 2018, Vol. 38
2018, Vol. 38文章信息
- 王晓娟, 孔繁花, 尹海伟, 徐海龙, 李俊生, 蒲英霞.
- WANG Xiaojuan, KONG Fanhua, YIN Haiwei, XU Hailong, LI Junsheng, PU Yingxia.
- 高温天气植被蒸腾与遮荫降温效应的变化特征
- Characteristics of vegetation shading and transpiration cooling effects during hot summer
- 生态学报. 2018, 38(12): 4234-4244
- Acta Ecologica Sinica. 2018, 38(12): 4234-4244
- http://dx.doi.org/10.5846/stxb201712032173
-
文章历史
- 收稿日期: 2017-12-03
- 修订日期: 2018-04-10





2. 南京大学建筑与城市规划学院, 南京 210093;
3. 南京大学地理与海洋科学学院, 南京 210023
2. School of Architecture and Urban Planning, Nanjing University, Nanjing 210093, China;
3. Department of Geographical Information Science, Nanjing University, Nanjing 210023, China
城市绿地通过植被冠层对太阳辐射的阻挡(遮荫)与叶片的蒸腾作用能有效降低植被冠层表面和林下的大气温度, 产生明显降温效应[1], 从而有效缓解城市热岛强度、减少降温能耗[2]。因而, 开展对城市中不同树种植被遮荫与蒸腾降温效应的量化研究是科学优化植被温度调控服务的重要基础。
目前植被遮荫降温效应的测算方法主要有3种:测量冠层上下的太阳辐射差[3]、测量冠层上下的空气温度差[4-5]和采用专业软件进行模拟[6]。例如, Tsiros[4]采用测量冠层上下空气温度差的方法研究了雅典5个街区的树木遮荫降温效应, 发现14:00时遮荫降温约为0.5—1.6℃。与遮荫降温的测度方法相比, 目前植被蒸腾作用量化方法的种类不多, 使用最普遍的是树干液流(Sap Flow)测定法[7], 主要包括热脉冲[8-10]、热平衡[11-12]、热扩散[13]等热技术探测方法[14]。然而, 通过热技术仅能测定单株尺度的植物蒸腾耗水量, 因而许多学者利用胸径(Diameter at breast height, DBH)[10, 12, 15]、边材面积[16-17]、叶面积[8, 18]等作为扩展指标, 实现了由单株植被蒸腾耗水量到林分尺度上冠层蒸腾量的尺度扩展[10, 19-20]。例如, Bosch等[10]采用热脉冲法观测了湿地松(Pinus elliottii)、长叶松(Pinus palustris Mill)、北美鹅掌楸(Liriodendron tulipifera)的树干液流, 利用胸径作为扩展指标对树干液流进行尺度拓展。目前国内外针对植被蒸腾或遮荫单项降温效应的研究较多, 但同时进行植被遮荫与蒸腾降温效应综合量化, 并进而挖掘遮荫与蒸腾对植被总降温效应贡献率变化特征的研究尚不多见[3]。
本文以南京市栖霞区某小型绿地单元为研究区, 采用热扩散探针技术对高温晴朗天气下香樟(Cinnamomum camphora)、雪松(Cedrus deodara)、杨树(Populus tomentosa Carr)3个树种典型植株树干液流进行了观测, 并利用叶面积指数(Leaf Area Index, LAI)对单株植被蒸腾量进行尺度拓展, 估算了3个树种林分尺度上的蒸腾量与蒸腾降温效应, 并根据林上、林下太阳辐射值计算了不同树种与整个绿地单元的遮荫降温效应, 进而阐明了蒸腾与遮荫降温对总降温效应贡献率的变化特征。研究结果将有助于更好地理解不同树种蒸腾与遮荫降温效应的变化特征与规律, 为科学优化不同树种植被的温度调控服务提供参考和依据。
1 研究区概况本文以位于南京市栖霞区万达茂以东、仙林大道以北(32°7′N, 118°59′E)的一个小型绿地单元作为研究区(图 1)。该小型绿地呈三角形, 面积约为1.97 hm2; 最初为苗圃用地, 主要有香樟、雪松、杨树3种类型的植被, 面积分别为0.52、0.79、0.66 hm2, 林相整体长势较为一致, 树龄均为7a。该研究区属于亚热带季风气候, 四季分明, 多年平均温度15.4℃, 多年平均降水量1106 mm。

