 2018, Vol. 38
2018, Vol. 38文章信息
- 庞碧剑, 李天深, 蓝文陆, 黎明民, 骆鑫, 陈莹.
- PANG Bijian, LI Tianshen, LAN Wenlu, LI Mingmin, LUO Xin, CHEN Ying.
- 钦州湾秋季和春季浮游动物分布特征及影响因素
- Distribution patterns and environmental factors of zooplankton in the Qinzhou Bay in spring and autumn
- 生态学报. 2018, 38(17): 6204-6216
- Acta Ecologica Sinica. 2018, 38(17): 6204-6216
- http://dx.doi.org/10.5846/stxb201708121451
-
文章历史
- 收稿日期: 2017-08-12
- 网络出版日期: 2018-05-30

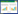




钦州湾位于广西北部湾的中部, 由内湾(茅尾海)和外湾(钦州港)组成, 是一个口小腹大呈葫芦形的半封闭海湾。近年来钦州湾沿岸的工农业、养殖业、港口、城市等迅速扩展, 该海域的生态环境承受着越来越大的压力[1]。浮游动物是食物链(网)中的关键环节, 部分种类同时也是生态环境变化的指示物种, 因此探究其变化规律及与环境因子的相互关系, 对保护海洋生态环境具有重要意义。但目前钦州湾这方面的研究很少, 仅见蓝文陆等[2]报道了丰水期(夏季)和枯水期(春季)的浮游动物群落特征, 不足以全面掌握钦州湾浮游动物的时空变化特征及其影响因素。此外2010—2015年是钦州湾快速发展的时段, 日益频繁的开发活动是否导致了浮游动物群落结构发生较大变化尚不明确。为此, 本文拟通过研究2014年秋季和2015年春季钦州湾浮游动物的群落结构、分布及其与主要环境因子的相关关系, 以期掌握该海湾浮游动物的时空变化规律和关键环境影响因子, 同时与2011—2012年的监测结果进行比较, 阐明钦州湾浮游动物年际变化以及现有的开发利用活动对浮游动物群落的影响, 为推动钦州湾生态环境保护和资源的可持续续利用提供科学依据。
1 材料与方法 1.1 样品采集与分析本研究在钦州湾内共布设了13个站位, 其中内湾5个, 外湾8个(图 1), 于2014年10月和2015年3月进行了2个航次的调查。其中浮游动物样品使用浅水Ⅰ型浮游生物网垂直拖网采集, 用终浓度5%(v/v)甲醛保存, 在显微镜和体视镜下对样品进行剔除杂质、分类和计数, 并称量浮游动物的生物量(湿重)。浮游动物样品的采集、保存、分析等均按《海洋监测规范》(GB17378.7—2007)[3]的要求进行。水温、盐度采用多参数水质仪(Multi 350)现场测定, 其他水质监测项目样品用8 L采水器采集后带回实验室分析。其中叶绿素a采用分光光度法测定, 硝酸盐采用镉柱还原法测定、亚硝酸盐采用萘乙二胺分光光度法测定、铵盐采用次溴酸盐氧化分光光度法测定、无机磷采用磷钼蓝分光光度法测定、硅酸盐采用硅钼黄法测定。样品的采集、保存、分析等按《海洋监测规范》(GB17378.7—2007)的要求进行。

|
| 图 1 钦州湾采样站位 Fig. 1 Sampling stations in QinzhouBay “Q”表示钦州湾 |
浮游动物群落的Shannon-Wiener多样性指数(H′)[3]和各站位物种多维尺度分析(MDS)使用Primer 6.0进行计算分析。环境因子的主成分分析(principal component analysis, PCA)以及浮游动物丰度与主要环境因子的相关性分析由SPSS 16.0完成。站位图、主要环境因子分布图以及浮游动物群落参数分布图采用Surfer 11.0克里金(Kriging)插值法绘制。
浮游动物优势度(Y)[4]的计算公式为:

|
式中, N为样品的总丰度;ni为样品中第i种的丰度;fi为该种在各样品中的出现频率。若Y≥0.02, 则认定该物种为优势种[5]。
2 结果 2.1 钦州湾秋季和春季主要环境因子的变化特征钦州湾秋季和春季表层水温变化范围分别为27.5—29.7℃、17.0—21.3℃(图 2);盐度变化范围分别为8.8—27.7和18.7—32.0, 均从内湾向外湾递增。秋季和春季叶绿素a浓度范围分别为0.7—4.9 μg/L、0.5—3.3 μg/L, 秋季内湾的叶绿素a浓度与外湾相近, 春季则外湾明显高于内湾(图 2)。秋季和春季的无机氮、硅酸盐和磷酸盐分布均是由内湾向外湾递减, 秋季钦州湾海域营养盐浓度均值略高于春季。

|
| 图 2 钦州湾秋季和春季主要环境因子的分布 Fig. 2 Distribution of environmental parameters in autumn and spring in the Qinzhou Bay |
