 2018, Vol. 38
2018, Vol. 38文章信息
- 范小晨, 代存芳, 陆欣鑫, 范亚文.
- FAN Xiaochen, DAI Cunfang, LU Xinxin, FAN Yawen.
- 金河湾城市湿地浮游植物功能类群演替及驱动因子
- Study on phytoplankton functional group succession and driving parameters in the Jinhewan Urban Wetland
- 生态学报. 2018, 38(16): 5726-5738
- Acta Ecologica Sinica. 2018, 38(16): 5726-5738
- http://dx.doi.org/10.5846/stxb201710251916
-
文章历史
- 收稿日期: 2017-10-25
- 修订日期: 2018-05-30

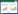




浮游植物在湿地生态系统中占据关键作用, 它可以利用光能驱动光合作用产生有机物, 是水生生物食物链的基础环节, 促进了能量在食物链中的传递, 最终影响整个湿地水域系统的稳定[1]。浮游植物的时空分布模式是探索湿地生态系统功能的重要手段之一, 它不仅影响了湿地生态过程、功能和稳定性, 而且反映了湿地生态环境的变化[2-3]。目前, 浮游植物的生态分布在湿地生态系统的相关研究中, 主要集中在不同环境梯度和不同生活类型下浮游植物群落结构的差异性分析[4-6]。这类研究多使用传统的浮游植物集群方法, 不能有效地反映浮游植物在湿地生态系统中的功能。2002年, Reynolds等提出基于浮游植物的功能类群的概念[7], 随后Padisák等[8]补充完善了功能类群的划分及生态特性, 最终形成了探索浮游植物生态过程的重要理论[9]。浮游植物功能类群分类法以浮游植物的功能性特征为基础(包括生理学、形态学和生态学特征), 将同一生境下共存的藻类归为一组, 同组内的浮游植物通常具有相似的环境适应性特征[8]。关于浮游植物功能类群生理和生态的相关研究已成为当今水域生态学的热点问题, 目前已经广泛应用于湖泊、水库等生态系统的研究与保护中[10-12]。
城市湿地公园作为湿地生态恢复、湿地资源可持续利用以及湿地保护的有机结合体, 是对《湿地公约》提倡合理利用湿地资源原则的充分体现和具体实施[13]。城市湿地公园对于改善城市气候、维护区域生态平衡、调节区域小气候以及优化城市景观格局等方面具有重要的生态作用, 另外城市湿地公园也是众多湿地保护和利用模式中的一种较为理想的模式之一[14]。目前对于城市湿地公园的生态研究, 已经成为城市生态建设的重点和研究的热点之一[15-16]。金河湾湿地公园位于松花江哈尔滨段主城区左岸, 西四方台大桥与阳明滩大桥之间的滩地上, 东西长3800 m, 南北最宽处1500 m, 总面积1.8 km2。金河湾湿地对哈尔滨市保护生物多样性、调节地区小气候等方面有着重要作用[17]。目前, 有关金河湾湿地的研究多为景观体验优化策略、规划设计、POE调查等[17-18]。关于金河湾湿地浮游植物生态多样性的研究较少, 且研究内容采用传统分类方法对浮游植物群落结构进行探讨[19]。
本研究将浮游植物功能类群运用于金河湾湿地浮游植物群落结构研究中, 阐释了不同功能类群在季节间的演替特征, 同时探究影响浮游植物在群落演替过程中生长选择机制的主要环境变量。以期为金河湾湿地浮游植物的多样性保护提供重要的生态学基础资料, 同时为金河湾湿地公园的生态管理提供理论依据。
1 材料与方法 1.1 采样点设置我们于2015年春(5月)、夏(8月)、秋(10月)三季对金河湾湿地公园进行水样及浮游植物样品采集。根据金河湾湿地的水体生态特征(与河道相通程度、水深、湿生植物等), 本研究共设置12个采样点(图 1), 基本涵盖了金河湾湿地的4种典型水体; 样点S1—S7的生境类型为湖泊型湿地, 采集地水体为较开阔的湖面; 样点S8—S10为河流型湿地, 所在水域与松花江河道相通; 样点S11为沼泽湿地, 有薄层常年积水, 水面生长大面积喜湿性草本植物; 样点S12为人工湿地, 是金河湾湿地植物园打造的圆形水池, 平均水深1 m。采样点的设置定位采用GPS全球卫星定位系统。采样点地图的绘制通过ArcMap 10.2完成。

|
| 图 1 金河湾湿地采样点布设示意图 Fig. 1 Map and sampling sites in Jin Hewan wetland 样点S1—S7的生境类型为湖泊型湿地, 采集地水体为较开阔的湖面; 样点S8—S10为河流型湿地, 所在水域与松花江河道相通; 样点S11为沼泽湿地, 有薄层常年积水, 水面生长大面积喜湿性草本植物; 样点S12为人工湿地 |
根据常规方法对浮游植物标本进行定量、定性采集[20], 现场对采集样本用鲁格试剂固定, 静止24 h后浓缩至50 mL, 镜检计数[21]。浮游植物的鉴定及生态分布信息主要参考胡鸿钧、魏印心等文献[21-23]。
1.3 理化指标的测定使用pH水温测试仪现场测定水温(WT)和pH; 电导率(SpCond); 溶解氧(DO)、氧化还原电位(ORP)、浊度(Tur)以及叶绿素a(Chl-a)用Hydrolab多参数水质分析仪进行测定; 采集现场用黑色密封袋将水样密封保存, 12小时内使用DRB200型数字反应器对总磷(TP)、总氮(TN)及化学需氧量(COD)进行测定。
1.4 数据分析独立样本t检验(t-test)和双因素方差分析(Two-way ANOVA)使用SPSS 22.0软件完成; 使用PRIMER 5.0软件进行单因素交叉相似性分析(One-way crossed ANOSIM)和SIMPER分析。去趋势分析(DCA)和典范对应分析(CCA)采用软件CANOCO 4.5进行, 在进行排序前, 对解释变量和响应变量数据进行log(x+1)转换, 使样本总体趋于正态分布。通过蒙特卡洛置换检验, 判断解释变量对响应变量生态分布影响的显著性, 建立二维排序图。
