 2018, Vol. 38
2018, Vol. 38文章信息
- 冯晓钰, 周广胜.
- FENG Xiaoyu, ZHOU Guangsheng.
- 夏玉米叶片水分变化与光合作用和土壤水分的关系
- Relationship of leaf water content with photosynthesis and soil water content in summer maize
- 生态学报. 2018, 38(1): 177-185
- Acta Ecologica Sinica. 2018, 38(1): 177-185
- http://dx.doi.org/10.5846/stxb201612092534
-
文章历史
- 收稿日期: 2016-12-09
- 网络出版日期: 2017-09-12

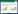




水分是作物的重要组成成分, 是控制作物光合作用和生物量的主要因素之一[1]。叶片水分是进行光合作用的原料, 相比于土壤含水量, 叶片含水量更能直接反映作物生长发育的实际状况, 是机理研究中反映作物水分盈亏程度的最佳指标[2]。特别是, 叶片含水量是大气干燥度、土壤供水能力和作物耐旱能力综合作用的体现, 是监测作物受胁迫和灌溉决策的重要参考, 同时遥感技术的快速发展也为监测叶片水分含量变化提供了强有力的支撑[3-6]。因此, 研究叶片水分含量与光合作用的关系不仅有助于正确理解光合作用变化的机制, 也有助于准确模拟光合作用动态变化。
目前, 关于土壤含水量及其对光合生理生态的研究已有大量报道[7-9]。土壤水分亏缺将引起气孔和非气孔因素限制间接导致光合速率降低[10-13]。叶片净光合速率对土壤水分变化的响应呈二次曲线关系[14-17], 但有关叶片含水量变化及对叶片净光合速率的影响研究较少。王纪华等[18]发现小麦叶片含水量由75%下降到70%左右是叶片光合生理活性的一个重要转折点。作物从土壤中吸收水分以维持自身细胞的生存和正常功能, 且为光合作用提供原料以维持植物生长发育的可持续性, 即叶片含水量不能低于叶片进行光合作用所需的水分, 也就是叶片可能存在一个光合作用为零的叶片水分阈值。张文丽[19]研究发现, 当土壤相对含水量从100%逐渐干旱到90%过程中, 玉米叶片相对含水量变化趋势平缓, 基本保持在一个水平上;当土壤相对含水量干旱到60%, 叶片相对含水量随土壤相对含水量的降低迅速降低, 变化几近于直线, 表明叶片并不能无限制地从土壤获取水分, 即叶片可能存在一个最大水分阈值。然而由于以往试验研究中水分梯度不广泛, 或仅有几个梯度的对照分析, 没有能够准确理解净光合速率为零时的临界叶片水分阈值和叶片所能持有的最大水分含量, 制约着叶片光合作用对叶片水分变化响应的准确模拟。
以华北夏玉米为研究对象, 利用夏玉米三叶期6个水分梯度持续干旱的影响模拟试验资料, 分析夏玉米叶片水分变化及其与叶片净光合速率与土壤水分的关系, 揭示夏玉米叶片含水量随土壤水分变化的特点以及其水分变化对叶片净光合速率的影响特征, 探讨叶片水分是否存在光合作用的临界点及其最大持水量, 以增进叶片水分与光合作用和土壤水分关系的理解, 也为准确模拟光合作用提供依据。
1 材料与方法 1.1 试验材料本试验以夏玉米郑单958为供试材料, 于2014年6—10月在中国气象局固城生态与农业气象试验站(39°08′N, 115°40′E)开展。该区属于暖温带大陆性季风气候, 多年平均气温为12.2℃, 年平均降水量528 mm, 年平均日照时数2264 h[20]。试验设计为小区试验, 每个小区面积为8 m2(2 m×4 m), 各小区之间用3 m深混凝土墙隔离, 配有大型电动遮雨棚, 降雨时启用, 排除降雨影响, 其余时间开放, 保证玉米生长所需光照。土壤为沙壤土, 0—50 cm平均土壤容重为1.2 g/cm3, 平均田间持水量为22.1%, 全氮0.98 g/kg, 全磷1.02 g/kg, 全钾17.26 g/kg, pH值为8.19[21]。
1.2 试验设计2014年6月24日播种, 小区玉米行距50 cm, 株距25 cm, 每个小区64株, 播种后追施磷酸二铵300 kg/hm2, 每个小区240 g。试验共分6个处理, 每个处理3个重复, 共18个试验小区。播种前测定各小区土壤水分, 进行灌溉, 使各小区底墒一致, 0—50 cm的土壤相对湿度均为50%;播种至三叶期期间各小区均匀少量灌溉, 使根部水分适宜;出苗后(三叶期)于7月2日进行6个水平的梯度灌溉, 灌溉量由小到大分别按保定地区1981—2010年7月的30年平均降水的7%(W1)、20%(W2)、40%(W3)、60%(W4)、80%(W5)和100%(W6)进行一次性灌水处理(表 1), 之后不再灌溉, 各处理先后发生干旱, 以模拟不同强度与持续时间土壤水分胁迫对夏玉米的影响。