|
| 图 1 研究区、树干液流观测样树及微气象观测站分布图 Fig. 1 Distribution map of study area, sample trees used for sap flow measurement, and micro-meteorological observation stations |
使用美国Dynamax标准茎流观测系统对香樟、雪松和杨树3个树种的1—10号样树进行树干液流的观测(表 1, 图 1)。加热探针(Thermal Dissipation Probe, TDP)安装在离地面1.3 m处, 数据采样间隔为1 min, 数据存储间隔为60 min。由于TDP探针数量与线缆长度的限制, 每次只能同时观测2个树种。因而, 本文选取2015年8月3—5日的香樟、雪松、2016年8月13—15日的雪松、杨树树干液流的观测数据来进行高温天气下植被蒸腾量与蒸腾降温效应的估算。树干液流参照Granier[21]的研究进行计算(式1—4)。
| 样树编号 Tree number |
树种 Tree species |
胸径 Diameter at breast height/mm |
边材面积 Sapwood area/mm2 |
叶面积指数 Leaf area index |
树高 Height/m |
冠层高度 Canopy height/m |
冠幅 Canopy width/m |
观测时间 Observation time |
| 1# | 香樟 | 11.15 | 75.86 | 3.347 | 6.03 | 4.32 | 3.37 | 2015年8月 |
| 2# | 香樟 | 7.19 | 35.46 | 3.065 | 5.16 | 4.12 | 1.90 | 2015年8月 |
| 3# | 雪松 | 12.77 | 108.54 | 2.496 | 4.76 | 4.05 | 2.85 | 2015年8月 |
| 4# | 雪松 | 11.45 | 93.85 | 2.407 | 3.72 | 3.07 | 2.63 | 2015年8月 |
| 5# | 雪松 | 8.02 | 42.08 | 2.998 | 4.16 | 3.61 | 2.11 | 2016年8月 |
| 6# | 雪松 | 11.91 | 91.98 | 4.557 | 5.11 | 4.65 | 3.51 | 2016年8月 |
| 7# | 雪松 | 14.00 | 127.32 | 3.548 | 7.33 | 6.84 | 3.54 | 2016年8月 |
| 8# | 雪松 | 16.83 | 183.33 | 2.931 | 5.60 | 5.31 | 3.88 | 2016年8月 |
| 9# | 杨树 | 8.76 | 49.74 | 2.110 | 6.63 | 3.43 | 1.45 | 2016年8月 |
| 10# | 杨树 | 19.62 | 249.51 | 2.867 | 11.09 | 8.22 | 3.14 | 2016年8月 |
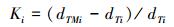
|
(1) |
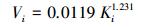
|
(2) |
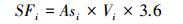
|
(3) |
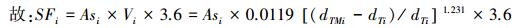
|
(4) |
式中, Ki为参数, 根据树种i探针观测温度获取; dTMi为树种i观测样树的上下探针之间的最大昼夜温差(℃); dTi为样树i瞬时温差(℃); Vi为树干液流速率(cm/s); Asi为单株样树的边材横截面积(cm2); SFi则为树种i单株样树的树干液流(kg/h)。
2.1.2 树种参数提取使用RIEGL VZ-400三维激光扫描成像系统获取了研究区香樟、雪松和杨树3个树种单株样树(图 2)和整片林地单元的三维点云数据, 计算提取了1—10号样树的树高、冠层高度、冠幅等植被特征参数(表 1), 并基于冠层孔隙度Beer-Lambert定律[22-23]计算了样树的叶面积指数(表 1)。本文叶面积指数是指单位地面面积上植物叶片总面积占地面面积的倍数, 使用三维点云数据计算而得, 其具体计算过程参见公式(5)和(6)。

|
| 图 2 3个树种的激光雷达点云数据 Fig. 2 LiDAR point cloud data of three tree species |