2 结果与分析 2.1 金河湾湿地浮游植物群落结构 2.1.1 浮游植物种类组成在研究期间共鉴定浮游植物376个种, 包括32变种2变型, 隶属于8门10纲19目39科101属。硅藻门(Bacillariophyta)种类数最多, 为149种及变种2变型, 占浮游植物总种类数的39.62%;绿藻门(Chlorophyta)134种及变种, 占35.64%;裸藻门(Euglenophyta) 38种及变种, 占10.11%;蓝藻门(Cyanophyta)37种及变种, 占9.84%;甲藻门(Dinophyta)10种及变种, 占2.66%;隐藻门(Cryptophyta)4种及变种, 占1.06%;黄藻门(Xanthophyta)3种; 占0.79%;金藻门(Chrysophyta)仅1种, 占0.26%(图 2)。主要优势种包括硅藻门的谷皮菱形藻(Nitzschia palea)、梅尼小环藻(Cyclotella meneghiniana); 绿藻门的具尾四角藻(Tetraëdron caudatum)、螺旋弓形藻(Schroederia spiralis)、四尾栅藻(Scenedesmus quadricauda); 蓝藻门的固氮鱼腥藻(Anabaena azotica)、中华小尖头藻(Raphidiopsis sinensia)、水华束丝藻(Aphanizomenon flos-aquae); 隐藻门的卵形隐藻(Cryptomonas ovata)、啮噬隐藻(Cryptomonas erosa)以及金藻门的分歧锥囊藻(Dinobryon divergens)。

|
| 图 2 金河湾湿地浮游植物种类组成 Fig. 2 The composition of phytoplankton taxa in Jin Hewan wetland |
研究期间, 整体上呈现夏季>秋季>春季的规律, 经独立样本t检验, 季节间丰度差异显著(P < 0.05)。其中, 夏季平均丰度最高为12.90×106个/L, 变化范围在0.47—30.17×106个/L之间, 其次为秋季, 平均丰度为5.95×106个/L, 变化范围在0.97—14.83×106个/L之间, 春季平均丰度最低为2.55×106个/L, 变化范围在0.23—6.12×106个/L之间。从空间分布来看, 湖泊型湿地藻类丰度显著高于其他类型的湿地(P < 0.05), 而河流型湿地、沼泽湿地和人工湿地的采样点中藻类丰度相对较低(如图 3)。如, 春季丰度最高值出现在样点S4(6.12×106个/L), 夏、秋两季丰度最高值都出现在样点S3, 丰度分别为30.17×106个/L和14.83×106个/L。春、夏两季浮游植物丰度最低值均出现在沼泽湿地样点S11, 丰度分别为0.23×106个/L和0.47×106个/L。秋季藻类丰度最低值为河流型湿地样点S8(0.97×106个/L)。

|
| 图 3 金河湾湿地各样点在春、夏、秋三季的浮游植物丰度 Fig. 3 The abundance of phytoplankton in Jin Hewan wetland sites |
根据Reynolds等[7]和Padisák等[8]对浮游植物功能类群的划分标准, 金河湾湿地浮游植物群落可划分为20个不同的功能类群, 分别为B/C/D/E/F/H1/H2/J/Lo/LM/MP/N/P/S1/S2/SN/W1/W2/X1/Y(表 1)。浮游植物功能类群出现频率分布如图 4所示, J/Lo/X1/P/H1/W2/MP/D/F/W1在四种水体中出现频率较高, 均为50%以上; LM/B/N/S2出现频率相对较低, 在30%—50%之间, Y/SN/H2/C/E/S1出现频率低于30%, 其中E/S1仅在人工湿地、沼泽湿地中被检测出。表 1给出了本研究中各浮游植物功能群的代表性藻种(属)。
| 功能类群 Groups |
代表性藻种(属) Genus/Species Represent in the groups |
耐受性 Tolerance |
敏感性 Sensitive |
| B | 小环藻属 Cyclotella Kütz. | 低光照 | pH升高、硅元素缺乏、水体分层 |
| C | 华丽星杆藻 Asterionella formosa | 低光照、低含碳量 | 硅消耗、水体分 |
| D | 菱形藻属 Nitzschia Hass. | 冲刷 | 营养缺乏 |
| 针杆藻属 Synedra Ehr. | |||
| E | 分歧锥囊藻 Dinobryon divergens | 低营养 | 二氧化碳缺乏 |
| F | 蹄形藻 Kirchneriella lunaris | 低营养、高浑浊 | 二氧化碳缺乏 |
| 网球藻 Dictyosphaerium ehrenbergianum | |||
| H1 | 鱼腥藻属 Anabaena Bory | 低含氮量、低含碳量 | 水体混合、低光照、低含磷量 |
| 水华束丝藻 Aphanizomenon flos-aquae | |||
| H2 | 项圈藻 Anabaenopsis sp. | 低含氮量、低含碳量 | 水体混合、低光照 |
| J | 栅藻属 Scenedesmus Meyen | 高光照 | |
| 四角藻属 Tetraedrom Kütz. | |||
| Lo | 色球藻属 Chroococcus Näg. | 营养分层 | 长时间或深层的混合 |
| 平裂藻属 Merismopedia Meyen | |||
| LM | 裸甲藻 Gymnodinium aeruginosum | 很低的碳含量 | 水体混合、低光照 |
| 角甲藻 Ceratium hirundinella | |||
| MP | 舟形藻属 Navicula Bory | 混合扰动 | |
| 异极藻属 Gomphonema Ehr. | |||
| N | 鼓藻属 Cosmarium Corda | 冲刷 | |
| P | 角星鼓藻属 Staurastrum Meyen | 中程度的低光照和低碳含量 | 水体分层、硅元素缺乏 |
| 颗粒直链藻 Melosira granulata | |||
| S1 | 鞘藻属 Oedogonium Link | 极低的光照 | 冲刷 |
| S2 | 螺旋藻属 Spirulina Turp | 低光照 | 冲刷 |
| SN | 中华小尖头藻 Raphidiopsis sinensia | 低光照和低营养 | 冲刷 |
| W1 | 裸藻 Euglena sp. | 高生化需氧量 | 牧食作用 |
| W2 | 囊裸藻属Trachelomonas Ehr. | 酸性 | |
| X1 | 纤维藻属 Ankistrodesmus Corda | 分层 | 营养盐缺乏、滤食作用 |
| 螺旋弓形藻 Schroederia spiralis | |||
| Y | 啮蚀隐藻 Cryptomonas erosa | 低光照 | 吞噬作用 |
| 卵形隐藻 Cryptomonas ovata |

|
| 图 4 金河湾湿地浮游植物功能类群出现频率 Fig. 4 Frequency distribution of functional groups in Jin Hewan wetland |
根据浮游植物代表性功能类群的划定标准, 至少在一个采样点相对丰度超过10%的功能类群为代表性功能类群[24]。在本次研究中, 共划分12个代表性功能类群, 分别为B/D/F/H1/J/Lo/MP/SN/S1/S2/X1/Y。春季代表性功能类群有7个, 包括B/D/H1/Lo/MP/X1/Y; 夏季代表性功能类群有9个, 分别为B/D/H1/Lo/S1/S2/SN/MP/Y; 秋季代表性功能类群共8个, 分别为B/D/F/H1/J/MP/S2/X1(图 5)。

