 2018, Vol. 38
2018, Vol. 38文章信息
- 杨路存, 赵玉红, 徐文华, 周国英.
- YANG Lucun, ZHAO Yuhong, XU Wenhua, ZHOU Guoying.
- 青海省高寒灌丛物种多样性、生物量及其关系
- Species diversity, biomass, and their relationship in the alpine shrubberies of Qinghai Province
- 生态学报. 2018, 38(1): 309-315
- Acta Ecologica Sinica. 2018, 38(1): 309-315
- http://dx.doi.org/10.5846/stxb201609231921
-
文章历史
- 收稿日期: 2016-09-23
- 修订日期: 2017-09-21





2. 中国科学院藏药研究重点实验室, 西宁 810008;
3. 青海省青藏高原特色生物资源研究重点实验室, 西宁 810008;
4. 西藏大学农牧学院, 林芝 860000
2. Key Laboratory of Tibetan Medicine Research, Chinese Academy of Sciences, Xining 810008, China;
3. Qinghai Key Laboratory of Qinghai-Tibet Plateau Biological Resources, Xining 810008, China;
4. Agriculture and Animal Husbandry College, Tibet University, Linzhi 860000, China
物种多样性对生态系统功能的影响是当前生态学领域所研究的重大科学问题。生物量水平是生态系统功能的重要表现形式, 而植物群落的生物量则是生态系统生物量的基础[1]。因此研究植物群落物种多样性与生物量的关系, 对于阐明植物多样性对生态系统功能的作用具有重要意义[2]。许多研究表明, 在自然群落中, 植物种多样性与生物量密切相关, 不同的生态学家提出了不同的关系模式, 主要有:正相关, 负相关, 单峰关系, 不相关, U形相关[3-7]。Tilman[8]经过长期实验认为, 物种多样性对生物量的影响有两种可能机制:(1)取样效应, 即群落的物种多样性越高, 高竞争力的物种被选中成为群落优势物种的机率越高;(2)生态位互补效应, 对资源和环境要求不同的物种组合, 能更完全地捕获和利用自然资源。因此, 多物种组合比单一物种在单作条件下具有更高的生物量。
近年来, 物种丰富度与生物量的关系的研究成为国内学者关注的焦点[9-11], 然而他们的研究大多集中于草原和草甸生态系统。已有的研究表明, 灌丛作为青藏高原主要的植被类型之一, 是该地区相对稳定的生态系统类型, 目前对该区灌丛研究主要集中在植物分类[12-13]、生物量和群落特征[14-15]、植被与环境因子关系[16-17]等方面, 对灌丛物种多样性特征与其生物量相互关系的研究少见报道[18]。为此, 本文以青海省高寒灌丛为研究对象, 通过对灌丛物种多样性、生物量及其相互关系的研究, 以期揭示青海省高寒灌丛物种多样性与生物量的关系, 为研究青藏高原地区的物种多样性与生物量的关系提供参考依据。
1 研究地区与方法 1.1 研究区概况青海省位于青藏高原的东北部(31.65°—39.32°N, 89.58°—103.07°E), 东西长约1200 km, 南北宽约800 km, 青海省土地总面积7215.24×104 hm2。燕山运动奠定地形复杂多样, 高山、丘陵、河谷、盆地交错分布, 平均海拔3000 m以上, 属典型高原大陆性气候。年平均气温-3.7—6.0℃, 年日照2340—3550 h, 年降水量16.7—776.1 mm(多为400 mm以下), 年蒸发量1118.4—3536.2 mm。青海省地处青藏高原, 环境条件较差, 乔林分布不多, 而灌林(灌丛)则充分发育, 总面积达200万hm2, 且类型繁多, 在生态环境保护上功能显著。按热量带划分, 青海省天然灌丛可分为温性灌丛、寒温性灌丛和高寒灌丛。高寒区域是青海灌林的主要分布地带, 占有较大的范围和面积, 也集中了众多的类型, 常见的高寒灌丛主要有:百里香杜鹃(Rhododendron thymifolium)、头花杜鹃(R. capitatum)、山生柳(Salix oritrepha)、金露梅(Potentilla fruticose)、高山绣线菊(Spiraea alpina)、鬼箭锦鸡儿(Caragana jubata)、窄叶鲜卑花(Sibiraea laevigata)等[19]。
