 2018, Vol. 38
2018, Vol. 38文章信息
- 郭萌萌, 姜文虎, 李探, 吴龙飞, 刘军侠.
- GUO Mengmeng, JIANG Wenhu, LI Tan, WU Longfei, LIU Junxia.
- 转双抗虫基因107杨节肢动物群落特征及相对稳定性
- Arthropod community characteristics and relative stability in transgenic Populus×euramericana 'Neva' carrying bivalent insect-resistant genes
- 生态学报. 2018, 38(1): 333-341
- Acta Ecologica Sinica. 2018, 38(1): 333-341
- http://dx.doi.org/10.5846/stxb201612202623
-
文章历史
- 收稿日期: 2016-12-20
- 网络出版日期: 2017-09-13

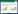




2. 河北省林木种质资源与森林保护重点实验室, 保定 071000
2. Hebei Key Lab of Forest Germplasm Resources and Protection, Baoding 071000, China
随着植物转基因工程研究的快速发展, 大量转基因植物已经进入了大田试验阶段, 并且部分转基因植物已商品化进入市场[1-3]。与此同时, 转基因植物向环境释放后可能带来的生态风险渐渐引起社会各界的关注[4-7]。Bt基因由于在抗虫方面表现突出, 所以应用范围最广, 随着转Bt抗虫基因植物进入大规模的商业化应用阶段, 其环境释放后可能存在的风险不断有报道, 如转Bt基因抗虫棉对目标害虫具备高抗虫性的优点, 在治理抗药性棉铃虫中发挥了重要作用, 同时提高了捕食性天敌的数量, 但是, 转Bt基因棉田间棉叶螨、棉蚜、棉盲蝽等次要害虫的发生危害加重[8];转基因玉米的花粉对大斑蝶(Danaus plexippus)的幼虫生长发育造成影响[9];转基因马铃薯会对幼鼠的生长发育造成严重影响并损害其免疫系统[10]。这都表明转基因植物的应用给人类带来了极大的经济和社会效益, 同时, 在生态环境方面可能存在的风险更不能忽视, 生物安全性问题已成为商业化应用转基因植物道路上最大的障碍[11]。
由于转基因树木的生长周期较长, 连续多年的在田间进行大面积种植对生态系统可能存在潜在的风险, 其生物安全问题越来越受到公众的重视[12-13]。近年来, 对抗虫转基因杨生态安全性的研究多集中于其对靶标和非靶标害虫, 以及天敌种群或节肢动物群落的影响, 并取得了较大进展[14-15]。以往研究表明, 转抗虫基因杨对节肢动物群落有一定影响。如转抗虫基因741杨对靶标害虫具有较好的抗性, 节肢动物群落总体物种丰富度增加[16-17];转Bt基因欧洲黑杨提高了寄生于靶标害虫的天敌数量、种类和寄生率[18];表明转抗虫基因杨对靶标害虫毒害作用明显, 具有较高的抗性, 对非靶标生物毒害作用不显著, 并且对天敌种群的数量有一定的促进作用。
杨树(Populus L.)作为木本植物研究中的模式植物, 是全球重要的栽培树种, 在我国是重要的速生丰产林、生态防护林和道路绿化树种。近年来, 杨树食叶害虫经常爆发成灾, 蛀干性害虫也时有发生, 造成了极大地损失, 而且大规模的施用农药已经让害虫产生了一定的抗药性。转抗虫基因107杨的成功获取, 不仅能有效地解决杨树的虫害问题, 而且该树种的速生性也将使其成为道路绿化及经济林应用的首选树种, 因此, 它的研发和培育在社会需求和经济效益上均是必要的[19]。转抗虫基因107杨是由河北农业大学生物技术研究所将部分改造的Bt杀虫蛋白基因(BtCry1Ac)与慈菇蛋白酶抑制基因(API基因)构建成双抗虫基因表达载体, 通过根癌农杆菌介导法, 共同转入到优良的未转基因的107杨(Populus × euramericana ‘Neva’)中, 再经过多次筛选及检测以后获得不同系号的转抗虫基因107杨无性系。本研究在以往转抗虫基因107杨抗虫性研究基础之上, 首次对中试试验林地节肢动物群落进行监测和系统分析, 以期为转抗虫基因杨树的生态安全性评价提供数据支撑, 为转抗虫基因107杨的推广与应用提供参考。