| 处理Treatments | W1 | W2 | W3 | W4 | W5 | W6 |
| 灌溉量Irrigation amount/mm | 10 | 30 | 60 | 90 | 120 | 150 |
| 占7月份降水量的百分比Percentage of precipitation in July /% | 7 | 20 | 40 | 60 | 80 | 100 |
| 1981—2010年7月的30年平均降水的7%(W1)、20%(W2)、40%(W3)、60%(W4)、80%(W5)和100%(W6) | ||||||
采用烘干称重法测定由土钻法得到的0—50 cm每10 cm一层的土壤分层重量含水量θv, 由土壤重量含水量与田间持水量的比值求出土壤相对湿度RSWC(%)。
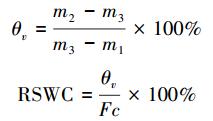
|
式中, θv为土壤重量含水量(g/g);m1为铝盒重(g), m2为铝盒和湿土总重(g), m3为铝盒和干土总重(g);Fc为田间持水量(%)。
1.3.2 叶片净光合速率利用美国LI-COR公司生产的Li-6400型便携式光合系统分析仪测定。研究表明土壤水分和光合有效辐射是影响夏玉米生理指标最主要的生态因子[22], 为控制光合有效辐射的影响, 进行各处理光响应曲线测定, 发现叶片净光合速率在光强为1500 μmol m-2 s-1时达到饱和。因此选择气候条件稳定的晴天, 气温在31—33℃左右, CO2浓度380 μl/L左右, 大气相对湿度为50%左右。采用荧光叶室, 设定光强为1500 μmol m-2 s-1, 流速为300 μmol/s, 于9:00—12:00进行观测。每个小区选取1株玉米标准植株测定其顶部第一片完全展开叶中上部位置的净光合速率Pn (μmol m-2 s-1), 以保证取样叶片对植株光合性能的代表性。
1.3.3 叶片含水量取样于上午光合生理生态参数观测后进行, 所取叶片样本与光合观测为同一叶片, 参照《农业气象观测规范》[23], 将植株叶片分离并标记, 称取鲜重, 再放入烘箱105℃杀青1 h, 在80℃下烘干至恒重, 并称取干重, 以求取叶片样本的含水量。
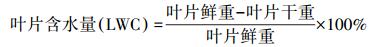
|
2014年7月2日进行最后1次水分灌溉处理, 为处理第1天, 选择晴朗天气, 对上述指标进行测定, 测定时间见表 2。
| 观测时间 Observation time |
生育期 Growth stages |
处理时间 Processing time/d |
| 7月11日 | 三叶期 | 10 |
| 7月18日 | 三叶期—七叶期 | 17 |
| 7月31日 | 七叶期—拔节期 | 30 |
| 8月7日 | 七叶期—拔节期 | 37 |
| 8月20日 | 拔节期—抽雄期 | 50 |
叶片含水量的饱和值采用米氏方程(Michaelis-Menten Equation)[24]得出。叶片含水量与土壤相对湿度的关系可表示如下:
y=ax/(b+x)
式中, y为叶片含水量, x为土壤相对湿度, a、b为常数。
一般采用Lineweaver-Burk双倒数作图法分析, 1/y=(b+x)/ax=b/a·1/x+1/a, 记Y=1/y, X=1/x, 则Y=b/a·X+1/a, 则变换为线性关系, 求取土壤相对湿度为100%时的叶片含水量。
1.6 数据处理试验数据采用Microsoft Excel 2013软件整理, 以平均值±标准误表示, 并用SigmaPlot 12.5软件进行曲线拟合、分析拟合方程显著性并绘图。
2 结果与分析 2.1 干旱发生发展过程及叶片水分的时间变化特征研究表明夏玉米85%左右的根部干重分布在40 cm以上[25]。为此, 本研究主要观测0—40 cm土层的土壤相对湿度。
