 2018, Vol. 38
2018, Vol. 38文章信息
- 胡芳, 曾馥平, 杜虎, 彭晚霞, 张芳, 谭卫宁, 宋同清.
- HU Fang, ZENG Fuping, DU Hu, PENG Wanxia, ZHANG Fang, TAN Weining, SONG Tongqing.
- 桂西北喀斯特常绿落叶阔叶混交林物种多样性分布格局的尺度效应
- Scale-dependent spatial patterns for species diversity in a karst evergreen and deciduous broad-leaved mixed forest of northwest Guangxi
- 生态学报. 2018, 38(17): 6074-6083
- Acta Ecologica Sinica. 2018, 38(17): 6074-6083
- http://dx.doi.org/10.5846/stxb201709231713
-
文章历史
- 收稿日期: 2017-09-23
- 网络出版日期: 2018-05-30





2. 中国科学院环江喀斯特生态系统观测研究站, 环江 547100;
3. 广西壮族自治区木论国家级自然保护区管理局, 环江 547200
2. Huanjiang Observation and Research Station of Karst Ecosystem, Chinese Academy of Sciences, Huanjiang 547100, China;
3. Administrative Bureau of Mulun National Nature Reserve, Guangxi Zhuang Autonomous Region, Huanjiang 547200, China
物种多样性一般是指物种丰富度、多度或者是两者在特定区域下的组合[1]。物种多样性是当前生态学研究中十分重要的内容和热点之一[2], 不同的取样尺度下物种多样性可能不同[3], 因为空间尺度对物种多样性通常具有非线性影响, 此外, 异质性随着空间尺度的增加而增大, 在空间尺度增加的过程中常会导致较小尺度的信息丢失[4]。不同的多样性指数自身性质不同, 如物种丰富度和多度在不同空间尺度反应不同。一个尺度适合这个指数, 但是不一定适合另外的指数。尽管生态学家现在敏锐地意识到, 将信息从一个尺度转移到另一个尺度是必不可少的, 并在过去的20多年已经取得了很大进步[5-6], 尺度效应仍是一个具有挑战性的问题, 因为其对生物多样性研究以及保护具有重大意义[7]。因此, 分析不同空间尺度内物种多样性将有助于自然生态系统的保护与管理[1]。
物种多样性在不同气候带的地理位置上表现出明显的变化, 例如, 西双版纳20 hm2的热带森林动态监测样地有468种[8], 而长白山阔叶红松林25 hm2森林大样地仅51种[9]。即使在相似的气候带下, 物种多样性也有差异, 如茂兰喀斯特原始森林的1 hm2的样地中共有木本植物63科140属199种[10], 而木论喀斯特原始森林的0.8 hm2样地仅有43科91属123种[11]。在相同的研究区域随不同的取样尺度的变化, 物种多样性也不同[12-16]。因此, 研究物种多样性时非常有必要考虑到尺度[7]。
在热带雨林[12-13]、南亚热带常绿阔叶林[14]、温带针阔混交林[3]以及温带次生林[15]中开展的研究表明物种多样性具有尺度依赖性[12-14]。喀斯特地区普遍具有景观异质性高、土壤总量少、土层薄、岩石裸露率高、土层贫瘠等特点, 生境和植被具有高度的异质性, 植被随海拔变化垂直现象明显[11, 16-17], 已有张忠华等[10]对喀斯特地区两个1 hm2的样地的多样性进行了研究, 在更大尺度上的研究显得尤为重要。本文以木论喀斯特常绿落叶阔叶混交林的25 hm2动态监测样地为研究对象, 探讨了不同空间尺度下各物种多样性指标的空间分布特征, 以期为喀斯特森林在空间尺度和空间位置上的多样性模式提供重要的见解, 同时为物种的多样性的管理和保护提供一个合理的科学基础。
1 材料与方法 1.1 研究区概况木论国家自然保护区(25°07′01″—25°12′22″ N, 107°54′01″—108°05′51″ E)位于广西环江毛南族自治县的西北部的喀斯特峰丛洼地集中连片区, 南北长10.75 km, 东西宽19.80 km, 总面积约89.69 km2, 属中亚热带季风气候, 年均日照为1451 h, 无霜期为310天, 年平均气温为19.3 ℃, 年降雨量丰沛, 海拔420 m处年均降雨量为1529.2 mm(海拔420 m), 平均蒸发量为1571.1 mm, 平均相对湿度为79%。研究区属于亚热带石灰岩区常绿落叶阔叶混交林生态系统, 是我国生物区系相汇交错区的中心, 生境异质性极高, 区系成分复杂, 物种种类丰富, 生态环境脆弱, 是目前世界上喀斯特地区保存最完好、面积最大的原生林。林区岩石裸露率高达80%, 甚至部分区域达到90%以上, 土层浅且分布不均, 区内主要以碳酸盐岩发育的深色或棕褐色石灰土为主, 典型景观单元为峰丛洼地。