|
| 图 5 春、夏、秋各水体优势功能类群相对丰度 Fig. 5 Relative abundance of representative functional groups in different sites |
双因素方差分析(Two-way ANOVA)表明, 研究期间金河湾湿地代表性功能类群中B/H1/Lo/S2/X1的丰度随着季节的变化存在显著差异(P < 0.05), 各水体代表性功能类群随季节变化呈现明显的演替现象(表 2)。通过对代表性功能类群的单因子交叉相似性分析(One-way crossed ANOSIM)显示季节变化对金河湾湿地浮游植物功能类群的生态分布具有极其显著的影响(P < 0.01)(表 3)。湖泊型湿地浮游植物功能类群季节性分布为:春季X1/B/Lo/D, 夏季S2/SN/Lo, 秋季H1/S2/B, 其演替表现为春季代表性功能类群X1(狭形纤维藻、螺旋弓形藻)/B(梅尼小环藻)演替为夏季绝对优势类群S2(螺旋藻)/SN(中华小尖头藻)再到秋季主要功能类群H1(水华束丝藻)/S2(螺旋藻)/B(小环藻属)。河流型湿地浮游植物功能类群季节性分布为:春季X1/B/D, 夏季S2/H1/D/B, 秋季B/X1/F, 其中春季优势类群X1(狭形纤维藻)/B(梅尼小环藻), 夏季为H1(水华束丝藻)/B(小环藻属), 最后秋季B(小环藻属)/X1(狭形纤维藻)又重新回到优势地位, B(小环藻属)成为3个季节中的优势类群。沼泽湿地浮游植物功能类群季节性分布为:春季D/MP/Y, 夏季MP/Y/S1/D, 秋季MP/D/B, D(菱形藻属)/MP(舟形藻属)在该水体的3个季节中都为优势类群。人工湿地浮游植物功能类群季节性分布为:春季D, 夏季B/D, 秋季J/B/F, 其中春季优势类群D(针杆藻属), 夏季B(梅尼小环藻)/D(菱形藻属)演替为秋季主要功能类群J(四尾栅藻)/B(梅尼小环藻)/F(蹄形藻)。SIMPER分析指示了不同季节间群落差异的主要贡献功能类群(表 3), S2/H1/B/D/Lo/X1/MP在3组SIMPER分析中均有稳定贡献, 经分析S2/H1/B/D/Lo/X1/MP对于季节间水体环境变量变化的生态适应性, 可能是导致金河湾湿地季节间功能类群群落差异的主要原因。
| 季节 Seasons |
湿地 Wetlands |
代表性功能类群丰度/(×106个/L) Abundance of representative functional groups |
|||||||||||
| B | D | F | H1 | J | Lo | MP | S1 | S2 | SN | X1 | Y | ||
| 春季 Spring | 湖泊型湿地 | 0.45±0.36 | 0.07±0.18 | 0.00±0.18 | 0.06+0.14 | 0.00±0.00 | 0.30±0.28 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 1.39±0.86 | 0.00±0.00 |
| 河流型湿地 | 0.25±0.02 | 0.13±0.10 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.27±0.00 | 0.00±0.00 | |
| 沼泽湿地 | 0.00 | 0.13 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 | |
| 人工湿地 | 0.00 | 1.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 夏季 Summer | 湖泊型湿地 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.26±0.00 | 0.00±0.00 | 1.70±1.87 | 0.00±0.00 | 0.00±0.00 | 14.07±5.04 | 0.48±0.00 | 0.00±0.00 | 0.00±0.00 |
| 河流型湿地 | 0.39±0.28 | 0.20±0.00 | 0.00±0.00 | 0.15±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.91+0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | |
| 沼泽湿地 | 0 | 0.06 | 0 | 0 | 0 | 0 | 0.15 | 0.08 | 0 | 0 | 0 | 0.08 | |
| 人工湿地 | 0.31 | 0.28 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 秋季 Autumn | 湖泊型湿地 | 0.41±0.50 | 0.00±0.00 | 0.00±0.00 | 3.77±3.22 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.70+0.62 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 |
| 河流型湿地 | 0.58±0.08 | 0.00±0.00 | 0.03±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.00±0.00 | 0.16±0.01 | 0.00±0.00 | |
| 沼泽湿地 | 0.60 | 1.05 | 0.00 | 0.00 | 0.00 | 0.00 | 1.78 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 人工湿地 | 0.32 | 0.00 | 0.00 | 0.22 | 0.38 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 双因素方差分析 Two-way ANOVA (P-value) | |||||||||||||
| 季节间 | P < 0.05 | P>0.05 | P>0.05 | P < 0.05 | P>0.05 | P < 0.05 | P>0.05 | P>0.05 | P < 0.05 | P>0.05 | P < 0.05 | P>0.05 | |
| 不同水体间 | P>0.05 | P < 0.05 | P>0.05 | P>0.05 | P < 0.05 | P>0.05 | P < 0.05 | P < 0.05 | P>0.05 | P>0.05 | P>0.05 | P < 0.05 | |