钦州湾2次调查共检出浮游动物111种(类), 包括14类浮游幼虫, 浮游动物种类名录见表 1。钦州湾浮游动物以桡足类种类数最多, 其次为毛颚类, 浮游幼虫居第三。其中秋季出现13大类87种, 春季则出现11大类48种(表 2), 秋季种类多样性明显高于春季, 优势类群呈现出由秋季的暖水性种类向春季的暖温性种类更替的特征。钦州湾内湾和外湾的种类数量差异较大。秋季内湾和外湾种类数分别为12种和84种, 春季内湾和外湾种类数分别为10种和41种。内湾浮游动物种类数较少, 秋季内湾各站位平均种类数为4种, 春季内湾各站位平均出现种类数为3种。内湾以中小型桡足类以及浮游幼虫为主要类群, 代表种类有太平洋纺锤水蚤、中华异水蚤、长尾类幼虫以及短尾类幼虫。外湾浮游动物种类较丰富, 秋季和春季各站位平均出现的种类分别为10种(类)和20种(类), 此外外湾浮游动物类群中个体较大的种类所占比例远高于内湾。
| 种名 Species |
秋季 Autumn |
春季 Spring |
| 夜光虫Noctiluca scientillans | + | |
| 拟帽水母Paratiara digitalis Kramp et Damas | + | |
| 蟹形和平水母Eirene kambara | + | |
| 细颈和平水母Eirene menoni | + | |
| 刺胞水母Cytaeis tetrastyla | + | |
| 杯水母属Phialidium sp. | + | |
| 拟细浅室水母Lensia subtiloides | + | + |
| 钟浅室水母Lensia campanella | + | |
| 鸟喙尖头溞Penilia avirostris | + | |
| 肥胖三角溞Evadne tergestina | + | |
| 尖尾海萤Cypridina acuminata | + | + |
| 中华哲水蚤Calanus sinicus | + | |
| 强额孔雀水蚤Pavocalanus crassirostris | + | |
| 精致真刺水蚤Euchaeta concinna | + | |
| 矮拟哲水蚤Paracalanus nanus | + | |
| 小拟哲水蚤Paracalanus parvus | + | |
| 微驼隆哲水蚤Acrocalanus gracilis | + | |
| 弓角基齿哲水蚤Clausocalanus arcuicornis | + | |
| 锥形宽水蚤Temora turbinata | + | + |
| 伯氏平头水蚤Candacia bradyi | + | |
| 汤氏长足水蚤Calanopia thompsoni | + | |
| 孔雀唇角水蚤Labidocerapavo | + | |
| 三指角水蚤Pontella tridactyla | + | |
| 红纺锤水蚤Acartia erythraeus | + | |
| 刺尾纺缍水蚤Acartia spinicauda | + | |
| 钳形歪水蚤Tortanus forcipatus | + | |
| 右突歪水蚤Tortanus dextrilobatus | + | |
| 细长腹剑水蚤Oithona affenuatus | + | |
| 近缘大眼剑水蚤Corycaeus affinis | + | |
| 尖额真猛水蚤Euterpina acutifrons | + | + |
| 中华节糠虾Siriella sinensis | + | |
| 圆柱水虱属Cirolana sp. | + | |
| 敏捷蜾蠃蜚Corophium kitamorii | + | |
| 玻璃钩虾属Hyale sp. | + | |
| 角钩虾属Ceradocus sp. | + | |
| 日本毛虾Acetes japonicus | + | |
| 亨生莹虾Lucifer hanseni | + | |
| 箭虫属Sagitta sp. | + | |
| 强壮箭虫内海型Sagitta crass f. naikaiensis | + | |
| 肥胖箭虫Sagitta enflata | + | + |
| 圆囊箭虫Sagitta johorensis | + | |
| 小箭虫Sagitta neglecta | + | |
| 微型箭虫Sagitta minima | + | |
| 太平洋箭虫Sagitta pacifica | + | + |
| 假锯齿箭虫Sagitta pseudoserratodentata | + | |
| 寻觅箭虫Sagitta zetesios | + | + |
| 弱箭虫Sagitta delicata | + | + |
| 大头箭虫Sagitta macrocephala | + | |