1 材料与方法 1.1 试验材料未转基因107杨(对照杨)和转抗虫基因107杨是由河北农业大学生物技术研究所提供。转抗虫基因107杨的系号分别为#3、#2、#1、#9, 树龄:2年生。
1.2 试验地概况试验地位于唐山市滦南县苗圃场, 试验林内种植有对照杨和转抗虫基因107杨的不同系号, 分别是#3、#2、#1、#9。试验林采用随机区组设计, 每个系号为一个小区, 种植30株, 4次重复, 株距2 m, 行距5 m。整块试验地地势、地貌、土质、气温、降水等自然条件和人为管理均一致。试验林地除进行正常的水肥及基本管理措施外, 不施用任何农药。
1.3 调查方法节肢动物群落调查于2015年5—9月进行, 每15—20天调查1次(共计8次)。采取随机抽样的方法, 在区组中分别随机选取5株样树, 在样株东、南、西、北4个方位, 按上、下两个层次各选取50 cm长的枝条, 记录其上节肢动物的种类和数量;调查距地表 3 m内的主干上节肢动物的种类和数量;在各区组地表随机选取5个1 m×1 m的样方, 调查取样范围内节肢动物的种类及数量, 并作详细记载。未知种类装入毒瓶或酒精试管, 按统一标号进行编号, 带回实验室进行鉴定[20]。特殊处理:对于样枝上的螨类、蚜虫类等, 选取此样枝顶尖倒数第3片叶子上的虫口数量进行统计。
1.4 功能团的划分Root提出集团的概念[21], 他认为集团是以相似方式利用相同等级的生境资源的一个类群中的物种。因此可以将群落中的物种划分为多个取食行为、利用资源和生境选择相似的集合体[17]。本研究主要基于节肢动物的取食习性将转抗虫基因107杨和对照杨节肢动物群落中所有节肢动物划分为食叶昆虫、刺吸昆虫、其他植食性昆虫、捕食性天敌、寄生性天敌和中性昆虫等6个功能团。
1.5 数据分析方法(1) 相对多度
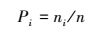
|
式中:ni为样本中第i个样本的个体数量, n为样本的所有的个体数量总和。
(2) 群落特征指数分析
采用以下公式分析群落特征指数[22]。
ⅠShannon-Wiener多样性指数(Diversity Index)
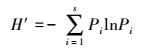
|
式中,S为物种数;Pi为相对多度。
ⅡPielou均匀度指数(Evenness Index)

|
式中, H′为Shannon-Wiener多样性指数;Hmax为给定物种完全均匀时的群落多样性指数;S为物种数。
ⅢBerger-Parker优势度指数(Dominance Index)
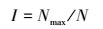
|
式中, Nmax为优势类群的数量;N为群落中总的个数。
(3) 群落相似性分析采用Jaccard指数Cj[23]
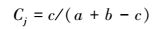
|
式中:c为两样地相同的物种数;a、b分别为两样地各自物种数。该公式规定相似性等级是:当Cj=0.00—0.25时, 极不相似;当Cj=0.25—0.50时, 中等不相似;当Cj=0.50—0.75时, 中等相似;当Cj=0.75—1.00时, 极相似。
(4) 群落相对稳定性分析
采用St/Si即群落内物种数与群落内个体数之比, 可反映种间数量上的制约作用;Sn/Sp即天敌类群物种数与植食性类群物种数之比, 可以反映食物网关系和天敌—害虫相互制约的复杂程度[24]。
(5) 主成分分析
以各类功能团节肢动物的物种数量和个体数量为因子, 作主成分分析, 并计算各因子的特征根向量、各主成分的累积贡献率和负荷量[25]。
1.6 数据处理方法本文使用Forstat和DPS软件对转抗虫基因107杨林中节肢动物群落特征指数进行处理及差异性显著分析;采用SPSS软件对转抗虫基因107杨林中节肢动物群落进行主成分分析;利用Excel软件进行作图、作表。
2 结果与分析 2.1 转抗虫基因107杨对节肢动物群落组成的影响根据本次调查统计, 共有节肢动物6818头, 隶属于2纲分别为昆虫纲和蛛形纲, 8目(分别为鳞翅目Lepidoptera、鞘翅目Coleoptera、膜翅目Hymenoptera、双翅目Diptera、半翅目Hemiptera、脉翅目Neuroptera、直翅目Orthoptera、蜘蛛目Araneida), 43科, 58种。其中害虫29种, 分属5个目, 其中发生量较大的有美国白蛾Hyphantria cunea(Drury)、杨扇舟蛾Clostera anachoreta (Fabricius)、杨银叶潜蛾Phyllocnistis saligna Zeller;天敌有18种, 分属5个目, 以瓢虫科Coccinellidae数量最多;中性昆虫有11种, 隶属于3个目, 以双翅目为主。将主要节肢动物物种数及个体数进行统计, 并进行差异性显著分析及个体数相对多度的计算, 结果见表 1。