1.2 样地的设置及取样方法本研究于2011年7月至2014年7月先后在青海省境内选择具有代表性, 且发育程度基本一致的10个灌丛群落, 设置大小为20 m×20 m的样地。样地采用GPS定位, 测定海拔, 10个灌丛群落概况见表 1、图 1。每个样地设置3个大小为5 m×5 m的样方, 每个样方的间距约10 m。在对每个样方内的灌木层物种进行每木调查的同时, 选择1个1 m ×1 m的代表性区域, 设置收获样方, 对地上生物量进行收割, 对地下生物量进行挖掘, 并分种称取根、茎、叶等器官的鲜重。同时, 将称地过鲜重的根、茎、叶带回室内(分部分样品多于100 g取样100 g), 用烘干法在65℃条件下烘干至恒重, 并称取样品干质量。通过样品鲜重与样品干重的比值, 结合收获样方内各器官的鲜重, 计算不同灌木物种不同器官的单位面积生物量[20]。通过3个样方不同物种不同器官生物量平均值相加, 计算单位面积灌木层生物量。此外, 在每个灌丛样方四角做4个1 m×1 m的小样方, 对小样方内的草本进行详细调查, 记录种类、数量、平均高度和盖度。同时, 设置1个1 m × 1 m的草本收获样方, 采用与灌木相同的处理方法, 按优势种和非优势种分别获取草本地上部分、地下部分的鲜重, 并取大约100 g作为样品带回室内, 采用与测定灌木层生物量相同的方法获取草本层生物量。
| 丛群落 代号 Code | 灌丛群落类型 Community types | 样地数 Sample No. | 灌木伴生种 The associate species of shrub | 盖度/% Coverage |
| Ⅰ | 金露梅群落 P. fruticose community | 12 | 山生柳 S. oritrepha、高山绣线菊 S. alpine、百里香杜鹃 R. thymifolium、鬼箭锦鸡儿 C. jubata、沙棘 H. rhamnoides | 54.3—85.7 |
| Ⅱ | 百里香杜鹃+头花杜鹃群落 R. thymifolium + R. capitatum community | 4 | 山生柳 S. oritrepha、高山绣线菊 S. alpina、鬼箭锦鸡儿 C. jubata、金露梅 P. fruticosa、刚毛忍冬 L. hispida、瑞香 D. odora | 70.5—95.0 |
| Ⅲ | 头花杜鹃群落 R. capitatum community | 3 | 山生柳 S. oritrepha、金露梅 P. fruticosa、高山绣线菊 S. alpina、鬼箭锦鸡儿 C. jubata | 70.5—72.4 |
| Ⅳ | 百里香杜鹃群落 R. thymifolium community | 4 | 山生柳 S. oritrepha、高山绣线菊 S. alpina、金露梅 P. fruticosa、华西银露梅 P. glabra var. mandshurica、刚毛忍冬 L. hispida、鬼箭锦鸡儿 C. jubata、鲜卑花 S. laevigata、烈香杜鹃 R. anthopogonoides | 63.5—81.0 |
| Ⅴ | 细枝绣线菊群落 S. myrtilloides community | 3 | 刚毛忍冬 L. hispida、华西银露梅 P. glabra var. mandshurica、小檗 B. thunbergii、金露梅 P. fruticosa | 33.1—75.0 |
| Ⅵ | 鲜卑花群落 S. laevigata community | 3 | 山生柳 S. oritrepha、金露梅 P. fruticosa、高山绣线菊 S. alpina、百里香杜鹃 R. thymifolium、鬼箭锦鸡儿 C. jubata、刚毛忍冬 L. hispida | 48.3—72.9 |