| 由于采样点数目限制, 沼泽湿地和人工湿地无标准差数据 | |||||||||||||
| 季节 Seasons |
总差异 Global test R |
组间差异 Pairwise test R |
显著水平 P value |
主要贡献功能类群 Main contribute groups |
| 0.384 | ||||
| 春季+夏季 Spring + Aummer | 0.5 | 0.001 | S2/X1/B/D/Lo/H1 | |
| 春季+秋季 Spring + Autumn | 0.335 | 0.001 | X1/H1/B/D/S2/Lo | |
| 夏季+秋季 Summer + Autumn | 0.303 | 0.005 | S2/H1/B/D/Lo/X1/MP |
金河湾湿地春、夏、秋三季4种水体环境变量及双因素方差分析结果见表 4。在本次研究期间, 金河湾湿地WT、pH、SpCond和TP在春、夏、秋季节间变化差异显著(P < 0.05)。另外6项环境变量虽然在统计分析上差异不明显(P>0.05), 但随着季节变化仍呈现一定的时空分布规律(表 4)。在调查期间金河湾湿地夏季水温(29.40℃)明显高于春、秋两季(22.98℃和12.19℃)(P < 0.05)。金河湾湿地水体为碱性, pH为8.78, 变化范围在7.40—9.82之间。总磷(TP)呈现从春季到秋季下降的规律, 平均为0.19 mg/L, 变化范围在0.08—0.36 mg/L之间。叶绿素a(Chl-a)平均浓度为19.82μg/L, 变化范围在5.98—39.77 μg/L之间。溶解氧(DO)、氧化还原电位(ORP)和浊度(Tur)呈现随着春、夏、秋三季的推移逐渐升高的规律, 平均DO、ORP和Tur分别为7.42 mg/L、344.28 U/mV和14.06 NTU, 变化范围分别在5.15—10.51 mg/L、173.00—548.00 U/mV和2.30—38.50 NTU之间。总氮(TN)和化学需氧量(COD)呈现春季最高, 夏季最低的规律, 平均TN和COD分别为0.99 mg/L和21.81 mg/L, 变化范围分别在0.13—2.00 mg/L和7.00—35.00 mg/L之间。
| 季节 Seasons |
湿地 Wetlands |
水温 WT/℃ |
pH | 溶解氧 DO/ (mg/L) |
点多率 SpCond/ (μs/cm) |
氧化还原电位 ORP/ (U/mV) |
浊度 Tur/ (NTU) |
叶绿素 Chl-a/ (μg/L) |
总氮 TN/ (mg/L) |
总磷 TP/ (mg/L) |
化学需氧量 COD/ (mg/L) |
| 春季 Spring | 湖泊型湿地 | 23.01±2.00 | 9.08±0.25 | 6.52±0.71 | 222.86±8.81 | 227.14±23.5. | 7.71±2.36 | 21.84±3.96 | 1.64±0.34 | 0.17±0.08 | 24.00±3.85 |
| 河流型湿地 | 21.87±0.67 | 8.51±0.07 | 6.41±0.10 | 223.33±5.77 | 257.33±11.02 | 19.93±7.51 | 21.32±6.44 | 1.63±0.55 | 0.18±0.06 | 31.33±4.73 | |
| 沼泽湿地 | 26.00 | 7.40 | 6.05 | 460.00 | 295.00 | 3.50 | 6.01 | 1.80 | 0.27 | 33.00 | |
| 人工湿地 | 23.00 | 8.65 | 5.70 | 260.00 | 255.00 | 2.90 | 9.85 | 0.80 | 0.36 | 14.00 | |
| 夏季 Summer | 湖泊型湿地 | 29.26±0.78 | 9.55±0.23 | 7.80±0.29 | 180.96±7.09 | 366.00±6.63 | 10.51±3.14 | 13.48±1.68 | 0.20±0.09 | 0.13±0.04 | 17.29±4.23 |
| 河流型湿地 | 29.90±0.44 | 8.93±0.19 | 7.52±0.27 | 203.00±1.73 | 379.00±6.56 | 18.80±2.88 | 23.17±4.98 | 0.15±0.00 | 0.27±0.15 | 11.33±3.51 | |
| 沼泽湿地 | 28.40 | 7.56 | 6.99 | 334.00 | 389.00 | 19.80 | 7.80 | 0.80 | 0.27 | 28.00 | |
| 人工湿地 | 29.90 | 8.37 | 7.66 | 211.00 | 387.00 | 2.30 | 5.98 | 0.18 | 0.26 | 7.00 | |
| 秋季 Autumn | 湖泊型湿地 | 12.30±0.92 | 8.62±0.361 | 8.57±0.91 | 164.29±9.76 | 424.71±126.72 | 20.36±10.02 | 28.92±6.60 | 1.10±0.21 | 0.18±0.03 | 26.29±4.54 |
| 河流型湿地 | 12.70±2.13 | 8.19±0.05 | 8.27±0.53 | 166.67±5.77 | 413.33±72.76 | 17.20±6.23 | 18.64±9.40 | 1.23±0.06 | 0.19±0.11 | 16.00±4.58 | |
| 沼泽湿地 | 9.50 | 7.69 | 5.97 | 360.00 | 436.00 | 33.60 | 13.88 | 1.40 | 0.26 | 33.00 | |
| 人工湿地 | 12.60 | 8.73 | 7.94 | 130.00 | 358.00 | 6.20 | 29.91 | 1.00 | 0.14 | 21.00 | |
| 双因素方差分析 Two-way ANOVA (P-value) | |||||||||||
| 季节间 | P < 0.05 | P < 0.05 | P>0.05 | P < 0.05 | P>0.05 | P>0.05 | P>0.05 | P>0.05 | P < 0.05 | P>0.05 | |
| 不同水体间 | P>0.05 | P < 0.05 | P>0.05 | P < 0.05 | P>0.05 | P < 0.05 | P>0.05 | P>0.05 | P < 0.05 | P < 0.05 | |
| 沼泽湿地和人工湿地, 由于采样点数目限制, 无标准差数据 | |||||||||||