| 所属目Groups | #3 | #2 | #1 | #9 | CK | ||||||||||
| NS | NI | Pi | NS | NI | Pi | NS | NI | Pi | NS | NI | Pi | NS | NI | Pi | |
| 鳞翅目Lepidoptera | 6a | 154a | 44.78 | 6a | 49a | 20.74 | 6a | 117a | 41.31 | 6a | 108a | 40.22 | 7a | 403b | 70.06 |
| 鞘翅目Coleoptera | 6a | 58a | 16.85 | 6a | 46a | 19.58 | 5a | 47a | 16.68 | 5a | 44a | 16.28 | 6a | 46a | 8.05 |
| 膜翅目Hymenoptera | 4a | 60a | 17.51 | 5a | 51a | 21.69 | 3a | 51a | 18.09 | 4a | 51a | 18.99 | 5a | 57a | 9.88 |
| 双翅目Diptera | 6a | 33a | 9.63 | 7a | 46a | 19.58 | 6a | 34a | 12.00 | 6a | 33a | 12.44 | 7a | 39a | 6.79 |
| 半翅目Hemiptera | 5a | 26ab | 7.66 | 5a | 30a | 12.70 | 5a | 23ab | 8.21 | 5a | 20ab | 7.30 | 5a | 16b | 2.74 |
| 脉翅目Neuroptera | 1a | 4a | 1.09 | 1a | 4a | 1.80 | 1a | 2a | 0.62 | 1a | 3a | 1.12 | 1a | 4a | 0.74 |
| 直翅目Orthoptera | 0a | 0a | 0.00 | 1a | 1a | 0.32 | 1a | 1a | 0.26 | 1a | 1a | 0.28 | 1a | 1a | 0.13 |
| 蜘蛛目Araneida | 4a | 9a | 2.48 | 4a | 9a | 3.60 | 5a | 8a | 2.82 | 5a | 9a | 3.37 | 5a | 9a | 1.61 |
| 总计Total | 343 | 100.00 | 236 | 100.00 | 283 | 100.00 | 267 | 100.00 | 575 | 100.00 | |||||
| NS:物种数Number of species;NI:个体数Number of individuals;Pi为相对多度Relative abundance;同一行不同小写字母代表同一指标不同株系间在0.05水平下存在显著性差异 | |||||||||||||||
由表 1可以看出, 转抗虫基因107杨与对照杨, 从目的类别上看, #3无直翅目昆虫, 只有7个目, 其他株系均为8个目, 在物种数量上与对照杨不存在显著差异。从个体数量上看, 鳞翅目、鞘翅目、膜翅目、双翅目是转抗虫基因107杨和对照杨节肢动物群落的主要类群, 其中鳞翅目昆虫数量最多;转抗虫基因107杨鳞翅目昆虫个体数较对照显著减少, 半翅目昆虫个体数均多于对照杨, 其中#2显著增加, 其他目的个体数与对照杨不存在显著差异。从相对多度上看, 对照杨鳞翅目昆虫的相对多度高达70.06%, 转抗虫基因107杨均低于50%, 其中#2最低, 为20.74%;鞘翅目、膜翅目、双翅目、半翅目、蜘蛛目的相对多度均高于对照杨, 其中#2的相对多度均最高。
功能团的划分可以直观的了解节肢动物群落结构的组成情况。转抗虫基因107杨节肢动物群落各功能团的物种数与个体数统计结果见表 2。
| 株系 Strains |
食叶昆虫 Herbivorous insects |
刺吸昆虫 Stick suction insects |
其他植食性昆虫 Other phytophagous insects |
捕食性天敌 Predacious insects |