1.2 样地设置及调查自2013年初经过多次实地勘察, 反复测量并调整样地边框, 按照CTFS(Center for Tropical Forest Science)全球森林生物多样性监测规范[18], 于2014年在木论自然保护区内建设完成25 hm2森林动态监测样地。样地为正方形, 东西、南北边长均为500 m(图 1)[19]。整个样地被划分成625个20 m×20 m样方, 样方4角用水泥桩作永久标记。每个20 m ×20 m样方又细分为16个5 m×5 m小样方。野外调查以20 m×20 m样方为单元, 记录了其中所有胸径≥1 cm的木本植物树种名称、胸径、坐标, 计划今后每隔5年复查一次。首次调查结果表明, 样地共有胸径(DBH)≥1 cm的独立木本植物个体108, 667株, 隶属61科147属227种。

|
| 图 1 木论样地等高线图 Fig. 1 Contour map of Mulun forest plot |
本研究将整个25 hm2样地划分为以下6个空间尺度:0.0025 hm2(5 m×5 m), 0.01 hm2(10m×10 m), 0.04 hm2(20 m×20 m), 0.25 hm2(50 m×50 m), 1 hm2(100 m×100 m)和6.25 hm2(250 m×250 m)。6个空间尺度的样方数见表 1。分别计算6个空间尺度中样方内的物种多样性指数, 包括:多度、丰富度、Shannon-Wiener指数、Simpson指数以及Pielou均匀度指数等。
| 取样尺度Sampling scales | 5 m×5 m | 10 m×10 m | 20 m×20 m | 50 m×50 m | 100 m×100 m | 250 m×250 m |
| 相关系数Correlation coefficients | 0.344 | 0.478 | 0.365 | 0.325 | 0.450 | 0.400 |
| 样方数Number of plots | 10000 | 2500 | 625 | 100 | 25 | 4 |
| P | 0.000 | 0.000 | 0.000 | 0.001 | 0.024 | 0.600 |
各物种多样性指数[20]、方差、变异系数计算公式如下:
1) 丰富度指数S(Species richness): S =样方内物种数
2) Shannon-Wiener指数计算公式为: 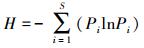
3) Simpson指数计算公式为: 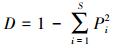
4) Pielou均匀度指数计算公式为: E=H/ln(S)
5) 方差: 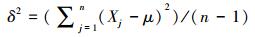
6) 变异系数: CV=(sd/μ)×100%
式中, Pi为单个样方内第i个物种的多度占所有多度之和的比例, S为单个样方内的物种数, n为样方总数, Xj为计算第j个样方物种多样性指标的数值, μ为平均值, sd为标准差。
根据计算结果, 绘制各多样性指数的空间分布图, 以对整个样地不同取样尺度下的物种多样性空间分布特点及空间变化进行分析。计算物种丰富度和多度在每一取样尺度下的Spearman秩相关系数, 探讨物种丰富度和多度的相关性随尺度变化的分异。同时利用多样性指标的方差和变异系数对不同取样面积的自然对数进行作图, 以探讨取样尺度的变化对物种多样性格局的影响。
数据统计分析和物种多样性空间分布格局制图均通过R 3.3.1软件完成, 物种丰富度与多度Spearman秩相关检验通过SPSS 21.0软件完成, 物种多样性指数空间变异制图通过ORINGIN 8.0软件完成。
2 结果 2.1 多度和物种丰富度的空间分布格局多度和物种丰富度随着取样空间尺度的增加而增加(图 2, 图 3)。在小尺度上(5 m×5 m至20 m×20 m)上, 多度和物种丰富度的空间分布格局表现出相同的趋势, 即样地中部洼地区域的多度和物种丰富度明显低于东部山脊以及西北角;在中等尺度(50 m×50 m), 这种分布差异不明显;在较大尺度(100 m×100 m至250 m×250 m)上, 多度和物种丰富度的空间分布格局明显不同, 在250 m×250 m的尺度上丰富度的最大值出现在东南角, 而多度的最大值出现在东北角。此外, 多度和物种丰富度的秩相关分析结果表明, 在5 m×5 m至20 m×20 m尺度上, 多度和物种丰富度呈现出极显著(P < 0.001)的正相关关系, 当尺度超过100 m×100 m时, 两者的相关性则不再显著(表 1)。

|
| 图 2 多度在6种取样尺度下的空间分布特征 Fig. 2 Spatial distribution of abundance index at six scales |

|
| 图 3 物种丰富度在6种取样尺度下的空间分布特征 Fig. 3 Spatial distribution of species richness index at six scales |
