 牟希东, 章家恩, 胡隐昌. 不同壳色福寿螺形态性状与体质量的关系. 生物安全学报, 2012, 21(4): 287–290. DOI:10.3969/j.issn.2095-1787.2012.04.009 |
| [15] | 张健, 高子涵, 马行厚, 李德亮, 肖调义. 大通湖河蚬(Corbicula fluminea)干重的换算方法. 湖南农业大学学报:自然科学版, 2014, 40(6): 633–636. |
| [16] | 赵伟华, 王海军, 王洪铸, 刘学勤. 淡水螺类去壳干重和无灰干重的换算方法. 应用生态学报, 2009, 20(6): 1452–1458. |
| [17] | Charrier M, Marie A, Guillaume D, Bédouet L, Le Lannic J, Roiland C, Berland S, Pierre J S, Le Floch M, Frenot Y, Lebouvier M. Soil calcium availability influences shell ecophenotype formation in the sub-antarctic land snail, Notodiscus hookeri. PLoS One, 2013, 8(12): e84527. DOI:10.1371/journal.pone.0084527 |
| [18] | 罗渡, 牟希东, 宋红梅, 顾党恩, 杨叶欣, 汪学杰, 罗建仁, 胡隐昌, 章家恩. 不同生境福寿螺(Pomacea canaliculata)的营养成分及其利用价值. 生态学杂志, 2012, 31(8): 2004–2010. |
| [19] | Wood P J, Hannah D M, Sadler J P. Hydroecology and Ecohydrology:Past, Present and Future. Hoboken: Wiley, 2008. |
| [20] | Bourdeau P E, Butlin R K, Brönmark C, Edgell T C, Hoverman J T, Hollander J. What can aquatic gastropods tell us about phenotypic plasticity? A review and meta-analysis. Heredity, 2015, 115(4): 312–321. DOI:10.1038/hdy.2015.58 |
| [21] | Van Leeuwen C H A, Van Der Velde G, Van Lith B, Klaassen M. Experimental quantification of long distance dispersal potential of aquatic snails in the gut of migratory birds. PLoS One, 2012, 7(3): e32292. DOI:10.1371/journal.pone.0032292 |
| [22] | Sokołowski A, Pawlikowski K, Wołowicz M, Garcia P, Namieśnik J. Shell deformations in the Baltic clam Macoma balthica from southern Baltic Sea (the Gulf of Gdansk):hypotheses on environmental effects. AMBIO:A Journal of the Human Environment, 2008, 37(2): 93–100. DOI:10.1579/0044-7447(2008)37[93:SDITBC]2.0.CO;2 |
| [23] | Vermeij G J, Covich A P. Coevolution of freshwater gastropods and their predators. American Naturalist, 1978, 112(987): 833–843. DOI:10.1086/283326 |
| [24] | Mehler K, Acharya K, Sada D W. Spatial and temporal pattern in length-mass regressions of freshwater gastropods in Nevada spring ecosystems. Malacologia, 2015, 58(1/2): 167–177. |
| [25] | 梁艳, 赵杰, 吴承伟. 不同生长期贝壳的结构和力学性能. 复合材料学报, 2011, 28(3): 121–126. |
| [26] | 陈求稳, 欧阳志云. 生态水力学耦合模型及其应用. 水利学报, 2005, 36(11): 1273–1279. DOI:10.3321/j.issn:0559-9350.2005.11.001 |
| [27] | Blanckaert K, Garcia X F, Steiger J, Uijttewaal W. Ecohydraulics:linkages between hydraulics, morphodynamics and ecological processes in rivers. Ecohydrology, 2013, 6(4): 507–510. DOI:10.1002/eco.v6.4 |
| [28] | 陆咏晴, 严岩, 丁丁, 赵春黎, 宋扬, 赵景柱. 我国极端降水变化趋势及其对城市排水压力的影响. 生态学报, 2018, 38(5): 1661–1667. |
| [29] | 廖文婷, 邓红兵, 李若男, 郑华. 水利工程对坝下径流的影响——以葛洲坝、三峡水利枢纽为例. 生态学报, 2018, 38(5): 1750–1757. |
| [30] | 张亚, 高学平. 生态水力学浅议//第五届全国环境水力学学术会议. 重庆:中国水利学会, 2003: 182–186. |
| [31] | 周会, 刘丛强, 闫慧, 丁文慈, 王兵, 江伟, 赵彦龙. 中国主要流域河蚬[Corbicula fluminea (Müller, 1774)]形态及其对环境的适应性. 生态学杂志, 2011, 30(7): 1497–1503. |
| [32] | Byers J E, Pringle J M. Going against the flow:retention, range limits and invasions in advective environments. Marine Ecology Progress Series, 2006, 313: 27–41. DOI:10.3354/meps313027 |
| [33] | Patrick C J, Cooper M J, Uzarski D G. Dispersal mode and ability affect the spatial turnover of a wetland macroinvertebrate metacommunity. Wetlands, 2014, 34(6): 1133–1143. DOI:10.1007/s13157-014-0572-8 |
| [34] | Qian P Y. Larval settlement of polychaetes. Hydrobiologia, 1999, 402: 239–253. DOI:10.1023/A:1003704928668 |
| [35] | Carlsson N O L, Brönmark C. Size-dependent effects of an invasive herbivorous snail (Pomacea canaliculata) on macrophytes and periphyton in Asian wetlands. Freshwater Biology, 2006, 51(4): 695–704. DOI:10.1111/fwb.2006.51.issue-4 |
| [36] | Capinha C, Essl F, Seebens H, Moser D, Pereira H M. The dispersal of alien species redefines biogeography in the Anthropocene. Science, 2015, 348(6240): 1248–1251. DOI:10.1126/science.aaa8913 |
| [37] | Jablonski D. Body-size evolution in Cretaceous molluscs and the status of Cope's rule. Nature, 1997, 385(6613): 250–252. DOI:10.1038/385250a0 |
| [38] | 汪伟, 梁幼生, 戴建荣, 黄轶昕. 南水北调工程对日本血吸虫中间宿主湖北钉螺(Oncomelania hupensis)分布的影响. 生态学报, 2008, 28(9): 4235–4245. DOI:10.3321/j.issn:1000-0933.2008.09.021 |
| [39] | 罗明珠, 章家恩, 胡九龙, 赵本良. 福寿螺和田螺消化酶活性比较. 生态学报, 2015, 35(11): 3580–3587. |
| [40] | De Medeiros Carvalho C, De Almeida Bessa E C, D'ávila S. Life history strategy of Bradybaena similaris (Fèrussac, 1821) (Mollusca, Pulmonata, Bradybaenidae). Molluscan Research, 2006, 28(3): 171–174. |
| [41] | Zieritz A, Clucas G, Axtmann L, Aldridge D C. Shell ecophenotype in the blue mussel (Mytilus edulis) determines the spatial pattern in foraging behaviour of an oystercatcher (Haematopus ostralegus) population. Marine Biology, 2012, 159(4): 863–872. DOI:10.1007/s00227-011-1862-9 |