寄生性天敌 Parasitic insects |
中性昆虫 Neutral insects | ||||||
| NS | NI | NS | NI | NS | NI | NS | NI | NS | NI | NS | NI | |
| #3 | 8a | 186a | 5a | 26ab | 3a | 10ab | 8a | 75a | 2a | 10a | 6a | 36a |
| #2 | 8a | 80a | 5a | 30a | 3a | 9a | 9a | 63a | 3a | 8a | 7ab | 47a |
| #1 | 8a | 140a | 5a | 23ab | 3a | 13ab | 9a | 65a | 2a | 5a | 6ab | 38a |
| #9 | 7a | 133a | 5a | 20ab | 2a | 10ab | 7a | 63a | 2a | 6a | 7ab | 36a |
| CK | 9a | 425b | 5a | 16b | 3a | 20b | 8a | 58a | 2a | 13a | 8b | 43a |
| NS:物种数Number of species;NI:个体数Number of individual;同一列不同小写字母代表同一指标不同株系间在0.05水平下存在显著性差异 | ||||||||||||
由表 2可知, 转抗虫基因107杨节肢动物群落与对照杨在功能类群上组成一致, 只有#3的中性昆虫物种数与对照杨存在差异。转抗虫基因107杨中食叶昆虫个体数较对照显著减少;刺吸昆虫个体数均多于对照杨, 其中#2较对照杨刺吸昆虫虫口数显著增加;其他植食性昆虫个体数数均少于对照杨, #2较对照显著减少;捕食性天敌、寄生性天敌及中性昆虫个体数与对照杨相似, 差异不显著。比较转抗虫基因107杨各株系, #2的食叶昆虫、其他植食性昆虫个体数最少, 刺吸昆虫的个体数最多, 捕食性天敌数量较对照有所增加, 中性昆虫数量最多。
2.2 转抗虫基因107杨节肢动物群落特征指数转抗虫基因107杨节肢动物群落特征指数见图 1。

|
| 图 1 转抗虫基因107杨节肢动物群落特征指数 Fig. 1 Characteristic indexes of arthropod community in transgenic insect-resistant 107 poplars 图中不同小写字母表示同一指标不同株系间在0.05水平下存在显著性差异 |
由图 1可见, 转抗虫基因107杨的多样性指数和均匀性指数均高于对照杨, 其中#2较对照杨显著增高, 其他株系与对照杨差异不显著;从优势度指数上看, 转抗虫基因107杨低于对照杨, 其中#2较对照杨显著降低。调查结果显示转抗虫基因107杨与对照杨中主要优势类群有灯蛾科Arctiidae、舟蛾科Notodontidae、卷叶蛾科Tortricidae、叶甲科Chrysomelidae等, 转抗虫基因107杨优势度指数降低, 说明优势类群的地位下降。
2.3 转基因107杨节肢动物群落相似性分析由表 3可知, 转抗虫基因107杨节肢动物群落与对照杨相似度较高, 其中除#1为中等相似外, 其他株系与对照杨的相似性系数均大于0.75, 为极相似, #9与对照杨节肢动物群落相似性最大, 相似性系数高达0.8039;比较转抗虫基因107杨各株系间相似性系数, 除#3与#9为中等相似外, 其他株系间相似性系数均大于0.75, 表示其他株系间节肢动物群落极相似。由此表明转抗虫基因107杨对节肢动物群落物种组成无明显作用。
| 株系Strains | #3 | #2 | #1 | #9 | CK |
| #3 | 1 | 0.8462 | 0.8077 | 0.7170 | 0.7925 |
| #2 | 1 | 0.7778 | 0.7885 | 0.7636 | |
| #1 | 1 | 0.7843 | 0.6964 | ||
| #9 | 1 | 0.8039 | |||
| CK | 1 |
转抗虫基因107杨节肢动物群落相对稳定性分析结果见图 2。

|
| 图 2 转抗虫基因107杨节肢动物群落相对稳定性 Fig. 2 The relative stability value of arthropod community in transgenic insect-resistant 107 poplars 图中不同小写字母表示同一指标不同株系间在0.05水平下存在显著性差异 |