灌水处理前(7月1日)土壤水分测定(图 1)结果表明, 灌水处理前各处理间无显著性差异, 7月2日梯度灌水后没有进行土壤水分测定, 于7月11日土壤水分测定表明, 各处理土壤相对湿度存在显著性差异, 达到梯度控水效果。随着时间推移, 由于土壤水分蒸发和玉米生长消耗, 所有处理的土壤相对湿度均显著下降, 且处理间的差异呈逐渐减小趋势。总体来看, 土壤相对湿度在处理前期下降幅度较大, 后下降平缓, 且W1—W6处理的土壤相对湿度平均每天下降速率为0.25%、0.46%、0.44%、0.77%、0.91%、1.02%, 随土壤水分胁迫强度的增加而减小。

|
| 图 1 土壤相对湿度和夏玉米叶片含水量的动态变化 Fig. 1 Dynamics of relative soil water content and leaf water content of summer maize W1,一次性灌溉量为10mm的处理;W2,一次性灌溉量为30mm的处理;W3,一次性灌溉量为60mm的处理;W4,一次性灌溉量为90mm的处理;W5,一次性灌溉量为120mm的处理;W6,一次性灌溉量为150mm的处理 |
夏玉米叶片含水量随着干旱的发生发展总体呈下降趋势(图 1)。在水分处理前期, 玉米对水分胁迫响应显著, 所有处理的叶片含水量下降速率较大, 高水分处理之间的叶片含水量无显著性差异。在水分处理后17—30 d, 不同处理的叶片含水量下降均有减缓趋势, 其中较多灌水处理W4—W6的土壤水分仍保持较大下降速率, 夏玉米逐渐受到水分胁迫, 在轻度水分胁迫下存在一定干旱适应[26-27], 叶片含水量下降减缓;而W1—W3处理干旱胁迫较重, 持续时间较长, 相比于高水分处理, 叶片含水量下降趋势明显, 此时不同处理间的差异显著。持续控水30 d后, 所有灌水处理的叶片含水量显著下降, 处理间差异减小。在持续控水50 d后, 土壤相对湿度变化缓慢, W2—W6处理的叶片含水量基本接近, 与W1处理存在显著差异。
2.2 叶片水分与土壤水分的关系图 2给出了在不同强度灌水处理下, 随着干旱发生发展以及同一观测日期不同处理间, 夏玉米叶片含水量与土壤相对湿度的关系。叶片含水量随着土壤相对湿度的下降而显著下降, 且不同水分胁迫强度与胁迫时间均影响叶片含水量的变化速率。低灌水处理W1叶片含水量随土壤相对湿度减小而下降的速率显著大于高灌水处理W6的下降速率, 即随着土壤水分胁迫强度的增加, 叶片含水量随土壤水分变化的速率逐渐增加, 说明土壤水分胁迫强度显著影响叶片水分的下降速率。同时, W4、W5和W6处理的叶片水分下降斜率比较相近, 与其他处理存在显著差异, 可以认为轻度水分胁迫对叶片含水量的变化影响较小, 只有达到一定干旱强度后, 叶片含水量的下降速率开始明显增加。

|
| 图 2 不同水分处理及不同观测时间叶片含水量与土壤相对湿度的关系 Fig. 2 The relationship between leaf water content and relative soil water content at different watering treatments and different observation times |
在7月11日观测中, W3—W6处理的叶片含水量之间没有显著性差异, 而与W1、W2处理存在显著差异, 说明在土壤水分充足时, 夏玉米叶片含水量相对稳定且保持在最大值, 当土壤含水量降低至一定程度时, 叶片含水量开始随土壤相对湿度减小显著下降。随着干旱持续时间的增加, 叶片含水量的绝对值逐渐减小, 且不同观测时间下, 叶片含水量随土壤相对湿度变化的斜率不同呈减小趋势, 即处理间的叶片含水量差异逐渐减小。
整个试验数据显示, 夏玉米叶片含水量随土壤相对湿度的增加先增加到最大值后保持不变, 两者之间呈非直角双曲线关系。此时的叶片含水量为叶片最大持水能力, 是植物生理能利用的叶片水分上限(图 3)。根据米氏方程及Lineweaver-Burk双倒数作图法分析, 得Y=0.01545X+0.0102, Y为叶片含水量的倒数, X为土壤相对湿度的倒数(图 3)。根据拟合方程, 当土壤相对湿度为100%, 即X=0.01时代入, 求得叶片含水量为85.14%, 即土壤水分达到田间持水量后, 叶片持水量为85.14%。

|
| 图 3 整个试验期间叶片含水量与土壤相对湿度的关系及Lineweaver-Burk双倒数作图法 Fig. 3 The relationship between leaf water content and relative soil water content and the double reciprocal Lineweaver-Burk plot throughout the trial period |