| 软拟海樽Dolioletta gegenbauri | + | |
| 腕足类舌贝幼虫Lingula larvae | + | |
| 蔓足类无节幼虫Nauplius larvae (Cirripdia) | + | + |
| 短尾类溞状幼虫Zoea larvae (Brachyura) | + | + |
| 鱼卵Fish eggs | + | + |
| 才女虫幼虫Polydora larvae | + | + |
| 阿利玛幼虫Alima larvae | + | |
| 肠腮类柱头幼虫Enteropneustatornaria | + | |
| 水螅水母类Hydropolypse sp. | + | |
| 肉质介螅水母Hydractinia carnea | + | |
| 短腺和平水母Eirene brevigona | + | |
| 指突水母属Virginic sp. | + | |
| 日本真瘤水母Eutima japonica | + | + |
| 卵形侧丝水母Helgicirrha ovalis | + | |
| 双生水母Diphyes chamissonis | + | |
| 球形侧腕水母Pleurobrachia globosa | + | + |
| 诺氏三角溞Evadne nordmanni | + | |
| 针刺真浮萤Euconchoecia aculeata | + | |
| 异肢水蚤属Copepoda sp. | + | |
| 微刺哲水蚤Canthocalanus pauper | + | |
| 亚强真哲水蚤Eucalanus subcrassus | + | + |
| 狭额真哲水蚤Eucalanus subtenuis | + | |
| 裸拟哲水蚤Delius nudus | + | |
| 驼背隆哲水蚤Acrocalanus gibber | + | |
| 中华矮隆哲水蚤Bestiolina sinica | + | |
| 缘齿厚壳水蚤Scolecithrix nicobarica | + | |
| 瘦尾胸刺水蚤Centropages tenuiremis | + | + |
| 小长足水蚤Calanopia minor | + | |
| 真刺唇角水蚤Labidocera euchaeta | + | |
| 圆唇角水蚤Labidocera rotunda | + | + |
| 克氏纺缍水蚤Acartia clausi | + | |
| 太平洋纺锤水蚤Acartia pacifica | + | + |
| 中华异水蚤Acartiella sinensis | + | |
| 瘦形歪水蚤Tortanus gracilis | + | |
| 捷氏歪水蚤Tortanus derjugini | + | |
| 小长腹剑水蚤Oithona nana | + | |
| 平大眼剑水蚤Corycaeus dahli | + | |
| 分叉小猛水蚤Tisbe furcata | + | |
| 小拟节糠虾Hemisiriella parva | + | |
| 南沙沙钩虾Byblis nanshaensis | + | |
| 上野蜾蠃蜚Corophium uenoi | + | |
| 沙钩虾属Byblis sp. | + | |
| 裂颏蛮绒Lestrigonus schizogeneios | + | |
| 费氏莹虾Lucifer faxonii | + | |
| 中型莹虾Lucifer intermedius | + | + |
| 百陶箭虫Sagitta bedoti | + | + |
| 多变箭虫Saglta decipiens | + | + |
| 凶形箭虫Sagitta ferox | + | + |
| 琴形箭虫Sagitta lyra | + | + |
| 美丽箭虫Sagitta pulchra | + | |
| 拿卡箭虫Sagitta nagae | + | |
| 漂浮箭虫Sagitta planctonis | + | |
| 瘦型箭虫Sagitta tenuis | + | |
| 粗壮箭虫Sagitta robusta | + | |
| 贝勒福箭虫Sagitta bedfordii | + | |
| 异体住囊虫Oikopleura dioica | + | |
| 多毛类疣足幼虫Nectochaeta larvae(Polychacta) | + | |
| 桡足类无节幼虫Nauplius larvae (Copepoda) | + | |
| 长尾类幼虫Macrura larvae | + | + |
| 磁蟹溞状幼虫Zoea larvae (Porcellana) | + | |
| 仔鱼Fish larvae | + | |
| 蛇尾长腕幼虫Ophiopluteus larvae | + | |