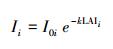
|
(5) |
故:
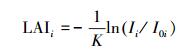
|
(6) |
式中, 对于树种i, LAIi表示叶面积指数, Ii是冠层下面的光, I0i是冠层上面的光, Ii/I0i为冠层孔隙度, 可根据点云数据计算获取。k是消光系数, 可通过叶倾角和天顶角获取, 具体如下式(7)—(8):
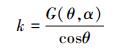
|
(7) |
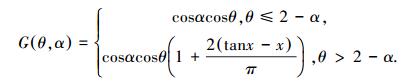
|
(8) |
式中, G(θ, α)表示叶面区域在入射光线的垂直方向的平均投影, x=1/cotαcotθ, α为叶面元素的倾角, θ为太阳入射光线的天顶角。
2.1.3 气象观测使用Hobo U30和CR1000小型气象观测站进行太阳辐射(W/m2)、风速(m/s)、空气温度(℃)、空气湿度(%)、降雨量(mm)等微气象环境因子的观测(表 2), 数据采样间隔为60 s, 数据存储间隔为1 min。由于2015年8月3—5日和2016年8月13—15日两个时段的气象特征比较相似, 均属于夏季高温晴朗天气(最低气温分别为25.7、25.7℃, 最高气温分别为39.7、36.9℃; 白天平均太阳总辐射分别为552.4、493.3 W/m2), 所以本文选取这两个时段来进行3个树种植被蒸腾与遮荫降温效应的对比分析。
| 传感器名称Smart sensor | 型号Product model | 测试参数Parameter | 精度Accuracy |
| 温湿度传感器 Temperature/Humidity sensor |
HMP155A S-THB-M002 |
温度、相对湿度 温度、相对湿度 |
±0.1℃、±1% ±0.2℃(25℃时)、±2.5% |
| 短波辐射传感器 Solar radiation sensor |
S-LIB-M003 | 太阳总辐射 | ±3% |
| 风速传感器 Wind speed sensor |
RM Young L35 S-WCA-M003 |
风速 风速、风向 |
±1% ±0.5m/s、±4° |
| 雨量筒Rainfall sensor | TE525MM | 降水量 | ±1.0% |
首先, 采用Soegaard等、白岩等[18, 20] “单位叶面积上的平均液流速率(SF)×叶面积指数(LAI)”的扩展方法, 将单株样树冠层蒸腾量扩展到林分尺度上的冠层蒸腾量(具体扩展方法与计算过程参见公式(9)和(10):
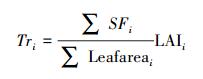
|
(9) |
将公式(3)代入(9), 得到下式(10):
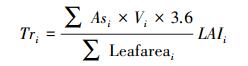
|
(10) |
借鉴已有相关研究[17, 24-25]计算得出3个树种单位地面面积内冠层蒸腾作用所消耗的能量(即蒸腾耗能量, 公式(11))。
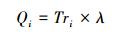
|
(11) |
参照杨士弘和Nakazato等[19, 26]蒸腾降温估算方法(公式(12)), 进行了3个树种蒸腾降温效应的计算。考虑到空气的湍流、对流和辐射作用, 以及空气与叶面之间、空气微气团之间不断地进行热量扩散和交换, 故本研究取底面积为10 m2、厚度为100 m的1000 m3的空气柱作为计算单元。100 m代表了研究区的城市冠层高度, 而10 m2可代表小气候的水平尺度[27]。
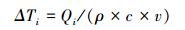
|
(12) |
式中, Tri为树种i林分尺度的冠层蒸腾量(mm/h), Leafareai为树种i单株样树的总叶面积(m2), 可基于三维点云数据计算获取, Qi为树种i冠层蒸腾作用使其单位地面面积内空气损失的热量(kJ h-1 m-2), λ为汽化潜热(kJ/kg), 可由空气温度计算得出(λ=2498.8-2.33Ta, Ta为空气温度), ρ为空气密度, 约为1.29 kg/m3; c为空气比热容, 约为1.03 kJ kg-1 ℃-1; v为空气柱体体积, 本文为1000 m3。
2.3 林分尺度上遮荫降温效应计算将树冠遮荫作用所降低的能量定义为林上、林下太阳总辐射的差值[3], 采用公式(13)和(14)计算得到3个树种单位地面面积内冠层遮荫作用所降低的能量, 并参照杨士弘和Nakazato等[19, 26]蒸腾降温估算方法(公式(15)), 对3个树种的遮荫降温效应进行了计算。