试验数据表明叶片含水量的变化对叶片净光合速率的影响显著。无论是随着干旱发生发展还是同一观测时间相同气象条件下, 夏玉米叶片净光合速率对叶片水分的响应主要呈二次曲线关系(图 4), 即随着叶片含水量的增加, 叶片净光合速率呈先增加后减少的抛物线关系, 并通过显著性检验(表 3、表 4)。不过高水分W6处理和7月11日观测可能由于叶片含水量已达到饱和, 低水分W1处理和8月20日观测由于干旱强度大, 干旱持续时间长, 叶片含水量梯度减小, 且叶片净光合速率受水分胁迫影响波动较大, 使得二次曲线关系没有达到显著或退化为线性方程也未通过显著性检验, 说明显著影响叶片净光合速率的叶片水分存在一定阈值范围。从不同水分处理的生育进程来看, W1处理的叶片净光合速率总体最低, 但并不是W6处理达到最大净光合速率, W3—W4处理的叶片净光合速率相对较高, 此时高灌水处理W6在生育期初期, 可能由于土壤水分过多阻碍根系呼吸, 间接影响玉米光合能力。当叶片含水量小于适宜范围时, 叶片净光合速率随着叶片含水量的下降而显著减小。

|
| 图 4 不同水分处理与不同观测时间的叶片净光合速率与叶片含水量的关系 Fig. 4 The relationship between Pn and LWC at different watering treatments and different observation times |
| 观测时间 Observation time |
拟合方程 Fitting equations |
R2 | Pn为零时的叶片含水量/% LWC when Pn reduces to zero |
| 7月11日 | y=-0.5494x2+ 92.051x-3826.3 | 0.8456 | 76.45 |
| 7月18日 | y=-1.2129x2+190.13x-7416.6 | 0.9944*** | 73.05 |
| 7月31日 | y=-0.3888x2+61.272x -2384.6 | 0.9424* | 70.10 |
| 8月7日 | y=-0.9183x2+137.03x-5093.9 | 0.9379* | 70.18 |
| 8月20日 | y=12.606x-899.63 | 0.3283 | 71.37 |
| ***, **, *分别表示所建立的回归方程通过了0.005、0.01、0.05水平的显著性检验 | |||
| 处理 Treatments |
拟合方程 Fittingequations |
R2 | Pn为零时的叶片含水量/% LWC when Pn reduces to zero |
| W1 | y=-0.3165x2+48.553x-1843.7 | 0.8752 | 69.12 |
| W2 | y=-0.4488x2+70.76x-2759 | 0.9756* | 70.66 |
| W3 | y=-0.4263x2+68.215x-2692.6 | 0.9491* | 70.76 |
| W4 | y=-0.3583x2 + 58.101x-2319.1 | 0.9621* | 71.02 |
| W5 | y=-0.4586x2+73.584x-2917.6 | 0.9898** | 71.58 |
| W6 | y=-0.2211x2+35.886x-1424.6 | 0.8050 | 69.19 |
| 全部处理All treatments | y=-0.2774x2+44.905x-1785.9 | 0.8034*** | 70.30 |
| ***, **, *分别表示所建立的回归方程通过了0.005、0.01、0.05水平的显著性检验 | |||
表 3、4的拟合方程计算7月11日至8月20日5次观测的叶片净光合速率为零的临界叶片含水量分别为76.45%、73.05%、70.10%、70.18%、71.37%, 随着土壤水分胁迫持续时间的增加临界叶片含水量呈先下降后上升的趋势, 即抗旱性随土壤水分胁迫持续时间的增加先增加后减小[16]。7月11日拟合方程得到的76.45%与其余临界叶片含水量存在显著差异, 可能由于此时处于三叶期, 叶片净光合速率显著小于其余生育期造成[28]。W1—W6灌水处理下的叶片净光合速率为零的临界叶片含水量分别为69.12%、70.66%、70.76%、71.02%、71.58%、69.19%, 除高水分处理W6外, 随着土壤水分胁迫强度的增加临界叶片含水量呈减小趋势, 可能是玉米对干旱胁迫有一定的适应性即光合驯化作用[29]。尽管土壤水分胁迫强度及持续时间不同, 得到的临界叶片含水量不同, 但比较相近, 在69.12%—73.05%范围内。平均而言, 当叶片含水量下降到70.30%时, 叶片净光合速率为零, 是判断叶片水分亏缺的重要参考。