| 真寄居虾后期幼虫Eupagurus post larvae | + | |
| 肠腮类柱头幼虫Enteropneustatornaria | + | |
| “+”表示该季节有出现 | ||
| 浮游动物类群 Zooplankton group |
秋季Autumn | 春季Spring | 总计Total | 占比Percentage/% |
| 桡足类Copepoda | 31 | 14 | 39 | 35.14 |
| 毛颚类Chaetognatha | 18 | 11 | 21 | 18.92 |
| 浮游幼虫Pelagic larvae | 14 | 5 | 14 | 12.61 |
| 水螅水母类Hydromedusae | 6 | 6 | 11 | 9.91 |
| 端足类Amphipoda | 4 | 3 | 7 | 6.31 |
| 樱虾类Sergestes | 4 | 1 | 4 | 3.60 |
| 枝角类Cladocera | 3 | 0 | 3 | 2.70 |
| 糠虾类Mysidacea | 2 | 0 | 2 | 1.80 |
| 管水母类Siphonophora | 1 | 3 | 3 | 2.70 |
| 栉水母类Ctenophora | 1 | 1 | 1 | 0.90 |
| 介形类Ostracocda | 1 | 2 | 2 | 1.80 |
| 被囊类Tunicata | 1 | 1 | 2 | 1.80 |
| 原生动物Protozoa | 0 | 1 | 1 | 0.90 |
| 等足类Isopod | 1 | 0 | 1 | 0.90 |
| 合计Total | 87 | 48 | 111 | 100 |
秋季, 各调查站位浮游动物的丰度变化范围为14.00—2320.63个/m3, 均值为528.92个/m3, 呈现外湾沿岸向中部水域以及内湾逐渐减少的分布趋势(图 3), 其中内湾和外湾的丰度均值分别为59.22、822.49个/m3, 内湾浮游动物丰度均值远远低于外湾。春季浮游动物丰度的变化范围为1.00—218.75个/m3, 均值为48.30个/m3, 外湾中部海域的丰度较高, 内湾河口附近丰度偏低, 呈现出由外湾向内湾递减的分布趋势, 其中内湾和外湾的丰度均值分别为11.70、65.63个/m3, 春季内湾丰度明显低于外湾。

|
| 图 3 秋季和春季浮游动物丰度水平分布/(个/m3) Fig. 3 Horizontal distribution of abundance of zooplankton in the Qinzhou Bay in autumn and spring |
钦州湾浮游动物优势种共有6种(表 3), 秋季和春季各为5种和2种, 其中太平洋纺锤水蚤为两季共有的优势种。秋季以太平洋纺锤水蚤为第一优势种群, 如图 4所示, 除Q13外其余站位均有分布, 调查海域的平均丰度为123.03个/m3, 其中外湾东侧金鼓江口Q9站位的丰度最高, 达645.00个/m3;优势度居次的是长尾类幼虫, 主要分布在外湾, 其中以Q11站位的丰度最高。春季中华哲水蚤的优势度最高, 但分布范围较小, 仅出现在外湾的Q9、Q10、Q12和Q13站位;优势度居次的是太平洋纺锤水蚤, 主要分布在内湾和外湾湾顶附近的站位(图 4)。
| 时间 Time |
优势种 Dominance species |
优势度Y Degree of dominance |
平均丰度 Averageabundance/(个/m3) |
占比 Percentage/% |
| 春季Spring | 中华哲水蚤Calanus sinicus | 0.151 | 23.66 | 49.02 |
| 太平洋纺锤水蚤Acartia pacifica | 0.031 | 2.42 | 5.01 | |
| 秋季Autumn | 太平洋纺锤水蚤Acartia pacifica | 0.215 | 123.03 | 23.26 |
| 长尾类幼虫Macrura larvae | 0.181 | 138.10 | 26.11 | |
| 肥胖三角溞Evadne tergestina | 0.059 | 67.64 | 12.79 | |
| 亨生莹虾Lucifer hanseni | 0.058 | 40.11 | 7.58 | |
| 百陶箭虫Sagitta bedoti | 0.030 | 25.48 | 4.82 |

|
| 图 4 秋季和春季主要优势种丰度的水平分布/(个/m3) Fig. 4 Horizontal distribution of abundance of dominant species in the Qinzhou Bay in autumn and spring |