对金河湾湿地WT、pH、SpCond、DO、ORP、Tur、Chl-a、TP、TN和COD共10种环境变量与代表性功能类群进行排序分析, 寻找金河湾湿地功能类群在季节间和空间上的分布规律, 并探索驱动功能类群演替的主要环境变量。将代表性功能类群进行去趋势分析(DCA), DCA结果显示排序轴最长梯度长度值为6.466, 因此属于单峰模型的典范对应分析(CCA), 可更准确的分析金河湾湿地浮游植物功能类群的时空分布规律。结果显示, CCA排序轴1和轴2的特征值分别为0.738和0.512, 功能类群与环境变量相关系数达到0.955和0.878, 前四个排序轴共解释88.5%的累计种类环境变量百分比。说明本次CCA排序结果可靠, 基本反映了研究期间金河湾湿地代表性功能类群与环境变量之间的生态关系。经蒙特卡洛检验, TN、pH和SpCond与浮游植物功能类群时空动态变化关系密切(图 6)。

|
| 图 6 代表性浮游植物功能类群与环境变量的CCA分析 Fig. 6 CCA of representative functional groups and environmental variables in Jin Hewan wetland |
浮游植物功能类群演替主要是由水体营养盐、稳定性及浮游植物的形态、生理适应特征、悬浮机制、营养盐吸收利用效率、光能捕获能力以及固碳速率等相互作用的结果[25]。这些因素共同决定着浮游植物对其特定生长环境的选择机制, 成为浮游植物在水环境中生存或者消失的限制因素[24-25]。
金河湾湖泊型湿地浮游植物功能类群特征为, 由春季的B(梅尼小环藻)/X1(狭形纤维藻)演替为夏季S2(螺旋藻)/Lo(平裂藻)/SN(中华小尖头藻), 最后被秋季的H1(水华束丝藻)/B(小环藻属)所取代。金河湾湿地春季营养盐浓度相对较高, 光照强度和水体温度相对较低(23℃), 功能类群B的代表种类为小环藻属, 其生境类型为浑浊较浅的富营养类型水体, 耐受条件为弱光和低温。与此同时, B类群小环藻属的形态有利于维持高水平的s/v比值, 同时具有高效的捕光能力, 因此在春季较低光照条件中获得良好的竞争优势。S2类群的螺旋藻为夏季绝对优势的种类, 此类群种类适宜生长在温暖、浅层且呈碱性的水体中, 对河流冲刷较为敏感。夏季平均水温29.3℃, 平均pH值9.1, 且湖泊型湿地受雨水河流波动影响较小, 为此种藻类的生长提供了适应条件。夏季浮游植物生长时吸收了水体中大量的CO2, 对pH升高起到促进作用, 春季占据优势的B类群对水体pH升高敏感, 这可能是B类群在夏季环境适应中处于相对劣势的原因之一, 而S2类群(螺旋藻)成为了夏季优势类群。H1类群(固氮鱼腥藻、水华束丝藻)通常适宜生存在营养水平较高的水体中, 同时可耐受较低氮含量的条件[9]。一些研究指出, 低N、P营养环境中, 水华束丝藻(H1类群)可通过调节自身的酶系统来提高对营养的吸收和利用, 从而满足自身生长和细胞增殖的需要[26]。金河湾湖泊型湿地水体总氮含量从春季到秋季存在明显的先下降后回升过程(P < 0.05), 夏季较低的营养含量下, H1类群虽然相对丰度较低, 由于其特殊的生态适应性, 仍在此条件下存活并占据一定比例。到了秋季, 营养盐水平升高, H1类群(水华水丝藻)重新占据优势地位, 成为秋季的绝对优势类群。相应的B类群(小环藻属)随着秋季水体温度及pH降低也重新回到优势地位。
从河流型湿地浮游植物功能类群分析, 春季以X1/B/D为代表性功能类群, 其优势种类为狭形纤维藻、梅尼小环藻及菱形藻属。夏季演替为S2/B/H1的优势种类螺旋藻、水华束丝藻, S2/H1类群浮游植物均为蓝藻门植物, 喜较高温度、低氮的水体环境, 夏季高水温(28.4 ℃)及TN含量的降低(0.15 mg/L)使得蓝藻门浮游植物(固氮鱼腥藻、水华束丝藻)在群落中占据生长优势。秋季水温降低, 水体营养盐浓度升高, B(小环藻属)/X1(狭形纤维藻)代替了夏季大量生长的蓝藻门种类, 其中B(小环藻属)为3个季节的优势类群藻种, 小环藻属的种类(Cyclotella sp.)具有高效的捕光能力, 因此可以进行良好的光合作用供自身生长繁殖。
从沼泽湿地来看, 其浮游植物功能类群变化特征为春季D/MP/Y, 夏季MP/Y/S1/D, 秋季MP/D/B, 湿地中D(菱形藻属)/MP(舟形藻属、曲壳藻属)在该水体中一直处于优势地位。沼泽湿地大量杂草丛生, 在水体低光照条件下, 耐受低光条件的功能类群长势良好, 所以D/MP处于3个季度沼泽湿地功能类群中优势地位。Y类群(啮蚀隐藻)能够利用鞭毛在水体中浮动, 适应于较高水温、低光照的夏季沼泽水体[7], 因此, 啮蚀隐藻在夏季的该类型的湿地中占据较高优势。同时此类型水体具有相对较高的营养盐条件, D/MP/Y一般生存在营养盐较高的小型水体中[25], 而此生境中低光条件在一定程度上限制了其他种类的生长, 因此D/MP/Y在环境适应和生存空间竞争中处于绝对的优势, 所以该类型湿地藻种丰富度相对较低, 优势种突出。
从人工湿地来看, 其浮游植物季节演替为春季D, 夏季B/D, 秋季J/B/F, 主要优势藻种为D(针杆藻属、菱形藻属)/B(梅尼小环藻)。B/D类群的藻类可耐受人为活动干扰所带来的光缺乏以及水体冲刷, 而F类群又可耐受相对较低的营养水平[25]。该湿地为人工打造的圆形水池, 水体面积较小, 水体受人为活动扰动明显, 水体营养盐水平为4种水体最低。因此, 其功能类群季节演替特征不明显, 浮游植物种类丰富度相比湖泊型湿地和河流型湿地明显降低。
在本次研究中, 所选取的水体类型多样, 水体环境变量时空变化较为明显, 浮游植物种类丰富, 而功能类群的演替过程, 反映了金河湾湿地浮游植物群落结构对季节间环境变化的响应。对于决定群落结构演替的功能类群如S2/H1/B/D/Lo/X1/MP, 可视为金河湾湿地季节间水体环境变化的指示类群。
3.2 浮游植物功能类群演替的影响因子浮游植物的生长和代谢受多种环境变量的共同作用, 如光照、营养盐含量(氮、磷)等, 在不同的环境变量驱动下, 可使浮游植物功能类群发生规律性的演替[27-28]。在CCA排序图中, 采样点沿着排序轴的分布情况, 体现了该样点中所有物种的贡献率及对环境梯度的响应过程[27]。本研究中, 金河湾湿地不同季节和生境的采样点沿着CCA排序轴分布特征明显, 基本反映了浮游植物功能类群在演替过程中对于主要环境变量分布模式所做出的响应。