3 讨论作物光合作用受到多种外界条件的影响[30], Foley等[31]对叶片尺度净光合速率的模型中给出了C3、C4植物叶片净光合速率对光照强度、胞间CO2浓度与叶片温度的响应模型, 而水分是影响叶片净光合速率的重要环境因子, 也是光合作用的原料之一。目前关于光合作用对水分的响应研究主要是基于土壤含水量。刘建栋等[32]建立了华北地区冬小麦叶片光合作用模型的土壤湿度订正函数。黄耀等[33]研究光合作用环境因子影响函数时给出基于最适土壤水分含量上下限、凋萎点土壤含水量给出水分响应函数。黄乾[34]基于最适土壤含水率构建了指数函数、二次函数和线性函数的水稻叶片水分修正光合模型。但是干旱是一个逐步发展的动态过程, 受持续时间、土壤水分传输等一系列因素影响, 静态的土壤含水量所包含的干旱的累积信息少[35]。土壤含水量相同并不能说明作物受旱程度一致, 在发生水分胁迫时, 由于植株不同器官对水分的敏感程度不同导致各器官的含水率不一致, 叶片作为光合器官, 其水分变化直接影响叶片净光合速率。因此, 本研究采用更能准确反映作物真实受旱程度的叶片含水量, 来探讨干旱胁迫下叶片水分变化及其对光合作用的影响规律。
干旱胁迫使玉米发生一系列生理变化, 首先叶片等器官的含水量发生响应, 然后叶片光合作用在叶片形态未发生变化时受到影响, 随着胁迫程度的加剧, 叶片形态颜色变化, 最终影响到生物量与产量。持续土壤水分胁迫使叶片含水量逐渐下降, 叶片净光合速率随叶片含水量先略有增加后减小为零, 此时的叶片含水量可以理解为叶片进行光合作用所需的最小叶片含水量。研究指出光合作用所需的水分只是植物所吸收的1%以下[1], 本研究也表明水分在植物体内的功能是多方面的。只有叶片水分含量高于维持细胞形态所需水分, 才能开始叶片光合作用, 叶片净光合速率才能大于零, 才有光合产物的形成供植物所需。同时, 任丽花等[36]研究土壤水分胁迫对圆叶决明叶片含水量研究中发现, 水分胁迫第10天的轻度、中度干旱处理的叶片含水量与对照相比无显著差异, 即在较高土壤水分时, 叶片含水量达到饱和值且受土壤水分变化影响小。王怀树等[37]研究土壤水分对小麦叶片含水量影响中得出叶片相对含水量指标与土壤含水量呈线性关系, 与本研究指出叶片含水量与土壤相对湿度呈非直角双曲线关系存在一定差异。可能由于土壤水分梯度小, 线性关系更显著, 并未得到叶片含水量的饱和值。本研究土壤水分梯度广, 得出夏玉米叶片含水量随土壤相对湿度的增加逐渐增加达到饱和叶片含水量后趋于不变。因此, 叶片存在净光合速率为零的叶片水分阈值和饱和叶片含水量, 对于叶片光合作用水分影响机制理解、作物耐旱性评价、农业灌溉指导以及进行植物生产力准确估算具有重要意义。
叶片净光合速率不仅受水分、温度等环境因子影响, 也受到叶片形态结构[38-39]、叶位[40]、叶龄[41-42]、所处生育期[43]等。本研究以夏玉米叶片为研究对象, 测定顶部第一片展开叶的中上部位, 因此叶片含水量阈值使用方面应注意其物种及叶片属性。本研究探究持续土壤水分胁迫对夏玉米叶片光合特性的影响, 存在干旱累积效应, 造成生育期的延后。在持续干旱后期, 即使均选取顶部第一片叶, 处理间仍存在生育期差异;同时, 1500 μmol m-2 s-1的光强设置在试验后期因持续干旱易产生光抑制[44], 均能影响叶片净光合速率的准确性。因此, 为进一步提高作物光合参数的准确性, 未来需要提高试验设计的合理性, 控制CO2浓度、温度等环境因子, 确定每次观测的饱和光强, 并开展不同生育期下叶片净光合速率对叶片含水量的响应研究, 进一步明确不同生育期的叶片水分阈值, 为光合作用准确模拟及定量遥感监测提供依据。
4 结论由于物种特性、叶片状况及环境因子不同, 不同物种的叶片净光合速率可利用的叶片水分及其叶片含水量阈值可能不一致, 但其对叶片水分响应的趋势是一致的, 这为开展其他作物干旱临界指标研究提供参考。根据夏玉米水分胁迫模拟试验, 分析叶片含水量变化与叶片净光合速率和土壤相对湿度的关系, 得到如下结论:
(1) 夏玉米叶片含水量与土壤相对湿度呈非直角双曲线关系, 土壤水分胁迫程度越重, 叶片含水量随土壤水分变化的速率越快。随土壤水分的增加, 叶片含水量先增加后趋于不变, 即叶片饱和含水量为85.14%。