| Ⅶ | 山生柳群落 S. oritrepha community | 6 | 花楸 S. pohuashanensis、高山绣线菊 S. alpina、华西银露梅 P. glabra var. mandshurica、百里香杜鹃 R. thymifolium、鲜卑花 S. laevigata、冰川茶蔍 R. glaciale、金露梅 P. fruticosa、鬼箭锦鸡儿 C. jubata、刚毛忍冬 L. hispida | 51.8—79.5 |
| Ⅷ | 山生柳+鬼箭锦鸡儿群落 S. oritrepha+ C. jubata community | 3 | 金露梅 P. fruticosa | 78.6—95.0 |
| Ⅸ | 小叶金露梅群落 P. parvifolia community | 3 | 33.7—45.0 | |
| Ⅹ | 鬼箭锦鸡儿群落 C. jubata community | 3 | 金露梅 P. fruticosa | 72.6—85.7 |

① 重要值计算
公式为:灌木重要值Ⅳ=(相对密度+相对频度+相对盖度+相对高度)/4;草本重要值Ⅳ=(相对频度+相对盖度+相对高度)/3[21]。
② 群落物种多样性的测定以重要值作为多样性指数的测度依据, 选用以下几种常用测度方法[22]:
丰富度指数R=样地内所有物种数目;
Shannon-Wiener指数:
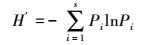
|
Simpson指数:
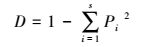
|
Pielou均匀度指数
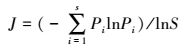
|
式中, Pi为样方内种i的重要值。
1.4 数据处理数据统计采用Excel软件, 相关性分析采用SPSS(11. 0版)软件。
2 结果 2.1 灌丛生物多样性在10个灌丛群落中, 共调查到植物种类207种, 分属43科130属。其中, 灌木18种, 以蔷薇科、杜鹃花科为主;草本189种, 以菊科、龙胆科、毛茛科和莎草科为主。灌丛总物种丰富度最高的为灌丛Ⅵ, 达42种, 最低的为灌丛Ⅱ, 仅有23种(表 2)。灌木物种丰富度最高和最低的分别为:灌丛Ⅱ(6)、Ⅲ(6)和Ⅸ(1), 而草本物种丰富度最高和最低的分别为:Ⅴ(37)、Ⅵ(37)和Ⅱ(17)。10个灌丛群落中, 草本物种数较灌木丰富。
| 代码 Code |
群落多样性Community diversity index | 灌木多样性Shrub diversity index | 草本多样性Herb diversity index | |||||||||
| R | H | D | J | R | H | D | J | R | H | D | J | |
| Ⅰ | 38 | 3.012 | 0.915 | 0.837 | 3 | 0.652 | 0.370 | 0.588 | 35 | 3.105 | 0.928 | 0.881 |
| Ⅱ | 23 | 2.448 | 0.870 | 0.783 | 6 | 1.578 | 0.738 | 0.855 | 17 | 2.260 | 0.902 | 0.871 |
| Ⅲ | 36 | 2.981 | 0.908 | 0.838 | 6 | 1.340 | 0.627 | 0.748 | 30 | 2.982 | 0.918 | 0.877 |
| Ⅳ | 34 | 2.763 | 0.875 | 0.784 | 5 | 1.324 | 0.642 | 0.788 | 29 | 2.810 | 0.902 | 0.871 |
| Ⅴ | 41 | 3.008 | 0.894 | 0.818 | 4 | 0.718 | 0.388 | 0.660 | 37 | 3.322 | 0.951 | 0.923 |
| Ⅵ | 42 | 3.006 | 0.907 | 0.803 | 5 | 1.110 | 0.579 | 0.741 | 37 | 3.154 | 0.930 | 0.872 |
| Ⅶ | 29 | 2.609 | 0.866 | 0.782 | 4 | 1.106 | 0.576 | 0.798 | 25 | 2.653 | 0.883 | 0.843 |