TN是评估湿地水体富营养水平的重要指标, 作为浮游植物生长代谢所需的大量元素之一, 当水体中TN含量过高或过低时, 往往会成为影响浮游植物群落结构的演替的限制因子[29-30]。调查期间, 金河湾湿地TN含量在0.12—2.00 mg/L之间变化, 最小值出现在夏季的河流型湿地。从整体来看, 夏季总氮含量最低, 该季节降雨量充沛, 而金河湾湿地整体水域环境较为封闭, 不受来自其余河流冲刷的影响, 导致氮磷含量降低, 而此时该湿地水体中大量生长的为H1/S2/Lo蓝藻类群(固氮鱼腥藻、水华鱼腥藻及平裂藻等), 这种现象在其他的一些研究中得到相同结论[31]。蓝藻之所以能够爆发性生长代替其他藻种, 一方面与其自身生理特征有关, 如固氮、伪空泡、光吸收及营养盐利用的能力, 另一方面则与系统内物理、化学和生物环境有关。蓝藻因其自身的固氮作用, 能在氮含量较低水体中大幅度生长[32]。此外, 还与蓝藻本身的某些生理特性有关, 如气囊及抗拒紫外线伤害的能力, 从而避免了夏季高光照射条件, 进而在氮含量较低的夏季水环境中处于优势地位[25]。类群X1/J/F/B体现为, 随着TN含量的升高丰度成上升的趋势, 并且成为春、秋两季河流和湖泊湿地中的优势类群(图 6)。推测在春、秋两季TN含量相对较高的水体环境中, 如X1类群作为“先锋种”可在资源竞争中占据优势[7, 9], 较高浓度的氮营养盐也可以维持纤维藻的藻细胞较长时间的增长[33]。因此, 在不同浓度TN环境中, 会衍生不同的功能类群分布模式。另外, 水体中氮磷比的含量会影响浮游植物种群适应及群落组成[25]。通常情况下, 浮游植物对营养盐的吸收利用大致遵循N/P比为16:1的Redfield定律, 当氮磷比大于16:1时, 表现为磷限制, 而N/P小于16:1时, 为氮限制[34]。春、夏、秋三季间, 金河湾湿地平均氮磷比为5.96, 属于氮限制。因此, 通过对水体中TN含量的调控, 可对金河湾湿地浮游植物群落演替起到一定的限制作用。
SpCond是与水体中浮游植物生长密切相关的环境变量, 也是反映水体富营养化程度的重要环境指标之一, 其离子种类的组成和水域地质及土壤特征密切相关[35]。通常水体纯净度越高, 则SpCond越低[36]。研究期间, 金河湾湿地SpCond均值为208 μs/cm, 在四种不同类型的湿地中SpCond差异显著(P < 0.05)。CCA排序图显示, D/MP/Y/S1分布与SpCond含量变化密切, 并呈现随着SpCond升高, 丰度上升的趋势, 而这些功能类群主要在沼泽湿地水体中体现较高的权重(图 6b)。沼泽湿地水体较浅, 丛生大量杂草水环境营养物质充足, 不受河流冲击和降雨量的影响, 所以沼泽湿地平均SpCond最高为385 μs/cm。水体中SpCond的高低, 在一定程度上也会反映营养盐浓度的水平[35]。沼泽湿地水体较高的SpCond伴随着相对较高的营养盐含量, D组对营养盐缺乏敏感, 较高的电导率条件, 在一定程度上避免了该类群的生存环境压力。同时沼泽湿地水体光照条件相对较低, 因此可耐受于低光条件的S1/Y类群在藻类群落中占据优势[25]。经分析, 由SpCond变化所反映的水体营养盐和纯净度的改变, 影响了沼泽湿地中D/MP/Y/S1类群的生长、演替和分布。这与在哈尔滨同纬度的另一个城市湿地(呼兰河湿地)的研究得出的结论一致[37]。pH与浮游植物群落生态分布密切相关, 尤其是一些硅藻门种类[38]。CCA排序表明, MP/B类群与pH呈现负相关, 类群MP(舟形藻属)为喜弱碱藻种, 类群B对pH升高比较敏感[25], 金河湾湿地整体pH值为7.40—9.82之间, 均值8.78, 呈碱性状态, 因此MP/B类群在环境适应性中处于劣势。H1/J类群在排序轴中呈现与pH正相关的趋势, 但多位于坐标轴原点附近, 经分析, 适度的pH可以促进部分类群藻类的生长, 但湿地水体酸碱度过高时, 有可能会限制大部分藻类生长[39]。综合来看, TN是驱动金河湾湿地浮游植物功能类群生长选择机制的主要环境变量, 而水体中SpCond浓度和pH值与功能类群演替过程密切相关。
水华束丝藻是引起我国淡水水华常见种之一, 在水华过程中可产生碱类神经毒素[26]。在本研究中, 包含水华束丝藻和固氮鱼腥藻的H1类群在秋季成为金湾湿地湖泊型水体中优势类群。以往的研究也认为水华束丝藻易出现在营养水平较高的水体中[7, 26], 虽然与杭州西湖1981年发生的束丝藻水华相比, 本次研究中水化束丝藻丰度较低, 但当TN浓度>0.2 mg/L、TP浓度>0.02 mg/L时水体易出现富营养化, 综合来看, 金河湾湿地水体未来可能存在富营养化的风险。城市湿地易受到人为活动干扰, 导致湿地水体环境变化多样、生态结构较为复杂[13]。在浮游植物功能类群的划分过程中, 每个类群不仅体现了浮游植物生态适应特点, 还结合了相应的环境特征[9]。虽然以往对于浮游植物功能类群的生态研究多集中在湖泊和河流生态系统中[11, 24], 但在本研究中, 金河湾湿地浮游植物功能类群的时空分布特征与水体某些环境变量(如SpCond、pH和TN)变化特点相匹配, 因此通过分析城市湿地浮游植物功能类群季节间的演替, 基本可以有效地反映浮游植物在湿地生态系统中的生态位及对水环境变化做出的响应, 并对未来环境的变化做出一定的预测。
| [1] | Cardoso S J, Roland F, Loverde-Oliveira S M, de Moraes Huszar V L. Phytoplankton abundance, biomass and diversity within and between Pantanal wetland habitats. Limnologica-Ecology and Management of Inland Waters, 2012, 42(3): 235–241. DOI:10.1016/j.limno.2012.01.002 |
| [2] | Mitsch W J, Gosselink J G. Wetlands.4th ed. Hoboken, NJ, USA: John Wiley & Sons, 2007. |
| [3] | Sebastiá M T, Rodilla M, Sanchis J A, Altur V, Gadea I, Falco S. Influence of nutrient inputs from a wetland dominated by agriculture on the phytoplankton community in a shallow harbour at the Spanish Mediterranean coast. Agriculture, Ecosystems & Environment, 2012, 152: 10–20. |
| [4] | Fang L, Wong P K, Lin L I, Lan C, Qiu J W. Impact of invasive apple snails in Hong Kong on wetland macrophytes, nutrients, phytoplankton and filamentous algae. Freshwater Biology, 2010, 55(6): 1191–1204. DOI:10.1111/j.1365-2427.2009.02343.x |