秋季浮游动物生物量的变化范围为18.67—476.90 mg/m3, 均值为110.60 mg/m3, 生物量较高的区域主要分布在外湾, 生物量分布趋势呈现由外湾湾顶水域向四周逐渐降低, 内湾和外湾生物量的均值分别为26.39、163.23 mg/m3, 内湾生物量均值远低于外湾(图 5)。春季浮游动物生物量的变化范围为3.46—187.52 mg/m3, 均值为68.64 mg/m3, 平面分布整体呈外湾中部水域向内湾递减, 内湾和外湾生物量的均值分别为28.20、93.92 mg/m3, 内湾生物量明显低于外湾(图 5)。

|
| 图 5 秋季和春季浮游动物生物量水平分布/(mg/m3) Fig. 5 Horizontal distribution of biomass of zooplankton in the Qinzhou Bay in autumn and spring |
秋季, 各调查站位浮游动物的多样性指数波动于0.37—3.61之间, 均值为2.22。其中内湾各站位多样性指数均值为1.20, 外湾多样性指数均值为2.86, 外湾多样性指数高于内湾。多样性指数较高的站位主要分布于外海海域(图 6)。春季, 各测站浮游动物多样性指数变化幅度为0—3.38, 均值为1.70。内湾各站位的多样性指数较低, 均值为1.10, 外湾则相对较高, 均值为2.10。多样性较高的站位主要位于外湾西南部海域(图 6)。

|
| 图 6 钦州湾秋季和春季浮游动物多样性的水平分布 Fig. 6 Horizontal distribution of diversity index of zooplankton in the Qinzhou Bay in autumn and spring |
钦州湾秋季和春季不同站位的浮游动物物种丰度多维尺度分析结果如图 7所示。秋季, 以40%的群落相似性将13个站位的浮游动物群落分为组Ⅰ—组Ⅶ共7组(图 8), 各区域浮游动物群落结构差异较大。其中组Ⅰ包括了内湾以及湾颈的大部分站位, 各站位间的相似度高达71.23%, 均以太平洋纺锤水蚤为主, 但种类和丰度相对较低。春季, 浮游动物群落分为组Ⅰ—组Ⅷ共8组, 其中组Ⅷ包括了外湾的Q10、Q12和Q13共3个站位, 各站位间的相似度高达60.98%, 均以中华哲水蚤为优势类群, 且丰度也较高。总体而言, 钦州湾秋季浮游动物群落为内湾区域相似性较高, 春季为外湾区域相似性较高。

|
| 图 7 各站位浮游动物群落结构的多维尺度分析 Fig. 7 Multidimensional scaling (MDS) of different zooplankton samples |

|
| 图 8 钦州湾秋季和春季浮游动物群落结构类型以及分布示意图 Fig. 8 Clusters and distributions of zooplankton community in the Qinzhou bay in autumn and spring |
对可能影响钦州湾浮游动物群落理化因子进行的主成分分析(PCA), 除材料与方法中提到的理化因子, 其余用于PCA分析的理化因子(透明度、溶解氧、化学需氧量、pH、铜、铅、锌、铬和镍)来源于广西海洋环境监测中心站2014年至2015年的监测数据。PCA的结果显示第一主成分涉及的因素有硅酸盐(0.990)、无机氮(0.945)、盐度(0.942)、磷酸盐(0.903)。第二主成分涉及的因素有叶绿素a(0.903)和水温(0.795)。因此, 选取硅酸盐、无机氮、叶绿素a、水温、盐度和磷酸盐作为主要环境因子。上述主要环境因子与浮游动物丰度相关性分析结果如表 4所示。秋季钦州湾的浮游动物丰度与盐度显著正相关(P < 0.05), 与无机氮、硅酸盐和磷酸盐显著负相关关系(P < 0.05);春季则与水温、盐度和叶绿素a显著正相关(P < 0.05), 与无机氮和硅酸盐显著负相关(P < 0.05)。
| 环境因子 Environmental factors |
水温 Temperature |
盐度 Salinity |
叶绿素a Chl-a |
无机氮 Dissolved inorganic nitrogen |
硅酸盐 SiO32--Si |
磷酸盐 PO43--P |
| 秋季Autumn | 0.468 | 0.579* | 0.051 | -0.645* | -.669* | -0.645* |
| 春季Spring | 0.584* | 0.573* | 0.598* | -0.563* | -.607* | -0.43 |
| *:P < 0.05, 显著相关 | ||||||