(2) 本试验证实了叶片水分是影响夏玉米叶片净光合速率的重要因子, 且在一定范围内的叶片净光合速率对叶片水分的响应呈显著的二次曲线关系。高土壤水分条件下, 叶片含水量达到饱和;严重水分胁迫下叶片含水量梯度减小且叶片净光合速率波动较大, 二次曲线规律不显著。
(3) 夏玉米叶片净光合速率为零时的叶片含水量在不同条件下存在差异, 它随着土壤水分胁迫时间的增加呈先减小后增加趋势;随着土壤水分胁迫程度的加剧总体呈减小趋势。平均而言, 当叶片含水量减小到70.30%时, 叶片净光合速率为零, 是叶片光合生理活性的一个重要转折点。
致谢: 感谢石耀辉、麻雪艳、王秋玲、周怀林、王敏政、王帆等在试验观测和文章撰写过程中给予的帮助。| [1] | 潘瑞炽. 植物生理学(第六版). 北京: 高等教育出版社, 2008: 8–9. |
| [2] | 牛芬菊. 不同灌溉条件下土壤水分对小麦叶片含水量的影响. 科学种养, 2016(3): 185–185. |
| [3] | Colombo R, Meroni M, Marchesi A, Busetto L, Rossini M, Giardino C, Panigada C. Estimation of leaf and canopy water content in poplar plantations by means of hyperspectral indices and inverse modeling. Remote Sensing of Environment, 2008, 112(4): 1820–1834. DOI:10.1016/j.rse.2007.09.005 |
| [4] | 王娟, 郑国清. 夏玉米冠层反射光谱与植株水分状况的关系. 玉米科学, 2010, 18(5): 86-89, 95-95. |
| [5] | 陈小平, 王树东, 张立福, 姜海玲. 植被叶片含水量反演的精度及敏感性. 遥感信息, 2016, 31(1): 48–57. |
| [6] | Zhu X, Wang T J, Skidmore A K, Darvishzadeh R, Niemann K O, Liu J. Canopy leaf water content estimated using terrestrial LiDAR. Agricultural and Forest Meteorology, 2017, 232: 152–162. DOI:10.1016/j.agrformet.2016.08.016 |
| [7] | Tezara W, Mitchell V J, Driscoll S D, Lawlor D W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature, 1999, 401(6756): 914–917. DOI:10.1038/44842 |
| [8] | Chaves M M, Pereira J S, Maroco J, Rodrigues M L, Ricardo C P P, Osorio M L, Carvalho I, Faria T, Pinheiro C. How plants cope with water stress in the field? Photosynthesis and growth. Annals of botany, 2002, 89(7): 907–916. DOI:10.1093/aob/mcf105 |
| [9] | Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress:regulation mechanisms from whole plant to cell. Annals of botany, 2009, 103(4): 551–560. DOI:10.1093/aob/mcn125 |
| [10] | Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33(1): 317–345. DOI:10.1146/annurev.pp.33.060182.001533 |
| [11] | 关义新, 戴俊英, 林艳. 水分胁迫下植物叶片光合的气孔和非气孔限制. 植物生理学报, 1995, 31(4): 293–297. |