|
(13) |
由于
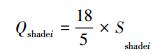
|
(14) |

|
(15) |
式中, Sshadei表示树种i因遮荫作用所降低的能量(W/m2), PARsi表示林上太阳总辐射值(W/m2), PARBi表示林下太阳总辐射值(W/m2), Qshadei为树冠遮荫所降低的能量(kJ/h)。
2.4 蒸腾与遮荫对总降温效应的贡献率计算根据植被蒸腾降温和遮荫降温的计算结果, 分别计算了3个树种蒸腾与遮荫降温对总降温效应的贡献率。为了将不同树种单位空气柱(1000 m3)的蒸腾与遮荫降温效应扩展到小型绿地单元尺度上, 本研究以不同树种面积所占该绿地单元总面积的比重作为权重, 采用加权求和法计算获取了该小型绿地单元的整体蒸腾与遮荫降温效应及其对总降温效应的贡献率。
3 结果与分析 3.1 不同树种树干液流的变化特征由图 3可见, 3个树种单株样树的树干液流均表现出昼高夜低的总体变化趋势; 树干液流在6:00左右开始启动, 12:00左右达到峰值, 且随后维持在高位(即存在一个较宽的平顶峰), 称之为树干液流的“午休”, 这主要是由于正午温度较高, 植物为了保存体内水分, 短暂关闭或减小气孔开合程度, 从而使蒸腾作用在达到峰值后不再增长; 18:00之后树干液流呈急剧下降趋势, 21:00之后变得非常微弱; 在夜间树干液流虽然很小但仍然存在, 这主要是由于根压的作用, 水分被吸收进入植物体内, 补充白天所消耗的水分。与此同时, 在同一个树种内, 随着胸径增大, 树干液流亦显著增大(表 3), 这表明胸径大小是影响树干液流的重要因素之一。

|
| 图 3 观测样树树干液流昼夜变化特征 Fig. 3 The variable characteristics of stem sap flow at each observed sample tree |
| 样树号 Tree number |
树种 Tree species |
边材面积 Sapwood area/cm2 |
日均液流量 Daily average stem sap flow volume /(kg/d) |
样树号 Tree number |
树种 Tree species |
边材面积 Sapwood area/cm2 |
日均液流量 Daily average stem sap flow volume/(kg/d) |
| 1# | 香樟 | 75.86 | 23.82 | 6# | 雪松 | 91.98 | 9.09 |
| 2# | 香樟 | 35.46 | 3.44 | 7# | 雪松 | 127.32 | 30.23 |
| 3# | 雪松 | 108.54 | 24.75 | 8# | 雪松 | 183.33 | 31.22 |
| 4# | 雪松 | 93.85 | 11.71 | 9# | 杨树 | 49.74 | 1.31 |
| 5# | 雪松 | 42.08 | 2.74 | 10# | 杨树 | 249.51 | 51.85 |
林分尺度上香樟、雪松1、雪松2以及杨树冠层蒸腾量的日变化与单株样树的日变化趋势大致相同, 均呈昼高夜低的总体变化趋势(图 3, 图 4), 但不同树种林分尺度上的蒸腾量大小与变化幅度却存在明显差异(图 4)。香樟的平均冠层蒸腾量最小, 为131.83 mm/h, 冠层蒸腾在7:00启动, 9:00—16:00间维持在高值(最大值为283.56 mm/h), 拥有宽且平坦的平顶峰(约7小时), 16:00以后蒸腾量开始快速下降; 雪松1的平均冠层蒸腾量为154.86 mm/h, 冠层蒸腾也在7:00左右启动, 但仅在10:00—13:00维持在高值(最大值为390.77 mm/h), 相对于香樟而言峰值较大, 但峰宽明显收窄且不平坦; 雪松2的平均冠层蒸腾量为150.98 mm/h, 与雪松1相比, 冠层蒸腾启动稍微晚一些, 在8:00左右启动, 13:00左右到达峰值(最大值为437.51 mm/h), 峰现时间明显偏后; 杨树的平均冠层蒸腾量最大, 为267.08 mm/h, 明显高于其他两个树种, 冠层蒸腾在11:00左右到达峰值(最大值为561.46 mm/h), 峰宽大(11:00—19:00, 8 h), 第一天平峰现象很明显, 后两天峰不平坦, 其冠层蒸腾到达峰值后开始下降的时间(19:00)和接近0值的时间(3:00—6:00)均明显晚于其他两个树种。综上所述, 林分尺度上3个树种的冠层蒸腾量从大到小依次为杨树、雪松、香樟, 杨树的峰或谷出现时间均明显晚于雪松和香樟。