本次调查发现钦州湾秋季和春季浮游动物的种类组成存在明显的季节变化, 从秋季到春季浮游动物种类组成由暖水性向暖温性种类更替, 优势种也发生了明显的季节演替。秋季浮游动物优势种的种类数较多, 主要有太平洋纺锤水蚤、长尾类幼虫、肥胖三角溞、亨生莹虾和百陶箭虫(表 3);春季浮游动物种类较少, 优势种只有中华哲水蚤和太平洋纺锤水蚤。太平洋纺锤水蚤为钦州湾秋季和春季共同的优势种类, 该物种为典型的河口沿岸种类[6]。钦州湾地处钦江、茅岭江等江河的入海口, 海水盐度相对偏低, 适宜太平洋纺锤水蚤生长。秋季, 随着径流量的增加以及水温的上升, 太平洋纺锤水蚤大量繁殖成为秋季第一优势种群, 从内湾河口区至外湾均有分布。而随着水温降低及盐度增高, 春季太平洋纺锤水蚤虽然在本次调查中居第二优势种群, 但优势度很低, 而且主要出现在内湾以及外湾湾顶等盐度较低的海域, 这表明了太平洋纺锤水蚤的分布与盐度密切相关。秋季地表径流影响范围的扩大可能是导致多维尺度分析中内湾多数站位群落之间无较大差异的主要原因。此外, 通过比较2012年3月与2015年3月的环境因子, 发现2012年钦州湾内湾的水温(< 15℃)[2]明显低于2015年的(>17℃, 图 2), 这直接导致了2012年3月的太平洋纺锤蚤的分布范围较小[2], 同时也表明了水温是决定其分布的另一个重要因子。中华哲水蚤是适应较低温度的近岸暖温种类[7], 温度被认为是影响中华哲水蚤群落分布的重要环境因素[8-9]。钦州湾地处亚热带, 冬春季水温偏低, 夏秋季水温较高, 因此中华哲水蚤成为本次春季调查的绝对优势种。除了水温之外, 盐度也是决定中华哲水蚤分布的重要决定要素, 其最适盐度为30左右[10], 这导致其只分布在外湾盐度较高的海域。此外春季地表径流量减少, 沿岸水势力减弱, 外海水势力向湾内推进作用增强, 中华哲水蚤随着外海水进入钦州湾, 引起春季钦州湾多维尺度分析中外湾浮游动物群落相似性较高。钦州湾浮游动物主要优势种从秋季高温低盐的太平洋纺锤水蚤向春季低温高盐的中华哲水蚤演替, 说明了该海域主要受到江河淡水输入和水温季节变化的影响, 也反映了钦州湾浮游动物的亚热带河口海湾群落结构特性。
3.2 钦州湾浮游动物分布的主要影响因素钦州湾受到茅岭江、钦江等大陆径流及北部湾外海水的综合影响, 环境变化剧烈。由图 2可以看出, 钦州湾盐度、无机氮、硅酸盐及磷酸盐的变化特征均呈现出明显的河口特性, 浮游动物丰度、生物量与上述因子的相关性分析结果也显示浮游动物分布受到了江河淡水输入的影响。秋季钦州湾沿岸水温与外海基本相同, 水温较高(28.3℃)且均匀, 与浮游动物丰度、生物量分布的关系较小。春季, 钦州湾水温平面分布差异较大, 呈现从河口向外湾递增的趋势, 恰与浮游动物的分布特征相似, 相关性分析也表明二者呈显著正相关。盐度是影响中华哲水蚤和太平洋纺锤水蚤生长的重要因素, 而两者作为春季和秋季浮游动物的主要优势种, 直接影响了浮游动物丰度的分布。浮游植物是浮游动物的重要饵料, 浮游动物丰度与叶绿素a的正相关关系可能与浮游动物的选择性摄食有关[11]。大多数以浮游植物为食的浮游动物与叶绿素a含量具有正相关, 而以其他浮游动物为食的次级消费者(毛颚类、水母类等浮游动物)与叶绿素a的相关性不明显[12]。春季外湾叶绿素a浓度较高的区域恰好也是中华哲水蚤的分布区, 这与中华哲水蚤营植食性的特性一致[13]。从无机氮、硅酸盐及磷酸盐的变化中可以看出, 高浓度的营养盐与低盐度有较好的吻合关系, 表明径流输入对海湾营养盐变化起着主导作用。一般来说, 营养盐丰富的水域有利于浮游生物的生长, 但钦州湾内湾浮游动物丰度较低, 这可能跟营养盐结构比例失调有关:钦州湾长期处于磷限制状态[14], 即使在营养盐充沛的条件下浮游植物也无法大量生长繁殖, 从而不能给更多的浮游动物生长提供食物来源, 这可能是导致内湾浮游动物的丰度较低的原因。综上所述, 江河淡水输入(盐度和营养盐)是决定钦州湾浮游动物平面分布的最主要因素。
3.3 钦州湾浮游动物群落结构及生物量的年际变化特征2011—2012年钦州湾调查结果显示(表 5), 夏季和春季的第一优势种与本次调查秋季和春季的第一优势种一致, 但优势种的丰度存在较大差异。2015年春季第一优势种的丰度显著低于2012年春季, 而2014年秋季第一优势种则明显高于2011年夏季。此外, 2011年夏季和2012年春季的第二优势种均为个体大小为5—12 mm的球形侧腕水母(Pleurobrachia globosa), 而本次调查秋季和春季的第二优势种分别是个体大小仅1.0—1.5 mm的长尾类幼虫和太平洋纺锤水蚤, 可见决定浮游动物生物量大小的优势种个体已趋向小型化, 造成尽管此次调查浮游动物的丰度有所上升, 但生物量明显下降。而这一现象在内湾尤为突出, 秋季和春季内湾均以太平洋纺锤水蚤为唯一优势种, 优势种群趋于单一化, 因此这些因素直接引起了内湾和外湾浮游动物丰度和生物量的较大差异。
| 调查时间 Investigatedtime |
平均丰度 Averageabundance/(个/m3) |
平均生物量 Average biomass/(mg/m3) |
夏/秋季主要优势种和平均丰度 Dominant species in summer/autumn and average abundance |