从图 4、图 5、图 6中可以看出, Shannon-Weiner指数、Simpson指数和Pielou均匀度指数等在不同的尺度上表现出相同的趋势。在最大尺度250 m×250 m(6.25 hm2)上, 3种多样性指数均是样地东南部的最高, 并且西半部多样性明显地于东半部;在中等尺度(100 m×100 m至20 m×20 m)上, 3个指数均在在西北角地区现了高值, 整体表现为山坡部分高于洼地、山顶区域, 符合“中间高度膨胀(mid-altitude bulge)”理论。在小尺度上(10 m×10 m至5 m×5 m), 每个样方下3个指数的数值都很低, 而且十分接近, 甚至很难从灰度图中区分出数值较高的样方。

|
| 图 4 Shannon-Weiner指数在6种取样尺度下的空间分布特征 Fig. 4 Spatial distribution of Shannon-Weiner index at six scales |

|
| 图 5 Simpson指数在6种取样尺度下的空间分布特征 Fig. 5 Spatial distribution of Simpson index at six scales |

|
| 图 6 Simpson指数在6种取样尺度下的空间分布特征 Fig. 6 Spatial distribution of Simpson index at six scales |
多度和物种丰富度的方差和变异系数随着取样尺度的增加有着类似的变化趋势(图 7)。多度和物种丰富度都随着尺度的增加表现出单峰型的变化特征, 且在100 m×100 m尺度上达到最大值。多度和物种丰富度的变异系数均随着尺度的增加呈现出线性减小的趋势, 而多度在小于100 m×100 m尺度上相对于物种丰富度较线性平缓减小。

|
| 图 7 多度和物种丰富度在6种尺度下的空间变异 Fig. 7 Spatial variation of species richness and abundance at six scales |
随着取样尺度的增加, Shannon-Weiner指数、Simpson指数和Pielou均匀度指数的方差都有着明显不同的变化趋势(图 8)。Shannon-Weiner指数、Simpson指数和Pielou均匀度指数的方差随着尺度的增加均表现出单峰型的变化特征, 且都是在100 m×100 m尺度上达到最大值。Shannon-Weiner指数、Simpson指数和Pielou均匀度指数的变异系数都随着尺度的增加呈现出线性减小的趋势, 且在5 m×5 m至20 m×20 m尺度上急剧下降, 之后则比较平缓。

|
| 图 8 Shannon-Weiner指数、Simpson指数和Pielou均匀度指数在6种尺度下的空间变异 Fig. 8 Spatial variation of Shannon-weiner, Simpson and Pielou evenness index at six scales |
物种丰富度指数、Shannon-Weiner指数、Simpson指数、Pielou均匀度指数等均没有叠加的特性, 特定的空间尺度上观察到的高的值可能在另一个空间尺度上表现为低, 这是因为当尺度增大时, 小尺度的物种丰富度不能简单的叠加。物种多样性指数在中尺度(100 m×100 m至20 m×20 m)上, 整体表现为山坡高于洼地区域和山顶, 这与前人研究结果相同, 木论喀斯特森林木本植物物种丰富度沿海拔梯度的曲线符合单峰格局, 峰值出现在坡腰位置[21]。这可能是由于样地处于亚热带区域, 海拔最高为651.4 m, 温度随海拔的升高下降不明显, 仍然适合植物的生长, 低海拔洼地地区, 土层较深厚、湿度条件较好, 但太阳直射光照时间不长, 仅限于一些喜荫和耐荫性强的物种, 导致物种较少;高海拔的山顶, 土层较薄, 山体陡峭, 日照时间长导致土壤中的水分蒸发旺盛, 不利于植物生长, 因而物种较少;而中等海拔的山坡地带, 土层厚度、温湿条件都相对而言适合植物生长, 因而物种相对丰富[21]。物种丰富度和多度指数在小尺度、中尺度以及的尺度上表现出明显的差异性, 这是因为小尺度上物种多样性主要是与样方内木本植物个体数量有关, 随着尺度的增加, 新生境的出现导致物种多样性随着空间尺度的变化表现出明显的差异性[22]。本研究结果表明物种丰富度和多度之间的相关性随空间取样尺度的增加逐渐变得不再显著, 类似的趋势同样出现在其他地区的热带山地雨林[13]、温带森林[3]。在本研究表明喀斯特常绿落叶阔叶混交林物种多样性指数均随着尺度的变化均表现出不连续性。这一结果意味着在生物多样性保护和管理时所谓的生物多样性“热点”地区的评估需要在多个尺度上进行评估[7]。另外, 本研究结果显示, 不同的物种多样性指数得出的结果不完全相同, 这与多样性指数的本身性质有关, 其中物种丰富度和多度主要受生境异质性(坡向和岩石裸露率)的影响[23], Shannon-Weiner多样性指数在群落类型、物种个体密度和生境差异等方面的反映结果较好, Simpson优势度指数是反映群落优势度的较好指标, 而Pielou均匀度指数受物种丰富度的影响, 即对样方面积的大小较敏感[24]。但就目前的各种物种多样性指数都不能完全客观地反映群落物种多样性的现状[25], Iudin等认为可以利用多维分形的方法来选择合适的物种多样性指数[26]。在木论喀斯特常绿落叶阔叶混交林木本植物多样性的测定中, 本研究所采用的物种多样性指数, 尽管表现形式有所不同, 但都可以明确反映物种数量的变化。