|
| 图 4 林分尺度冠层蒸腾量日变化特征 Fig. 4 Diurnal variable characteristics of stand canopy transpiration |
3个树种蒸腾降温值的日变化曲线呈昼低夜高的变化规律, 与其林分尺度上冠层蒸腾量的日变化曲线呈镜像关系, 表明降温效应与冠层蒸腾量具有显著相关性(图 4, 图 5)。3个树种在夜间(20:00—次日6:00)降温值均很小, 在3:00—6:00时段接近于0, 表明降温效应在夜间很不明显; 7:00左右降温值开始降低(即降温效应开始增强), 10:00—14:00左右降温值达到最小, 降温效应最大(香樟:-0.51℃, 雪松1:-0.71℃, 雪松2:-0.79℃, 杨树:-0.93℃), 表明最大蒸腾降温效应杨树>雪松>香樟; 3个树种均具有较为明显的宽谷特征, 表明在高温天气下植被在正午前后进入“午休”, 蒸腾量不再快速增长, 蒸腾降温效应也不再增强。

|
| 图 5 不同树种蒸腾降温效应日变化特征 Fig. 5 Diurnal variable characteristics of transpiration cooling effect |
由图 6可见, 3个树种的遮荫降温效应与太阳辐射的日变化规律基本一致; 在夜间(19:00—次日6:00)因无太阳辐射, 3个树种均无遮荫降温效应; 日出(6:00)之后, 3个树种的遮荫降温效应总体上随着太阳辐射的增强而不断增大, 在正午前后(11:00—13:00)降温效应达到最大, 随后遮荫降温效应随着太阳辐射的减弱而不断减小。另外, 与蒸腾降温效应相比(图 5), 3个树种遮荫降温效应的总体差异相对较小(图 6), 香樟、雪松1、雪松2和杨树遮荫降温效应的最大值分别为-2.48、-2.77、-2.39℃和-2.82℃。由于观测仪器位于绿地单元内的位置不同, 加之太阳天顶角的变化以及传感器上方树木浓密程度的差异, 会造成一些时刻遮荫效果的剧烈变化。例如, 香樟冠层下的观测仪器在12:20左右因接收了大量太阳辐射, 其降温效应急剧下降(仅为-0.41℃), 与其最大遮荫降温效应(12:00左右, -2.48℃)相差很大。

|
| 图 6 遮荫降温效应与太阳辐射日变化特征 Fig. 6 Diurnal variable characteristics of cooling effect of the shade and solar radiation |
由图 7可见, 在夜间(19:00—次日6:00)无论是3个树种还是整个小型绿地单元的总降温效应均非常微弱, 且全部为蒸腾降温; 在白天(6:00—19:00)3个树种的总降温效应峰值存在一定差异(杨树>雪松>香樟), 但其总体上均先增后减, 且遮荫降温对总降温的贡献率(60%—75%)均明显高于蒸腾降温(25%—40%)。香樟在9:00—15:00时段的总降温效应最强(最大为-2.78℃), 该时段内蒸腾降温约占总降温的23%;雪松1的最大总降温效应略高于香樟, 为-3.11℃, 在9:00—15:00时段蒸腾降温占总降温的比重(25%)与香樟的比较接近; 雪松2的总降温效应略低于雪松1, 为-3.06℃, 在10:00—16:00时段的总降温效应最强, 该时段蒸腾降温占总降温比重(36%)高于香樟与雪松1, 这与雪松2在该时段的蒸腾降温高于香樟与雪松1有关(图 5); 杨树的最大总降温效应大于香樟和雪松, 为-3.53℃, 且白天的总降温效应起伏波动较大(这与其遮荫效应的较大起伏波动有关(图 6)), 在9:00—15:00时段蒸腾降温占总降温比重为32%;整个小型绿地单元的最大总降温效应为-3.00℃, 在10:00—15:00时段的总降温效应最强, 该时段蒸腾降温占总降温的29%, 整个观测时期内的蒸腾降温占总降温的34%(图 5)。这一结果与Hoelscher等[3]对夏季晴朗天气下地锦(P. tricuspidata)与巴尔德楚藤蓼(F. baldschuanica)的遮荫降温和蒸腾降温效应的研究结果基本一致, 即蒸腾降温所占总降温效应的比重均低于40%。

|
| 图 7 总降温效应以及遮荫、蒸腾对总降温效应的贡献率 Fig. 7 Total cooling effects and the contribution of transpiration cooling effect and cooling effect of the shade to total cooling effect of different tree species |