由图 2可以看出, St/Si的比值排序为#2>#9>#1>#3>CK, 转抗虫基因107杨的St/Si比值均高于对照杨, 其中#2、#9与对照杨差异显著, 表明转抗虫基因107杨节肢动物群落物种数相对较多, 个体数相对较少, 反映转抗虫基因107杨节肢动物群落物种间在数量上的制约作用较强。转抗虫基因107杨与对照杨的Sn/Sp的比值排序为#1>#2>#9>CK>#3, 但差异不显著, 表明转抗虫基因107杨与对照杨在天敌物种与植食性物种比例上无显著差异。
2.5 转抗虫基因107杨林节肢动物群落主成分分析主成分分析方法明确了转抗虫基因107杨及对照杨林内节肢动物群落的主要构成因素以及各因素之间的相互关系, 结果见表 4。
| 株系 Strains |
成分 Component |
因子载荷量Factor Loading | 累计贡献率 Cumulative/% |
|||||||||||
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 | |||
| #3 | Ⅰ | -0.006 | 0.125 | 0.186 | 0.101 | 0.186 | 0.073 | 0.187 | 0.013 | -0.029 | 0.175 | 0.107 | -0.088 | 45.27 |
| Ⅱ | 0.279 | 0.120 | -0.017 | 0.218 | -0.017 | -0.295 | -0.068 | -0.041 | 0.178 | -0.034 | 0.080 | 0.154 | 74.25 | |
| Ⅲ | -0.018 | -0.137 | -0.015 | 0.071 | -0.015 | -0.113 | -0.060 | 0.301 | -0.197 | 0.083 | 0.248 | 0.305 | 100.00 | |
| #2 | Ⅰ | 0.159 | 0.157 | 0.164 | 0.134 | 0.030 | -0.062 | 0.125 | 0.033 | 0.058 | -0.044 | 0.144 | 0.146 | 50.89 |
| Ⅱ | 0.057 | -0.089 | -0.014 | 0.152 | 0.279 | 0.292 | -0.123 | 0.239 | 0.016 | -0.040 | -0.143 | 0.121 | 79.68 | |
| Ⅲ | 0.008 | 0.130 | 0.010 | -0.187 | -0.004 | -0.101 | -0.129 | 0.095 | 0.362 | 0.399 | 0.199 | -0.130 | 100.00 | |
| #1 | Ⅰ | -0.073 | 0.027 | 0.079 | 0.121 | 0.198 | 0.242 | -0.049 | -0.138 | 0.019 | 0.167 | -0.029 | 0.257 | 39.37 |
| Ⅱ | 0.261 | 0.265 | 0.249 | -0.030 | 0.000 | -0.011 | 0.079 | -0.051 | -0.099 | -0.025 | 0.215 | 0.015 | 71.05 | |
| Ⅲ | 0.085 | 0.056 | 0.000 | -0.339 | 0.010 | -0.102 | 0.308 | -0.116 | 0.207 | 0.061 | -0.085 | -0.185 | 100.00 | |
| #9 | Ⅰ | -0.024 | 0.165 | -0.172 | -0.140 | 0.172 | -0.023 | 0.066 | 0.015 | 0.172 | -0.068 | -0.083 | -0.138 | 49.37 |
| Ⅱ | -0.021 | -0.009 | 0.032 | 0.202 | -0.032 | 0.248 | 0.205 | 0.073 | -0.032 | 0.261 | -0.175 | -0.083 | 82.48 | |
| Ⅲ | 0.458 | 0.077 | 0.002 | 0.006 | -0.002 | -0.045 | 0.127 | 0.470 | -0.002 | 0.081 | 0.083 | 0.115 | 100.00 | |