春季主要优势种和平均丰度 Dominant species in springand average biomass |
文献来源 Literaturesource |
| 2011—2012 | 161.55 | 259.45 | 太平洋纺锤水蚤(12.80个/m3)、球形侧腕水母 | 中华哲水蚤(76.00个/m3)、球形侧腕水母 | [2] |
| 2014—2015 | 288.61 | 89.62 | 太平洋纺锤水蚤(123.03个/m3)、长尾类幼虫 | 中华哲水蚤(23.66个/m3)、太平洋纺锤水蚤 | 本次调查 |
2011—2015年5年间, 钦州湾浮游动物群落结构发生了一定的变化, 浮游动物种类趋于小型化、单一化, 导致生物量呈下降的趋势。经初步分析, 其原因可能有以下几个方面:
(1) 钦州湾湾颈及内湾海域高密集的牡蛎养殖相当于浮游植物过滤器:涨潮时外湾较高浮游植物生物量的水体在输送到内湾的过程中浮游植物被牡蛎大量摄食, 导致了内湾浮游植物优势类群以蓝藻和青绿藻等超微型类群为主, 外湾则以硅藻等小型和微型浮游植物为优势类群[2]。内湾粒级较小的浮游植物只能维持小型浮游动物的生长需求, 而个体较大的浮游动物则更偏向外湾摄食粒级较大的硅藻食物, 这可能是造成内湾浮游动物的丰度和生物量偏低的主要原因。近年来钦州湾牡蛎养殖区域从内湾扩大至外湾, 养殖面积不断增加, 至2015年养殖面积约为100 hm2[15], 几乎与内湾茅尾海面积相当。高密度牡蛎养殖规模的扩大对钦州湾浮游动物群落结构分布造成了一定的影响。
(2) 钦州湾高密度大面积养殖的牡蛎不仅通过滤食浮游植物与浮游动物形成食物竞争关系, 还会滤食相当一部分浮游动物的幼体、桡足类成体甚至是鱼卵和仔、稚鱼[16-17], 导致浮游动物的数量下降, 从而对浮游动物群落结构产生一定的影响。尽管钦州湾牡蛎滤食浮游动物的种类和数量尚未有相关报道, 但在本次调查中发现浮游动物幼体主要集中在外湾这一现象可以进一步反映牡蛎养殖给浮游动物带来的影响。值得注意的是, 太平洋纺锤水蚤主要分布区域恰好也是牡蛎养殖区, 这是否与牡蛎有选择性的滤食有关抑或是太平洋纺锤水蚤可以避开牡蛎的滤食作用, 将有待进一步研究。
(3) 2015年春季(3月)内湾水温与2012年同期相比高了2℃以上, 钦州湾海域水温升高可能与全球气候变暖现象有关。气候变暖导致的水温升高会对浮游动物群落结构造成一定的影响。在全球气候变暖这一环境背景下, 随着钦州湾燃煤电厂以及核电站的建设和运行, 温排水的排放无疑会加剧水温升高给浮游动物带来的影响。有研究表明温排水会降低排水口附近海域浮游动物的种类、丰度及多样性[18], 并且温排水对大型浮游动物种类的分布有较大影响, 而对中小型浮游动物种类分布几乎没有影响[19]。尽管目前钦州湾还没有相关的研究报道, 但温排水对钦州湾海洋生态的影响仍需引起关注。
(4) 钦州湾湾颈狭窄, 加上多年来大规模围填海建设以及内湾和湾颈密集的牡蛎养殖严重降低了水体交换率, 营养物质不易扩散, 导致内湾常年处于富营养化状态[20]。近几年的数据显示内湾富营养化程度日渐加剧[14]。营养盐的持续输入改变水体中营养盐的组成和比率。许多研究表明, 水体富营养化的加剧和营养盐结构的变化已使得近岸海域浮游植物群落构成趋于小型化[21-22], 并通过食物链的传递效应, 进一步引起主要摄食者浮游动物的小型化[23], 这与本次研究结果一致。水体富营养化很可能是促进钦州湾浮游动物群落结构单一化和小型化的原因之一。
4 结语钦州湾日益频繁的开发活动导致浮游动物群落结构发生了一定的变化。其中内湾受到的影响较大, 其浮游动物群落组成较为简单, 种类趋于小型化, 群落结构较为脆弱, 如不加以遏止, 最终会给整个钦州湾海洋生态系统的稳定和健康带来威胁。今后需对钦州湾的海洋资源进行合理开发以及加强对水质的监控和环境管理, 才能保持该海域生态环境可持续发展。
| [1] | 蓝文陆, 杨绍美, 苏伟. 环钦州湾河流入海污染物通量及其对海水生态环境的影响. 广西科学, 2012, 19(3): 257–262. DOI:10.3969/j.issn.1005-9164.2012.03.016 |
| [2] | 蓝文陆, 李天深, 刘勐伶, 庞碧剑, 刘昕明. 钦州湾丰水期和枯水期浮游动物群落特征. 海洋学报, 2015, 37(4): 124–132. DOI:10.3969/j.issn.0253-4193.2015.04.012 |
| [3] | 中华人民共和国国家质量监督检验检疫总局, 国家标准化管理委员会. GB17378.7-2007.海洋监测规范第7部分: 近海污染生态调查和生物监测.北京: 中国标准出版社, 2008. |
| [4] | 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13-15, 19-19. |