本文使用美国Dynamax标准茎流观测系统、RIEGL VZ-400三维激光扫描成像系统和Hobo U30与CR1000微气象观测系统, 对南京市某一小型绿地单元内3个树种的树干液流、冠层结构和微气象环境因子进行了观测与分析, 在揭示高温晴朗天气下单株样树蒸腾量日变化特征的基础上, 采用“单位叶面积上的平均液流速率×叶面积指数”的扩展方法实现了由单株到林分尺度上冠层蒸腾量与蒸腾降温效应的估算, 并根据林上、林下太阳辐射值计算了不同树种与整个绿地单元的遮荫降温效应, 进而阐明了蒸腾与遮荫降温对总降温效应贡献率的变化特征。主要结论如下:
(1) 3个树种的树干液流均呈现昼高夜低的总体变化趋势, 树干液流通常在6:00左右启动, 正午前后达到峰值, 且在正午前后存在“午休”现象(平顶峰), 而在同一树种内树干液流会随着样树胸径的增大而增大。
(2) 林分尺度上冠层蒸腾量与蒸腾降温效应均为杨树>雪松>香樟, 杨树峰或谷出现的时间(11:00—19:00)均明显晚于雪松(10:00—15:00)和香樟(9:00—16:00)。
(3) 林分尺度上3个树种的遮荫降温效应总体上与太阳辐射的日变化规律基本一致, 均随着太阳辐射的增强而不断增大, 正午前后遮荫降温效应达到最大之后开始随着太阳辐射的降低而快速下降; 最大遮荫降温效应:杨树>雪松>香樟, 但与3个树种的蒸腾降温效应相比, 树种间遮荫降温效应的总体差异相对较小。
(4) 3个树种与整个小型绿地单元的总降温效应在夜间均非常微弱, 且全部为蒸腾降温, 而在白天遮荫降温对总降温的贡献率(60%—75%)则明显高于蒸腾降温(25%—40%); 最大总降温效应:杨树>雪松>香樟。
本文通过实地观测定量分析了不同树种的蒸腾与遮荫降温效应的变化特征, 并阐明了蒸腾与遮荫对总降温效应的贡献率。虽然得到了一些初步结论, 但仍存在一些不足:(1)本文仅选取了高温晴朗天气下3 d的观测数据进行了对比分析, 不同天气条件下蒸腾与遮荫降温及其对总降温效应贡献率的差异仍需进一步探讨; (2)由于TDP探针数量与线缆长度的限制, 分两次对3个树种进行了树干液流的观测, 尽管都选择在气象条件相近的高温天气下观测, 但两次观测期间的微气象条件差异难免会对3个树种的比较分析产生一定的影响。(3)在树干液流观测期间, 虽然同步进行了气象因子与土壤环境因子(土壤温湿度、土壤热通量等)的观测, 但本文尚未进行降温效应与微气象因子的相关性分析, 也未考虑土壤蒸散发对绿地降温效应的潜在影响。
| [1] | Dimoudi A, Nikolopoulou M. Vegetation in the urban environment: microclimatic analysis and benefits. Energy and Buildings, 2003, 35(1): 69–76. DOI:10.1016/S0378-7788(02)00081-6 |
| [2] | Sugawara H, Shimizu S, Hagiwara S, Narita K I, Mikami T. How much does urban green cool town?. Journal of Heat Island Institute International, 2014, 9(2): 11–18. |
| [3] | Hoelscher M H, Nehls T, Jänicke B, Wessolek G. Quantifying cooling effects of facade greening: shading, transpiration and insulation. Energy and Buildings, 2016, 114: 283–290. DOI:10.1016/j.enbuild.2015.06.047 |
| [4] | Tsiros I X. Assessment and energy implications of street air temperature cooling by shade tress in Athens (Greece) under extremely hot weather conditions. Renewable Energy, 2010, 35(8): 1866–1869. DOI:10.1016/j.renene.2009.12.021 |