| [12] | Flexas J, Medrano H. Drought-inhibition of photosynthesis in C3 plants:stomatal and non-stomatal limitations revisited. Annals of Botany, 2002, 89(2): 183–189. DOI:10.1093/aob/mcf027 |
| [13] | Zhou S X, Duursma R A, Medlyn B E, Kelly J W G, Prentice I C. How should we model plant responses to drought? An analysis of stomatal and non-stomatal responses to water stress. Agricultural and Forest Meteorology, 2013, 182-183: 204–214. DOI:10.1016/j.agrformet.2013.05.009 |
| [14] | 张光灿, 贺康宁, 刘霞. 黄土高原半干旱区林木生长适宜土壤水分环境的研究. 水土保持学报, 2001, 15(4): 1–5. |
| [15] | 曾伟, 蒋延玲, 李峰, 周广胜. 蒙古栎(Quercus mongolica)光合参数对水分胁迫的响应机理. 生态学报, 2008, 28(6): 2504–2510. |
| [16] | 刘永宏, 段玉玺, 白育英. 大青山南麓阳坡油松生长的适宜土壤水分环境研究. 水土保持研究, 2013, 20(5): 69-72, 76-76. |
| [17] | 夏宣宣, 张淑勇, 张光灿, 方立东, 张松松, 李辉. 黄土丘陵区土壤水分对黄刺玫叶片光响应特征参数的影响. 生态学报, 2016, 36(16): 5142–5149. |
| [18] | 王纪华, 赵春江, 黄文江, 郭晓维, 李鸿祥. 土壤水分对小麦叶片含水量及生理功能的影响. 麦类作物学报, 2001, 21(4): 42–47. DOI:10.7606/j.issn.1009-1041.2001.04.099 |
| [19] | 张文丽. 玉米幼苗对不同土壤水分条件的生理反应[D]. 开封: 河南大学, 2004. http://cdmd.cnki.com.cn/Article/CDMD-10475-2004072492.htm |
| [20] | 谭凯炎, 郭建平. 固城生态与农业气象试验站定位观测与研究数据集. 北京: 气象出版社, 2015. |
| [21] | 任三学, 赵花荣, 姜朝阳, 谭凯炎. 土壤水分胁迫对冬小麦旗叶光合特性的影响. 气象科技, 2010, 38(1): 114–119. |
| [22] | 刘祖贵, 陈金平, 段爱旺, 孟兆江, 张寄阳, 刘战东. 夏玉米叶片生理特性与生态因子间关系的研究. 杂粮作物, 2006, 26(4): 288–292. |
| [23] | 国家气象局. 农业气象观测规范. 北京: 气象出版社, 1993: 28–29. |
| [24] | 王镜岩, 朱圣庚, 徐长法. 生物化学教程. 北京: 高等教育出版社, 2008: 152–154. |
| [25] | 赵秉强, 张福锁, 李增嘉, 李凤超, 史春余, 张骏, 张新春, 申加祥, 潘海军, 赵甲美. 套作夏玉米根系数量与活性的空间分布及变化规律. 植物营养与肥料学报, 2003, 9(1): 81–86. DOI:10.11674/zwyf.2003.0115 |
| [26] | 汤章城. 植物对水分胁迫的反应和适应性-Ⅱ植物对干旱的反应和适应性. 植物生理学报, 1983(4): 3–9. |
| [27] | 王智威. 水分胁迫下玉米苗期的生长及生理响应[D]. 杨凌: 西北农林科技大学, 2013. http://cdmd.cnki.com.cn/article/cdmd-10712-1013346225.htm |
| [28] | 童淑媛, 杜震宇, 徐洪文, 刘胜群, 朱先灿, 宋凤斌. 不同株型玉米叶片净光合速率差异研究. 东北农业大学学报, 2011, 42(4): 42–47. |
| [29] | 孙谷畴, 赵平, 曾小平. 两种木兰科植物叶片光合作用的光驯化. 生态学报, 2004, 24(6): 1111–1117. |
| [30] | 林金科, 赖明志, 詹梓金. 茶树叶片净光合速率对生态因子的响应. 生态学报, 2000, 20(3): 404–408. |