本文中用方差和变异系数来描述物种多样性指数的空间变异, 较好地描述了木论喀斯特常绿落叶阔叶混交林的物种多样性空间分布特征。本研究中, 物种多样性指数的方差随尺度的增加都表现出单峰分布的特征, 在100 m×100 m尺度上达到最大值, 与物种丰富度和多度格局分离尺度相一致, 这也印证了生态学家通过方差分析中产生的拐点来确定相应尺度下的生态过程的合理性。以往有研究认为这种单峰特征可能与样地内不同的稀有种组成有很大的关系[27], 本样地中有稀有种有100种, 占总树种数的39.37%[22], 而海南尖峰岭样地内的稀有物种数量为60种, 占总物种数的20.7%[27], 该区域多样性研究中没有出现类似的现象[12];类似的单峰曲线在其他研究中也出现过[14], 但是不同的森林类型其峰值的尺度不同。例如:海南尖峰岭热带山地雨林中物种丰富度的方差在20 m×20 m尺度上达到峰值[13], 而在长白山温带森林中物种丰富度的峰值出现在125 m×125 m尺度上[3]。在类似的森林类型中, 张忠华等研究表明物种丰富度的方差在25 m×25 m尺度上出现峰值[10], 这可能是由于不同的生境异质性导致的。同时, Shannon-Weiner指数、Simpson指数、Pielou均匀度指数的变异系数在5 m×5 m至20 m×20 m尺度上线性急剧减小, 之后则变化平缓, 表明小尺度比大尺度有更高的可变性。以上表明该区域的物种多样性具有区域依赖特征, 和鼎湖山亚热带常绿阔叶林[14]、长白山温带森林[9]以及海南尖峰岭热带森林[13]样地的研究结果一致。
张忠华等曾研究表明喀斯特常绿落叶阔叶混交林大多数物种是聚集分布的, 且聚集度随空间尺度的变化而变化[28];本样地前人研究结果表明也有类似的结果[29], 这在其他研究区域也是常见的[30-31]。一些研究者认为随机分布和规则分布取决于种内或种间对于资源的竞争, 而聚集分布则主要是生境异质性和种子传播方式导致的[32-33]。木论25 hm2喀斯特常绿落叶阔叶混交林样地的样地海拔在442.6—651.4 m之间, 最大高差达208.8 m。坡度范围0.12°—66.97°, 地形起伏较大, 生境的异质程度相当高, 导致物种多样性在不同的尺度上分布格局不同。对于喀斯特常绿落叶阔叶混交林来说, 植被类型、海拔高度、人为干扰、演替阶段、生境条件差异均对物种多样性指数产生一定的影响[34-35]。因此, 想要了解物种多样性随空间尺度的变化模式, 我们需要在类似的森林生态系统做更多的研究。
致谢: 感谢广西壮族自治区木论国家自然保护区管理工作人员和中南林业科技大学2012级生态学专业同学对样地调查给予的帮助。| [1] | Wang X G, Hao Z Q, Ye J, Zhang J, Li B H, Yao X L. Spatial variation of species diversity across scales in an old-growth temperate forest of China. Ecological Research, 2008, 23(4): 709–717. DOI:10.1007/s11284-007-0430-8 |
| [2] | Whittaker R J, Willis K J, Field R. Scale and species richness:towards a general, hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4): 453–470. DOI:10.1046/j.1365-2699.2001.00563.x |
| [3] | Crawley M J, Harral J E. Scale dependence in plant biodiversity. Science, 2001, 291(5505): 864–868. DOI:10.1126/science.291.5505.864 |
| [4] | He F L, Legendre P, Bellehumeur C, LaFrankie J V. Diversity pattern and spatial scale:a study of a tropical rain forest of Malaysia. Environmental and Ecological Statistics, 1994, 1(4): 265–286. DOI:10.1007/BF00469425 |
| [5] | Turner M G, O'Neill R V, Gardner R H, Milne B T. Effects of changing spatial scale on the analysis of landscape pattern. Landscape Ecology, 1989, 3(3/4): 153–162. |
| [6] | Gorresen P M, Willig M R, Strauss R E. Multivariate analysis of scale-dependent associations between bats and landscape structure. Ecological Applications, 2005, 15(6): 2126–2136. DOI:10.1890/04-0532 |