| [5] | Wondmagegne T, Wondie A, Mingist M, Vijverberg J. Seasonality in abundance, biomass and production of the phytoplankton of Welala and Shesher wetlands, Lake Tana Sub-Basin (Ethiopia). Journal of Water Resource & Protection, 2012, 4(10): 877–884. |
| [6] | Roy A, Sarkar D, Dutta S, Roy R, Mondal S, Roy M, Mitra A. Phytoplankton community of East Kolkata Wetlands as an indicator of the water quality. International Journal of Advanced Research, 2017, 5(4): 104–110. |
| [7] | Reynolds C S, Huszar V, Kruk C, Naselli-Flores L, Melo S. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 2002, 24(5): 417–428. DOI:10.1093/plankt/24.5.417 |
| [8] | Padisák J, Crossetti L O, Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification:a critical review with updates. Hydrobiologia, 2009, 621(1): 1–19. DOI:10.1007/s10750-008-9645-0 |
| [9] | 胡韧, 蓝于倩, 肖利娟, 韩博平. 淡水浮游植物功能群的概念、划分方法和应用. 湖泊科学, 2015, 27(1): 11–23. |
| [10] | Kruk C, Mazzeo N, Lacerot G, Reynolds C S. Classification schemes for phytoplankton:a local validation of a functional approach to the analysis of species temporal replacement. Journal of Plankton Research, 2002, 24(9): 901–912. DOI:10.1093/plankt/24.9.901 |
| [11] | Padisák J, Borics G, Fehér G, Grigorszky I, Oldal I, Schmidt A, Zámbóné-Doma Z. Dominant species, functional assemblages and frequency of equilibrium phases in late summer phytoplankton assemblages in Hungarian small shallow lakes. Hydrobiologia, 2003, 502(1/3): 157–168. |
| [12] | Sevindik T O, Çelik K, Naselli-Flores L. Spatial heterogeneity and seasonal succession of phytoplankton functional groups along the vertical gradient in a mesotrophic reservoir. Annales de Limnologie-International Journal of Limnology, 2017, 53: 129–141. DOI:10.1051/limn/2016040 |
| [13] | 封晓梅. 《湿地公约》与我国的湿地保护[D]. 青岛: 中国海洋大学, 2008. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1336998 |
| [14] | 殷康前, 倪晋仁. 湿地研究综述. 生态学报, 1998, 18(5): 539–546. DOI:10.3321/j.issn:1000-0933.1998.05.014 |
| [15] | 周俊. 苏州湿地公园藻类群落结构的研究[D]. 上海: 上海师范大学, 2012. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2115884 |
| [16] | 陆强, 陈慧丽, 邵晓阳, 王莹莹, 陶敏, 何京, 唐龙. 杭州西溪湿地大型底栖动物群落特征及与环境因子的关系. 生态学报, 2013, 33(9): 2803–2815. |
| [17] | 周芳菲. 基于AHP的哈尔滨金河湾湿地公园景观体验优化策略研究[D]. 哈尔滨: 哈尔滨工业大学, 2014. |
| [18] | 马婷慧. 哈尔滨金河湾湿地公园POE调查及优化策略研究[D]. 哈尔滨: 哈尔滨工业大学, 2013. |
| [19] | 林聪, 李志国, 梁庆宇, 王影, 范亚文. 黑龙江省金河湾湿地藻类植物的初步研究. 哈尔滨师范大学:自然科学学报, 2015, 31(4): 92–96. |
| [20] | 国家环境保护总局. 水和废水监测分析方法.第四版. 北京: 中国环境科学出版社, 2002. |
| [21] | 胡鸿钧, 魏印心. 中国淡水藻类-系统/分类及生态. 北京: 科学出版社, 2006. |
| [22] | Bao W M, Wang Q X, Reimer C W. Diatoms from the Changbaishan Mountain area. Bulletin of Botanical Research, 1992, 12(2): 125–143. |
| [23] | Rumrich U., Lange-Bertalot H., Rumrich M. Diatoms of the Andes from Venezuela to Patagonia/Tierradel Fuego. Icon. Diat, 2000, 9: 1–673. |