| CK | Ⅰ | -0.006 | 0.120 | 0.230 | 0.239 | -0.048 | -0.011 | 0.137 | -0.024 | 0.139 | 0.224 | -0.007 | -0.153 | 35.07 |
| Ⅱ | 0.209 | 0.166 | -0.103 | 0.026 | -0.107 | -0.140 | 0.262 | 0.330 | -0.277 | 0.078 | -0.035 | 0.125 | 69.54 | |
| Ⅲ | -0.001 | 0.361 | -0.034 | 0.075 | 0.106 | -0.356 | 0.137 | 0.159 | -0.126 | 0.035 | 0.193 | 0.263 | 100.00 | |
| X1:食叶昆虫物种数;X2:食叶昆虫个体数;X3:刺吸昆虫物种数;X4:刺吸昆虫个体数;X5:其他植食物种数;X6:其他植食性昆虫个体数;X7:捕食性天敌物种数;X8:捕食性天敌个体数;X9:寄生性天敌的物种数;X10:寄生性天敌个体数;X11:中性昆虫物种数;X12:中性昆虫个体数。Ⅰ:第一主成分;Ⅱ:第二主成分;Ⅲ:第三主成分 | ||||||||||||||
由表 4可知, 转抗虫基因107杨与对照杨林节肢动物群落中均提取了3个主成分, 且累计贡献率均高达100%, 足以概括组成昆虫群落的主要因子。
#3杨林的第Ⅰ主成分代表捕食性天敌物种数量、其他植食性昆虫物种数量、刺吸昆虫物种数量和食叶昆虫个体数量的综合因子, 累计贡献率较高, 为45.27%;第Ⅱ主成分代表其他植食性昆虫个体数量和食叶昆虫物种数量的综合因子;第Ⅲ主成分代表中性昆虫个体数量与捕食性天敌个体数量的综合因子。说明在#3中, 捕食性天敌物种数量、其他植食性昆虫物种数量、刺吸昆虫物种数量和食叶昆虫个体数量是导致昆虫群落变化的最主要因子, 其他植食性昆虫物种数和个体数、食叶昆虫物种数及中性昆虫的个体数与主要因子共同影响着整个群落的波动。
#2杨林的第Ⅰ主成分代表刺吸昆虫物种数量、食叶昆虫物种数量和个体数量的综合因子;第Ⅱ主成分代表其他植食性物种数量和个体数量、捕食性天敌个体数量的综合因子;第Ⅲ主成分代表寄生性天敌物种数量和个体数量的综合因子。节肢动物群落的不同功能类群均对#2杨林有大小不同的影响, 表明#2杨林的物种分布比较均衡, 也解释了#2杨林节肢动物群落均匀度最高的原因。
#1杨林的第Ⅰ主成分代表其他植食性昆虫和中性昆虫个体数量的综合因子;第Ⅱ主成分代表食叶昆虫物种数量和个体数量、刺吸昆虫物种数量的综合因子;第Ⅲ主成分代表刺吸昆虫的个体数量和捕食性天敌物种数量的综合因子。表明食叶昆虫、刺吸昆虫和捕食性天敌是#1杨林的主导因子。
#9杨林的第Ⅰ主成分代表刺吸昆虫、其他植食性昆虫、寄生性天敌物种数量和食叶昆虫个体数量的综合因子;第Ⅱ主成分代表其他植食性昆虫和寄生性天敌个体数量的综合因子;第Ⅲ主成分代表食叶昆虫物种数量与捕食性天敌个体数量的综合因子。表明食叶昆虫、寄生性天敌、刺吸昆虫共同影响#9杨林节肢动物群落的波动。
对照杨林的第Ⅰ主成分代表刺吸昆虫物种数量和个体数量、寄生天敌个体数量的综合因子, 累计贡献率为35.07%;第Ⅱ主成分代表捕食天敌的物种数量和个体数量、寄生天敌物种数量、食叶昆虫物种数量的综合因子, 累计贡献率为34.07%;第Ⅲ主成分代表食叶昆虫个体数量与其他植食性昆虫个体数量的综合因子, 累计贡献率为30.46%。表明第Ⅰ、Ⅱ、Ⅲ主成分的综合因子均对对照杨林有主导作用。
综合以上分析结果可得出, 影响转抗虫基因107杨和对照杨林节肢动物群落的主要因子存在差异, 转抗虫基因107杨林中, 食叶昆虫物种数量和个体数量、刺吸昆虫物种数量和其他植食性昆虫物种数量是主导节肢动物变化的最主要因子;对照杨林中, 食叶昆虫、刺吸昆虫、寄生性天敌、捕食性天敌物种数量和个体数量、其他植食性昆虫个体数量主导了节肢动物群落的变化。食叶昆虫物种数量与个体数量、刺吸昆虫物种数量和其他植食性昆虫个体数量是影响转抗虫基因107杨与对照杨节肢动物群落变化的共同主导因子。
3 结论与讨论转抗虫基因107杨节肢动物群落特征及相对稳定性是转抗虫基因杨安全性评价的重要内容之一, 是转抗虫基因107杨走向商品化的必经阶段。本文通过对转抗虫基因107杨节肢动物群落的野外调查, 研究了转抗虫基因107杨节肢动物群落的组成、群落特征及群落内部结构关系等, 可以得到如下结论:转抗虫基因107杨可以有效地抑制靶标鳞翅目害虫的个体数量, 具有显著的抗虫效果, 但对非靶标害虫在一定程度上有增殖作用, 对捕食性天敌无不利影响, 甚至还有促进作用;转抗虫基因107杨可在一定程度上提高节肢动物群落的多样性;转抗虫基因107杨与对照杨节肢动物群落间共有种较多, 但转抗虫基因107杨节肢动物群落物种间在数量上的制约作用较强;不同功能类群的物种数与个体数均对转抗虫基因107杨和对照杨林内节肢动物群落的构成有影响, 其中食叶昆虫、刺吸昆虫是影响转抗虫基因107杨与对照杨节肢动物群落变化的共同主导因子。