| Ⅷ | 26 | 2.543 | 0.871 | 0.781 | 3 | 0.9111 | 0.568 | 0.829 | 23 | 2.754 | 0.913 | 0.878 |
| Ⅸ | 31 | 3.039 | 0.928 | 0.885 | 1 | 0.517 | 0.335 | 0.747 | 30 | 3.087 | 0.932 | 0.908 |
| Ⅹ | 30 | 2.974 | 0.915 | 0.874 | 2 | 0.577 | 0.388 | 0.832 | 28 | 3.143 | 0.950 | 0.943 |
| R, 丰富度指数species richness;H, Shannon-Wiener指数Shannon-Wiener index;D, Simpson指数Simpson index;J, Pielou均匀度指数Pielou evenness index | ||||||||||||
从表 2可以看出, 灌丛群落Shannon-Wiener指数最高的是灌丛Ⅸ (3.039), 最低为灌丛Ⅱ(2.448)。灌木Shannon-Wiener指数最高, 为灌丛Ⅱ(1.578), 最低为灌丛Ⅸ(0.517)。草本Shannon-Wiener指数最高的为灌丛Ⅴ (3.322), 最低的为灌丛Ⅱ (2.260);Simpson指数在群落、灌木与草本中, 分别以灌丛Ⅸ、Ⅱ和Ⅴ最高, 其值为0.928、0.738和0.951, 以灌丛Ⅷ、Ⅸ和Ⅶ最低, 分别为0.866、0.335和0.883;灌丛群落均匀度指数介于0.781—0.874, 灌木均匀度指数介于0.588—0.855之间, 而草本介于0.871—0.943之间。
2.2 灌丛生物量青海省高寒灌丛不同灌丛群落生物量介于1893.03—7585.41 g/hm2之间, 平均值为3775.9 g/m2(图 2)。在灌丛各层中, 灌木生物量介于510.55—6740.07 g/m2之间, 平均值为2777.16 g/m2, 草本生物量介于369.06—1829.48 g/m2之间, 平均值为998.74 g/m2, 分别占灌丛总生物量平均值的73.55%和26.45%。在各灌丛类型中, 以山生柳+锦鸡儿灌丛生物量最高, 达7585.41 g/m2, 而小叶金露梅灌丛最低, 为1893.03 g/m2。

|
| 图 2 灌丛类型生物量特征 Fig. 2 Characteristic of the biomass of shrubberies |
物种多样性指数与生物量的相关分析表明(表 3), 不同灌丛类型物种丰富度与其总生物量负相关;草本物种丰富度与总生物量负相关;灌木物种丰富度与总生物量相关关系不显著。这说明灌丛总生物量受草本物种多样性影响较大, 而受灌木物种多样性影响较小, 总体表现为灌丛总生物量随物种多样性的增大而减小。
| 生物量 Biomass |
物种丰富度 Species richness |
Shannon-Wiener指数 Shannon-Wiener index |
Simpson指数 Simpson index |
Pielou均匀度指数 Pielou evenness index | ||||||||
| 合计 Total |
灌木 Shrub |
草本 Herb |
群落 Community |
灌木 Shrub |
草本 Herb |
群落 Community |
灌木 Shrub |
草本 Herb |
群落 Community |
灌木 Shrub |
草本 Herb | |
| 灌木生物量Shrub biomass | -0.085 | 0.109 | -0.074 | -0.081 | 0.162 | 0.023 | -0.142 | 0.233 | 0.107 | -0.108 | 0.318* | 0.189 |
| 草本生物量Herb biomass | -0.325* | 0.082 | -0.337* | -0.321* | 0.073 | -0.346* | -0.226 | 0.055 | -0.274 | -0.191 | -0.030 | -0.299 |
| 总生物量Total biomass | -0.331* | 0.149 | -0.333* | -0.324* | 0.182 | -0.266 | -0.292 | 0.220 | -0.145 | -0.238 | 0.213 | -0.104 |