| [24] | Xiao L J, Wang T, Hu R, Han B P, Wang S, Qian X, Padisák J. Succession of phytoplankton functional groups regulated by monsoonal hydrology in a large canyon-shaped reservoir. Water Research, 2011, 45(16): 5099–5109. DOI:10.1016/j.watres.2011.07.012 |
| [25] | Reynolds C S. Ecology of Phytoplankton. Cambridge: Cambridge University Press, 2006. |
| [26] | 刘永梅, 刘永定, 李敦海, 沈银武. 氮磷对水华束丝藻生长及生理特性的影响. 水生生物学报, 2007, 31(6): 774–779. DOI:10.3321/j.issn:1000-3207.2007.06.002 |
| [27] | Droop M R. Some thoughts on nutrient limitation in algae. Journal of Phycology, 1973, 9(3): 264–272. |
| [28] | Rochelle-Newall E J, Chu V T, Pringault O, Amouroux D, Arfi R, Bettarel Y, Bouvier T, Bouvier C, Got P, Nguyen T M H, Mari X, Navarro P, Duong T N, Cao T T T, Pham T T, Ouillon S, Torréton J P. Phytoplankton distribution and productivity in a highly turbid, tropical coastal system (Bach Dang Estuary, Vietnam). Marine Pollution Bulletin, 2011, 62(11): 2317–2329. DOI:10.1016/j.marpolbul.2011.08.044 |
| [29] | Pavlou S P, Friederich G E, Macisaac J J. Quantitative determination of total organic nitrogen and isotope enrichment in marine phytoplankton. Analytical Biochemistry, 1974, 61(1): 16–24. DOI:10.1016/0003-2697(74)90327-3 |
| [30] | Paparazzo F E, Williams G N, Pisoni J P, Solís M, Esteves J L, Varela D E. Linking phytoplankton nitrogen uptake, macronutrients and chlorophyll- a in SW Atlantic waters:The case of the Gulf of San Jorge, Argentina. Journal of Marine Systems, 2017, 172: 43–50. DOI:10.1016/j.jmarsys.2017.02.007 |
| [31] | 秦伯强, 杨桂军, 马健荣, 邓建明, 李未, 吴挺峰, 刘丽贞, 高光, 朱广伟, 张运林. 太湖蓝藻水华"暴发"的动态特征及其机制. 科学通报, 2016, 61(7): 759–770. |
| [32] | Varnoosfaderani A M, Khoii J A, Rafiee G R. Investigating the effect of Biodrof systems based on algae-bacterial biofilm for removing total Nitrogen, Phosphorus from domestic wastewater. Journal of Aquatic Ecology, 2015, 4(4): 18–18. |
| [33] | 王英英. 不同氮磷质量浓度对太浦河四种优势藻类生长影响的研究[D]. 上海: 上海师范大学, 2016. |
| [34] | Redfield A C. The biological control of chemical factors in the environment. Science Progress, 1960, 11: 150–170. |
| [35] | Flores L N, Barone R. Phytoplankton dynamics in two reservoirs with different trophic state (Lake Rosamarina and Lake Arancio, Sicily, Italy). Hydrobiologia, 1998, 369-370: 163–178. DOI:10.1023/A:1017063708180 |
| [36] | Trebitz A S, Nestlerode J A, Herlihy A T. USA-scale patterns in wetland water quality as determined from the 2011 National Wetland Condition Assessment. Environmental Monitoring & Assessment, 2017. |
| [37] | 陆欣鑫, 刘妍, 范亚文. 呼兰河湿地夏、秋两季浮游植物功能分组演替及其驱动因子. 生态学报, 2014, 34(5): 1264–1273. |
| [38] | Jin M, Champagne P, Hall G. Effects of different substrates in the mitigation of algae-induced high pH wastewaters in a pilot-scale free water surface wetland system. Water Science & Technology, 2017, 75(1): 1–10. |
| [39] | Ter Braak C J F, Van Dame H. Inferring pH from diatoms:a comparison of old and new calibration methods. Hydrobiologia, 1989, 178(3): 209–223. DOI:10.1007/BF00006028 |