| [7] | He F L, LaFrankie J V, Song B. Scale dependence of tree abundance and richness in a tropical rain forest, Malaysia. Landscape Ecology, 2002, 17(6): 559–568. DOI:10.1023/A:1021514104193 |
| [8] | 兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才. 西双版纳热带森林动态监测样地——树种组成与空间分布格局. 植物生态学报, 2008, 32(2): 287–298. DOI:10.3773/j.issn.1005-264x.2008.02.006 |
| [9] | 郭忠玲, 马元丹, 郑金萍, 刘万德, 金哲峰. 长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究. 应用生态学报, 2004, 15(11): 2013–2018. DOI:10.3321/j.issn:1001-9332.2004.11.005 |
| [10] | 张忠华, 胡刚, 祝介东, 倪健. 喀斯特常绿落叶阔叶混交林物种多度与丰富度空间分布的尺度效应. 生态学报, 2012, 32(18): 5663–5672. |
| [11] | 宋同清, 彭晚霞, 曾馥平, 王克林, 曹洪麟, 李先琨, 覃文更, 谭卫宁, 刘璐. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征. 生物多样性, 2010, 18(4): 355–364. |
| [12] | Kallimanis A S, Halley J M, Vokou D, Sgardelis S P. The scale of analysis determines the spatial pattern of woody species diversity in the Mediterranean environment. Plant Ecology, 2008, 196(1): 143–151. DOI:10.1007/s11258-007-9341-6 |
| [13] | 李艳朋, 许涵, 李意德, 骆土寿, 陈德祥, 周璋, 林明献, 杨怀. 海南尖峰岭热带山地雨林物种多样性空间分布格局的尺度效应. 植物生态学报, 2016, 40(9): 861–870. |
| [14] | 王志高, 叶万辉, 曹洪麟, 练琚愉. 鼎湖山季风常绿阔叶林物种多样性指数空间分布特征. 生物多样性, 2008, 16(5): 454–461. DOI:10.3321/j.issn:1005-0094.2008.05.005 |
| [15] | 邓莉萍, 白雪娇, 秦胜金, 魏亚伟, 周永斌, 李露露, 牛沙沙, 韩美娜. 辽东山区次生林物种多样性的空间分布及尺度效应. 应用生态学报, 2016, 27(7): 2197–2204. |
| [16] | 彭晚霞, 王克林, 宋同清, 曾馥平, 王久荣. 喀斯特脆弱生态系统复合退化控制与重建模式. 生态学报, 2008, 28(2): 811–820. DOI:10.3321/j.issn:1000-0933.2008.02.044 |
| [17] | 蒋忠诚, 李先琨, 曾馥平. 岩溶峰丛洼地生态重建. 北京: 地质出版社, 2007. |
| [18] | Condit R. Tropical Forest Census Plots:Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Berlin, Heidelberg: Springer, 1998. |
| [19] | 兰斯安, 宋敏, 曾馥平, 杜虎, 彭晚霞, 覃文更, 何铁光. 喀斯特常绿落叶阔叶混交林木本植物组成特征. 广西植物, 2016, 36(10): 1156–1164. DOI:10.11931/guihaia.gxzw201606018 |
| [20] | Hill M O. Diversity and evenness:a unifying notation and its consequences. Ecology, 1973, 54(2): 427–432. DOI:10.2307/1934352 |
| [21] | 兰斯安, 宋敏, 曾馥平, 宋同清, 彭晚霞, 杜虎, 韩畅, 陈莉, 覃文更. 木论喀斯特森林木本植物多样性垂直格局. 生态学报, 2016, 36(22): 7374–7383. |
| [22] | Turner W R, Tjørve E, Hillebrand H. Scale-dependence in species-area relationships. Ecography, 2005, 28(6): 721–730. DOI:10.1111/eco.2005.28.issue-6 |