| [5] | Lin B S, Lin Y J. Cooling effect of shade trees with different characteristics in a subtropical urban park. HortScience, 2010, 45(1): 83–86. |
| [6] | Simpson J R, McPherson E G. Potential of tree shade for reducing residential energy use in California. Journal of Arboriculture, 1996, 22(1): 10–18. |
| [7] | Konarska J, Uddling J, Holmer B, Lutz M, Lindberg F, Pleijel H, Thorsson S. Transpiration of urban trees and its cooling effect in a high latitude city. International Journal of Biometeorology, 2016, 60(1): 159–172. DOI:10.1007/s00484-015-1014-x |
| [8] | Chabot R, Bouarfa S, Zimmer D, Chaumont C, Moreau S. Evaluation of the sap flow determined with a heat balance method to measure the transpiration of a sugarcane canopy. Agricultural Water Management, 2005, 75(1): 10–24. DOI:10.1016/j.agwat.2004.12.010 |
| [9] | Green S, Clothier B, Jardine B. Theory and practical application of heat pulse to measure sap flow. Agronomy Journal, 2003, 95(6): 1371–1379. DOI:10.2134/agronj2003.1371 |
| [10] | Bosch D D, Marshall L K, Teskey R. Forest transpiration from sap flux density measurements in a southeastern Coastal Plain riparian buffer system. Agricultural and Forest Meteorology, 2014, 187(8): 72–82. |
| [11] | Čermák J, Kučera J, Nadezhdina N. Sap flow measurements with some thermodynamic methods, flow integration within trees and scaling up from sample trees to entire forest stands. Trees, 2004, 18(5): 529–546. |
| [12] | Juhász Á, Hrotkó K. Comparison of the transpiration part of two sources evapotranspiration model and the measurements of sap flow in the estimation of the transpiration of sweet cherry orchards. Agricultural Water Management, 2014, 143: 142–150. DOI:10.1016/j.agwat.2014.06.014 |
| [13] | Granier A. A new method of sap flow measurement in tree stems. Annals of Forest Science, 1985, 42(2): 193–200. DOI:10.1051/forest:19850204 |
| [14] | 于萌萌, 张新建, 袁凤辉, 何秀, 关德新, 王安志, 吴家兵, 金昌杰. 长白山阔叶红松林三种树种树干液流特征及其与环境因子的关系. 生态学杂志, 2014, 33(7): 1707–1714. |