在灌丛不同层次中, 灌木Pielou均匀度指数与灌木生物量线性相关, 相关系数为0.381, 相关性显著(P < 0.05), 表明灌木生物量随灌丛均匀度的增加而增大。而草本物种丰富度、总物种丰富度、群落Shannon-Wiener指数、草本Shannon-Wiener指数与草本生物量相关系数分别为-0.325、-0.337、-0.321和-0.346, 多样性指数与其生物量显著负相关(P < 0.05), 这表明草本生物量随多样性指数的增大而减小。
3 讨论生物多样性是生物之间以及与生存环境之间复杂的相互关系体现, 也是生物资源丰富度的标志[23]。本次样方调查的灌木植被群落中出现了207种植物, 隶属于43科130属, 与青海省野生种子植物共计94科, 557属, 2497种[24]相比种类简单。这种较低的物种多样性反映了青海省高寒地区脆弱的生态系统。
高寒灌丛是由耐寒的中生或旱生灌木为优势种而形成的植被, 是青藏高原植被类型的重要组成部分。而生物量是群落结构和功能的主要测度指标之一, 它体现了群落结构、环境以及人类活动等因素的综合作用, 反映了群落的结构特征及生长状况[15]。研究表明:青海省高寒灌丛不同灌丛类型生物量介于1893.03—7585.41 g/hm2之间。在各灌丛类型中, 以山生柳+锦鸡儿灌丛生物量最高, 达7585.41 g/m2, 而小叶金露梅灌丛最低, 为1893.03 g/m2。
这种差异可能源于组成不同灌丛的灌木种类个体形态差异较大。个体形态通过影响群落结构影响单位面积生物量的大小。此外, 青海省高寒灌丛群落的平均值为3775.9 g/m2, 略低于高巧等[25]研究四川省甘孜14个高寒灌丛群落平均总生物量(6410 g/m2), 这可能是由以下原因造成的:①本研究的高寒灌丛群落包括灌木和草本, 而高巧等的高寒灌丛群落包括灌木、草本和凋落物;②可能与灌丛所处的立地条件有关。
在自然生态系统中, 物种多样性与生物量的关系主要表现为5种形式, 即正相关、负相关、单峰函数关系、U型相关和不相关[3-7]。然而, 由于研究对象所选时间及所处空间的差异, 对这5种关系格局的看法和解释并未形成统一定论。在高寒地区植物群落物种多样性与生物量关系研究中, 覃光莲等[26], 杜国祯等[27]的研究呈对数线性增加关系, 武彦朋等[28]的研究表明物种丰富度与生物量的关系在小尺度上(群落内和群落间)表现为线性正相关、负相关, 在大尺度上(区域)则以单峰相关为主, 而杨元合等[9]的结论呈显著正相关。王勇军等[18]对岷江干旱河谷灌丛的研究显示灌丛总生物量与物种多样性呈显著正相关。本研究表明, 灌丛总生物量与总物种丰富度、草本物种丰富度以及群落的Shannon-Wiener指数呈显著负相关。这与王勇军等[18]对干旱河谷灌丛的研究结果相反, 主要的原因在于:受青藏高原特殊地理环境的影响, 青海省现存的高寒灌丛是与高寒草甸作斗争的产物, 是长期适应环
境恶化的“胜利者”[19]。它们在这个过程中, 演化生成了一些特有的抗性, 如多为单优结构, 优势种或建群种明显, 以防止种间的更替性退化。因此, 群落的多样性主要取决于草本的多样性。当灌木的盖度较小、生物量较低时, 下层的草本植物能充分利用光能资源和地表温度较高的小生境条件而充分发育, 群落的多样性较高, 因此灌丛总生物量与多样性呈现负相关关系。此外, 过去的研究表明, 野外观测到的物种丰富度与生物量的关系常常表现为驼峰型[5, 29], 并且认为物种之间的竞争作用对产生驼峰型关系起着重要作用[30-32]:随着生物量的增加, 群落中物种间的竞争作用随之增加;但当生物量增加到一定程度时, 群落中的一些物种消失, 从而使得丰富度下降[33]。本研究结果与野外常常观测到的驼峰型关系并不一致:生物量增加时, 物种丰富度随之减小, 并没有出现驼峰型关系中的上升部分, 这与杨元合等[34]对高寒草原和高寒草甸的研究结果相似。一种可能的解释是, 为在青藏高原这一特殊生境条件下, 物种间的共生作用更加明显, 或者说是物种之间的竞争作用不够强烈, 从而使得生物量高的群落保持较高的物种丰富度。另一种可能是这种关系也可能与我们所调查的范围不足有关。
植物物种多样性与生物量的关系, 以及植物与地形地貌、土壤、气候因子、人类活动相互间的作用机理极其复杂, 还需对高寒灌丛植被开展深入研究, 以进一步揭示灌丛物种多样性与生物量的关系, 从而更好地服务于区域植被恢复实践。