物种种类和数量是群落基本的组成单元之一, 它们的变化影响着整个群落组成单元的变化[26]。转抗虫基因107杨对鳞翅目害虫的抗性减轻了对杨叶的危害, 从而促进了其他害虫种群的增长, 造成刺吸昆虫个体数量显著增加, 这与张真等[27]、刘万学等[28]等研究结果相似, 张真等研究结果表明Bt转基因杨树会改变其昆虫群落结构, 由于Bt主要作用于鳞翅目昆虫, 而对膜翅目影响不大, 在抑制鳞翅目昆虫的同时, 有利于其他食叶昆虫, 所以转基因纯林中杨叶蜂为优势种, 而在转基因与非转基因混交林中以杨扇舟蛾为优势种;刘万学的研究结果表明转抗虫基因棉花会使次要刺吸昆虫的数量增加;但与姜文虎等[29]对转抗虫基因741杨的研究中刺吸昆虫数量减少的结果不一致。因此, 转抗虫基因植物对刺吸昆虫的影响和作用机制还有待进一步的研究。
转抗虫基因107杨节肢动物群落与对照相比多样性增加, 除#2外, 其他株系与对照杨不存在显著性差异。可能原因有:试验杨林树龄较小, 也可能与试验地规模及栽培模式有关, 对照林与转抗虫基因107杨林较近, 当节肢动物群落受到外界干扰时可能会在转基因与非转基因杨林内相互扩散。
转抗虫基因107杨节肢动物群落是多种植食性昆虫、天敌和中性节肢动物彼此关联、相互制约的复杂网络系统[30], 转基因107杨进入生态系统时, 有可能对靶标害虫、非靶标害虫、天敌的种类组成、数量与发生动态以及群落结构、生物多样性、乃至整个生态过程产生影响, 甚至通过节肢动物的迁移和扩散对周围林分的节肢动物群落造成间接影响。本文说明了转抗虫基因107杨对靶标害虫有很好的抑制作用, 同时提高了节肢动物群落的多样性, 对外界环境的变化或来自群落内部的种群波动的缓冲作用增强。比较转抗虫基因107杨不同株系, #2表现出较高的抗虫性, 多样性指数、均匀性指数、天敌物种数及个体数均明显高于对照杨, 优势度指数明显低于对照杨, 主成分分析结果显示, 节肢动物群落的不同功能类群均对#2杨林有影响, 说明#2杨林的节肢动物物种分布比较均衡。
| [1] | 饶红宇, 陈英, 黄敏仁, 王明庥, 伍宁丰, 范云六. 杨树NL-80106转Bt基因植株的获得及抗虫性. 植物资源与环境学报, 2000, 9(2): 1–5. |
| [2] | 卢孟柱, 胡建军. 我国转基因杨树的研究及应用现状. 林业科技开发, 2006, 20(6): 1–4. |
| [3] | 胡建军, 杨敏生, 卢孟柱. 我国抗虫转基因杨树生态安全性研究进展. 生物多样性, 2010, 18(4): 336–345. |
| [4] | Pilson D, Prendeville H R. Ecological effects of transgenic crops and the escape of transgenes into wild populations. Annual Review of Ecology, Evolution, and Systematics, 2004, 35(1): 149–174. DOI:10.1146/annurev.ecolsys.34.011802.132406 |
| [5] | 卢宝荣, 张文驹, 李博. 转基因的逃逸及生态风险. 应用生态学报, 2003, 14(6): 989–994. |
| [6] | 张永军, 吴孔明, 彭于发, 郭予元. 转基因植物的生态风险. 生态学报, 2002, 22(11): 1951–1959. |
| [7] | 郭建英, 万方浩, 韩召军. 转基因植物的生态安全性风险. 中国生态农业学报, 2008, 16(2): 515–522. |
| [8] | 夏敬源, 崔金杰, 马丽华, 董双林, 崔学芬. 转Bt基因抗虫棉在害虫综合治理中的作用研究. 棉花学报, 1999, 11(2): 57–64. |
| [9] | Losey J E, Rayor L S, Carter M E. Transgenic pollen harms monarch larvae. Nature, 1999, 399(6733): 214–214. |