| [5] | 徐兆礼, 王云龙, 陈亚瞿, 沈焕庭. 长江口最大浑浊带区浮游动物的生态研究. 中国水产科学, 1995, 2(1): 39–48. DOI:10.3321/j.issn:1005-8737.1995.01.002 |
| [6] | 高亚辉, 林波. 几种因素对太平洋纺锤水蚤摄食率的影响. 厦门大学学报:自然科学版, 1999, 38(5): 751–757. |
| [7] | 陈清潮. 中华哲水蚤的繁殖、性比率和个体大小的研究. 海洋与湖沼, 1964, 6(3): 272–288. |
| [8] | 曹文清, 林元烧, 杨青, 李少菁. 我国中华哲水蚤生物学研究进展. 厦门大学学报:自然科学版, 2006, 45(S2): 54–61. |
| [9] | Wang R, Zuo T, Wang K. The Yellow Sea Cold Bottom Water an oversummering site for Calanussinicus (Copepoda, Crustacea). Journal of Plankton Research, 2003, 25(2): 169–183. DOI:10.1093/plankt/25.2.169 |
| [10] | 黄加祺, 郑重. 温度和盐度对厦门港几种桡足类存活率的影响. 海洋与湖沼, 1986, 17(2): 161–167. |
| [11] | 杜萍, 徐晓群, 刘晶晶, 江志兵, 陈全震, 曾江宁. 象山港春、夏季大中型浮游动物空间异质性. 生态学报, 2015, 35(7): 2308–2321. |
| [12] | 骆鑫, 曾江宁, 徐晓群, 杜萍, 廖一波, 刘晶晶. 舟山海域夏、秋季浮游动物的分布特征及其与环境因子的关系. 生态学报, 2016, 36(24): 8194–8204. |
| [13] | 杨纪明. 渤海中华哲水蚤摄食的初步研究. 海洋与湖沼, 1997, 28(4): 376–382. DOI:10.3321/j.issn:0029-814X.1997.04.007 |
| [14] | 蓝文陆. 钦州湾枯水期富营养化评价及其近5年变化趋势. 中国环境监测, 2012, 28(5): 40–44. DOI:10.3969/j.issn.1002-6002.2012.05.009 |
| [15] | 钦州市人民政府. 钦州年鉴2016. 南宁: 广西人民出版社, 2016. |
| [16] | Gibbs M T. Interactions between bivalve shellfish farms and fishery resources. Aquaculture, 2004, 240(1/4): 267–296. |
| [17] | Davenport J, Smith R J J W, Packer M. Mussels Mytilus edulis:significant consumers and destroyers of mesozooplankton. Marine Ecology Progress Series, 2000, 198: 131–137. DOI:10.3354/meps198131 |
| [18] | 傅强, 朱礼鑫, 沈盎绿, 李道季. 滨海电厂温排水对临近海域浮游动物生态特征的影响研究. 中国海洋大学学报:自然科学版, 2015, 45(7): 25–33. |
| [19] | 徐晓群, 曾江宁, 曾淦宁, 陈全震, 廖一波, 高爱根, 寿鹿. 滨海电厂温排水对浮游动物分布的影响. 生态学杂志, 2008, 27(6): 933–939. |
| [20] | 闭文妮, 唐艳葵, 洪鸣, 顾湛琦. 钦州湾无机氮入海通量及其富营养化症状分析. 环境科学与技术, 2014, 37(1): 174–178. |
| [21] | Pan Y L, Rao D V S. Impacts of domestic sewage effluent on Phytoplankton from Bedford Basin, eastern Canada. Marine Pollution Bulletin, 1997, 34(12): 1001–1005. DOI:10.1016/S0025-326X(97)00115-X |
| [22] | Dippner J W. Competition between different groups of phytoplankton for nutrients in the Southern North Sea. Journal of Marine Systems, 1998, 14(1/2): 181–198. |
| [23] | 张才学, 龚玉艳, 王学锋, 孙省利. 湛江港湾浮游桡足类群落结构的季节变化和影响因素. 生态学报, 2011, 31(23): 7086–7096. |