| [31] | Foley J A, Prentice I C, Ramankutty N, Levis S, Pollard D, Sitch S, Haxeltine A. An integrated biosphere model of land surface processes, terrestrial carbon balance, and vegetation dynamics. Global Biogeochemical Cycles, 1996, 10(4): 603–628. DOI:10.1029/96GB02692 |
| [32] | 刘建栋, 王馥棠, 于强, 毛飞, 毕建杰, 樊广华. 华北地区冬小麦叶片光合作用模型在农业干旱预测中的应用研究. 应用气象学报, 2003, 14(4): 469–478. |
| [33] | 黄耀, 杨兆芳, 于永强, 姜军, 孙文娟. 稻麦作物净初级生产力模型研究:模型的建立. 环境科学, 2005, 26(2): 11–15. |
| [34] | 黄乾. 控制灌溉条件下水稻光合特性试验研究[D]. 南京: 河海大学, 2005. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y716967 |
| [35] | 张玉书, 米娜, 陈鹏狮, 纪瑞鹏. 土壤水分胁迫对玉米生长发育的影响研究进展. 中国农学通报, 2012, 28(3): 1–7. DOI:10.11924/j.issn.1000-6850.2010-1968 |
| [36] | 任丽花, 王义祥, 翁伯琦, 方金梅, 应朝阳, 黄毅斌. 土壤水分胁迫对圆叶决明叶片含水量和光合特性的影响. 厦门大学学报:自然科学版, 2005, 44(S): 28–31. |
| [37] | 王怀树. 河西绿洲不同灌溉条件下土壤水分对小麦叶片含水量的影响. 安徽农业科学, 2014, 42(21): 6957–6959. DOI:10.3969/j.issn.0517-6611.2014.21.017 |
| [38] | 高照全, 冯社章, 李天红, 张显川. 苹果树冠不同部位叶片结构、内含物和模拟光合能力的比较. 中国生态农业学报, 2010, 18(6): 1245–1250. |
| [39] | 陈德兴, 王天铎. 叶片叶肉结构对环境光强的适应及对光合作用的影响. 应用生态学报, 1990, 1(2): 142–148. |
| [40] | 李伟, 眭晓蕾, 王绍辉, 关秋竹, 胡丽萍, 周明, 孟凡珍, 张振贤. 黄瓜幼苗不同叶位叶片光合特性对弱光的响应. 中国农业科学, 2008, 41(11): 3698–3707. DOI:10.3864/j.issn.0578-1752.2008.11.034 |
| [41] | Limousin J M, Misson L, Lavoir A V, Martin N K, Rambal S. Do photosynthetic limitations of evergreen Quercus ilex leaves change with long-term increased drought severity?. Plant Cell & Environment, 2010, 33(5): 863–875. |
| [42] | Peri P L, Arena M, Pastur G M, Lencinas M V. Photosynthetic response to different light intensities, water status and leaf age of two Berberis species (Berberidaceae) of Patagonian steppe, Argentina. Journal of Arid Environments, 2011, 75(11): 1218–1222. DOI:10.1016/j.jaridenv.2011.06.003 |
| [43] | 刘帆, 申双和, 李永秀, 李丽, 邹学智. 不同生育期水分胁迫对玉米光合特性的影响. 气象科学, 2013, 33(4): 378–383. DOI:10.3969/2012jms.0126 |
| [44] | 苏培玺, 严巧娣. C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征. 生态学报, 2006, 26(1): 75–82. |