| [15] | 赵平, 饶兴权, 马玲, 蔡锡安, 曾小平. 基于树干液流测定值进行尺度扩展的马占相思林段蒸腾和冠层气孔导度. 植物生态学报, 2006, 30(4): 655–665. |
| [16] | Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309–320. DOI:10.1093/treephys/3.4.309 |
| [17] | Zhu L W, Zhao P. Temporal variation in sap-flux-scaled transpiration and cooling effect of a subtropical Schima superba plantation in the urban Area of Guangzhou. Journal of Integrative Agriculture, 2013, 12(8): 1350–1356. DOI:10.1016/S2095-3119(13)60548-1 |
| [18] | Soegaard H, Boegh E. Estimation of evapotranspiration from a millet crop in the Sahel combining sap flow, leaf area index and eddy correlation technique. Journal of Hydrology, 1995, 166(3/4): 265–282. |
| [19] | Nakazato T, Inagaki T. Analysis of plant function as bio-thermal-conditioner using Pothos (Epipremnum aureum). Thermochimica Acta, 2012, 532: 49–55. DOI:10.1016/j.tca.2011.09.010 |
| [20] | 白岩, 朱高峰, 张琨, 马婷. 基于树干液流及涡动相关技术的葡萄冠层蒸腾及蒸散发特征研究. 生态学报, 2015, 35(23): 7821–7831. |
| [21] | Granier A, Huc R, Barigah S T. Transpiration of natural rainforest and its dependence on climatic factors. Agricultural and Forest Meteorology, 1996, 78(1/2): 19–29. |
| [22] | Solberg S. Mapping gap fraction, LAI and defoliation using various ALS penetration variables. International Journal of Remote Sensing, 2010, 31(5): 1227–1244. DOI:10.1080/01431160903380672 |
| [23] | 骆社周, 王成, 张贵宾, 习晓环, 李贵才. 机载激光雷达森林叶面积指数反演研究. 地球物理学报, 2013, 56(5): 1467–1475. DOI:10.6038/cjg20130505 |
| [24] | 刘鑫, 张金池, 谢德晋, 庄家尧, 邵永昌, 张水锋. 南京近郊麻栎林冠层蒸腾时间变化及降温效应. 浙江农林大学学报, 2015, 32(4): 529–536. DOI:10.11833/j.issn.2095-0756.2015.04.006 |
| [25] | Rahman M A, Moser A, Rötzer T, Pauleit S. Within canopy temperature differences and cooling ability of Tilia cordata trees grown in urban conditions. Building and Environment, 2017, 114: 118–128. DOI:10.1016/j.buildenv.2016.12.013 |
| [26] | 杨士弘. 城市绿化树木的降温增湿效应研究. 地理研究, 1994, 13(4): 74–80. |
| [27] | 吉野正敏. 局地气候原理. 郭可展, 李师融, 译. 南宁: 广西科学技术出版社, 1989. |