| [1] | 陈灵芝, 钱迎倩. 生物多样性科学前沿. 生态学报, 1997, 17(6): 565–572. |
| [2] | Loreau M. Biodiversity and ecosystem functioning:recent theoretical advances. Oikos, 2000, 91(1): 3–17. DOI:10.1034/j.1600-0706.2000.910101.x |
| [3] | Whittaker R H. Forest dimensions and production in the great Smoky mountains. Ecology, 1966, 47(1): 103–121. DOI:10.2307/1935749 |
| [4] | Redmann R E. Production ecology of grassland plant communities in western North Dakota. Ecological Monographs, 1975, 45(1): 83–106. DOI:10.2307/1942332 |
| [5] | Zobel K, Liira J. A scale-independent approach to the richness vs biomass relationship in ground-layer plant communities. Oikos, 1997, 80(2): 325–332. DOI:10.2307/3546600 |
| [6] | Wheeler B D, Shaw S C. Above-ground crop mass and species richness of the principal types of herbaceous rich-fen vegetation of lowland England and Wales. Journal of Ecology, 1991, 79(2): 285–301. DOI:10.2307/2260713 |
| [7] | McNaughton S J. Serengeti grassland ecology:the role of composite environmental factors and contingency in community organization. Ecological Monographs, 1983, 53(3): 291–320. DOI:10.2307/1942533 |
| [8] | Tilman D. Diversity and production in European grasslands. Science, 1999, 286(5442): 1099–1100. DOI:10.1126/science.286.5442.1099 |
| [9] | 安尼瓦尔·买买提, 杨元合, 郭兆迪, 方精云, 潘伯荣, 胡玉昆. 新疆巴音布鲁克高山草地物种丰富度与生产力的关系. 干旱区研究, 2006, 23(2): 289–294. |
| [10] | 杨利民, 周广胜, 李建东. 松嫩平原草地群落物种多样性与生产力关系的研究. 植物生态学报, 2002, 26(5): 589–593. |
| [11] | 尚占环, 姚爱兴, 龙瑞军. 干旱区山地植物群落物种多样性与生产力关系分析. 干旱区研究, 2005, 22(1): 74–78. |
| [12] | 关文彬, 冶民生, 马克明, 刘国华, 汪西林. 岷江干旱河谷植被分类及其主要类型. 山地学报, 2004, 22(6): 679–686. |
| [13] | 杨钦周. 岷江上游干旱河谷灌丛研究. 山地学报, 2007, 25(1): 1–32. |
| [14] | 冶民生, 关文彬, 谭辉, 马克明, 刘国华, 汪西林. 岷江干旱河谷灌丛α多样性分析. 生态学报, 2004, 24(6): 1123–1130. |
| [15] | 刘国华, 马克明, 傅伯杰, 关文彬, 康永祥, 周建云, 刘世梁. 岷江干旱河谷主要灌丛类型地上生物量研究. 生态学报, 2003, 23(9): 1757–1764. |