| [10] | 赵凯, 朱宏, 赵星海, 李鹏, 唐雪明. 转基因植物环境安全性研究进展. 生物学通报, 2009, 44(4): 8–11. |
| [11] | 刘标, 韩娟, 薛堃. 转基因植物环境监测进展. 生态学报, 2016, 36(9): 2490–2496. |
| [12] | Irwin R, Jones P B C. Biosafety of Transgenic Trees in the United States//Landscapes, Genomics and Transgenic Conifers. Netherlands:Springer, 2006: 245–261. |
| [13] | 康向阳, 刘志民, 李胜功. 论转基因林木的潜在生态风险性. 应用生态学报, 2004, 15(7): 1281–1284. |
| [14] | 高宝嘉, 张芳, 侯东永, 吴伯军, 张树平, 赵秀玲. 转基因741杨林节肢动物群落结构的研究. 北京林业大学学报, 2003, 2(1): 62–64. |
| [15] | 张晓芬. 转基因抗虫杨的抗虫性测定及对节肢动物群落影响研究[D]. 北京: 北京林业大学, 2009. http://cdmd.cnki.com.cn/article/cdmd-10022-2009161673.htm |
| [16] | 高素红. 转双抗虫基因741杨节肢动物群落生态安全性评价[D]. 保定: 河北农业大学, 2003. http://cdmd.cnki.com.cn/article/cdmd-11920-2003091388.htm |
| [17] | 高宝嘉, 高素红, 刘军侠, 姜文虎. 转双抗虫基因741杨节肢动物群落营养结构及生态位变化. 生态学报, 2006, 26(10): 3499–3507. |
| [18] | 胡建军, 李淑梅, 卢孟柱, 李继祥, 李开花, 孙雪芹, 赵自玉. 转Bt基因欧洲黑杨抗虫稳定性及其对天敌昆虫的影响. 林业科学研究, 2007, 20(5): 656–659. |
| [19] | 张益文, 任亚超, 刘娇娇, 梁海永, 杨敏生. 转双抗虫基因欧美杨107杨中外源基因的表达. 林业科学, 2015, 51(12): 45–52. |
| [20] | 崔金杰, 夏敬源. 转Bt基因棉对天敌种群动态的影响. 棉花学报, 1999, 11(2): 84–91. |
| [21] | Adams J. The definition and interpretation of guild structure in ecological communities. Journal of Animal Ecology, 1985, 54(1): 43–59. DOI:10.2307/4619 |
| [22] | 戈峰. 昆虫生态学原理与方法. 北京: 高等教育出版社, 2008. |
| [23] | 丁岩钦. 昆虫数学生态学. 北京: 科学出版社, 1994. |
| [24] | 高宝嘉, 张执中, 李镇宇. 封山育林对昆虫群落结构及多样性稳定性影响的研究. 生态学报, 1992, 12(1): 1–7. |
| [25] | 余世孝. 数学生态学导论. 北京: 科学技术文献出版社, 1995. |
| [26] | 姚丽, 高宝嘉. 转基因741杨对节肢动物群落结构的影响. 福建农业学报, 2016, 31(5): 480–486. |
| [27] | 张真, 王军辉, 张建国, 张守攻. Bt转基因杨树对杨树昆虫群落结构的影响. 林业科学, 2004, 40(2): 84–89. |
| [28] | 刘万学, 万方浩, 郭建英. 转Bt基因棉田节肢动物群落营养层及优势功能团的组成与变化. 生态学报, 2002, 22(5): 729–735. |
| [29] | 姜文虎, 刘军侠, 张芳, 高宝嘉. 转基因741杨对靶标和非靶标害虫及主要天敌种群动态的影响. 山东农业大学学报:自然科学版, 2009, 40(2): 195–199. |
| [30] | 李超丽. 转基因杨棉复合系统节肢动物群落变化规律的研究[D]. 保定: 河北农业大学, 2011. http://cdmd.cnki.com.cn/article/cdmd-11920-1011074600.htm |