| [23] | 彭晚霞, 宋同清, 曾馥平, 王克林, 傅伟, 刘璐, 杜虎, 鹿士杨, 殷庆仓. 喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系. 生态学报, 2010, 30(13): 3472–3481. |
| [24] | 马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究——Ⅱ丰富度、均匀度和物种多样性. 生态学报, 1995, 15(3): 268–277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
| [25] | Tóthmérész B. Comparison of different methods for diversity ordering. Journal of Vegetation Science, 1995, 6(2): 283–290. DOI:10.2307/3236223 |
| [26] | Iudin D I, Gelashvili D B, Rozenberg G S. Multifractal analysis of the species structure of biotic communities. Doklady Biological Sciences, 2003, 389(1/6): 143–146. DOI:10.1023/A:1023427010517 |
| [27] | 许涵, 李意德, 林明献, 吴建辉, 骆土寿, 周璋, 陈德祥, 杨怀, 李广建, 刘世荣. 海南尖峰岭热带山地雨林60 ha动态监测样地群落结构特征. 生物多样性, 2015, 23(2): 192–201. |
| [28] | Zhang Z H, Hu G, Zhu J D, Ni J. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China. Journal of Plant Ecology, 2013, 6(2): 131–140. DOI:10.1093/jpe/rts027 |
| [29] | Du H, Hu F, Zeng F P, Wang K L, Peng W X, Zhang H, Zeng Z X, Zhang F, Song T Q. Spatial distribution of tree species in evergreen-deciduous broadleaf karst forests in southwest China. Scientific Reports, 2017, 7(1): 15664. DOI:10.1038/s41598-017-15789-5 |
| [30] | Hardy O J, Sonke B. Spatial pattern analysis of tree species distribution in a tropical rain forest of Cameroon:assessing the role of limited dispersal and niche differentiation. Forest Ecology and Management, 2004, 197(1/3): 191–202. |
| [31] | 卢志军, 刘福玲, 吴浩, 江明喜. 八大公山常绿落叶阔叶混交林枯立木物种组成、大小级与分布格局. 生物多样性, 2015, 23(2): 167–173. |
| [32] | Boyden S, Binkley D, Shepperd W. Spatial and temporal patterns in structure, regeneration, and mortality of an old-growth ponderosa pine forest in the Colorado Front Range. Forest Ecology and Management, 2005, 219(1): 43–55. DOI:10.1016/j.foreco.2005.08.041 |
| [33] | Druckenbrod D L, Shugart H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont. Journal of Vegetation Science, 2005, 16(1): 37–48. DOI:10.1111/j.1654-1103.2005.tb02336.x |
| [34] | 黄忠良, 孔国辉, 何道泉. 鼎湖山植物群落多样性的研究. 生态学报, 2000, 20(2): 193–198. |
| [35] | 彭闪江, 黄忠良, 徐国良, 欧阳学军, 张池. 生境异质性对鼎湖山植物群落多样性的影响. 广西植物, 2003, 23(5): 391–398. DOI:10.3969/j.issn.1000-3142.2003.05.002 |