| [16] | 冶民生, 关文彬, 白占雄, 马克明, 刘国华, 汪西林. 岷江干旱河谷植物群落生态梯度分析. 中国水土保持科学, 2005, 3(2): 70–75. |
| [17] | 冶民生, 关文彬, 吴斌, 马克明, 刘国华, 汪西林, 陈箐妍. 岷江干旱河谷植物群落的复杂性. 生态学报, 2006, 26(10): 3159–3165. |
| [18] | 王勇军, 黄从德, 张健, 杨万勤, 王宪帅. 岷江干旱河谷灌丛物种多样性、生物量及其关系. 干旱区研究, 2010, 27(4): 567–572. |
| [19] | 魏振铎. 对青海天然灌林(灌丛)热量带和类型划分的重新认识. 青海环境, 2006, 16(2): 47–50. |
| [20] | 郝文芳, 陈存根, 梁宗锁, 马丽. 植被生物量的研究进展. 西北农林科技大学学报:自然科学版, 2008, 36(2): 175–182. |
| [21] | 熊莉军, 郭柯, 赵常明, 刘长成. 四川大巴山巴山水青冈群落的物种多样性特征. 生物多样性, 2007, 15(4): 400–407. |
| [22] | 胡正华, 于明坚, 徐学红, 付海龙. 浙江古田山自然保护区甜槠群落特征研究. 生态学杂志, 2004, 23(2): 15–18. |
| [23] | 牛丽丽, 张学培, 曹奇光. 我国西北干旱区生物多样性研究. 水土保持研究, 2007, 14(1): 223–225. |
| [24] | 陈桂琛, 彭敏. 青海湖地区植被及其分布规律. 植物生态学与地植物学学报, 1993, 17(1): 71–81. |
| [25] | 高巧, 阳小成, 尹春英, 刘庆. 四川省甘孜藏族自治州高寒矮灌丛生物量分配及其碳密度的估算. 植物生态学报, 2014, 38(4): 355–365. |
| [26] | 覃光莲, 杜国祯, 李自珍, 杨广运, 马建云, 娘毛加. 高寒草甸植物群落中物种多样性与生产力关系研究. 植物生态学报, 2002, 26(S1): 57–62. |
| [27] | 杜国祯, 覃光莲, 李自珍, 刘正恒, 董高生. 高寒草甸植物群落中物种丰富度与生产力的关系研究. 植物生态学报, 2003, 27(1): 125–132. |
| [28] | 武彦朋, 陈克龙, 张斐, 曹广超, 刘志杰, 苏茂新. 青海湖流域典型草地物种丰富度与生产力的关系. 生态学杂志, 2011, 30(7): 1449–1453. |
| [29] | Guo Q F, Berry W L. Species richness and biomass:dissection of the hump-shaped relationships. Ecology, 1998, 79(7): 2555–2559. DOI:10.1890/0012-9658(1998)079[2555:SRABDO]2.0.CO;2 |
| [30] | Newman E I. Competition and diversity in herbaceous vegetation. Nature, 1973, 244(5414): 310–310. DOI:10.1038/244310a0 |
| [31] | Tilman D. Resource Competition and Community Structure. New Jersey: Princeton University Press, 1982. |
| [32] | Huston M, Smith T. Plant succession:life history and competition. The American Naturalist, 1987, 130(2): 168–198. DOI:10.1086/284704 |
| [33] | Bonser S P, Reader R J. Plant competition and herbivory in relation to vegetation biomass. Ecology, 1995, 76(7): 2176–2183. DOI:10.2307/1941691 |
| [34] | 杨元合, 饶胜, 胡会峰, 陈安平, 吉成均, 朱彪, 左闻韵, 李轩然, 沈海花, 王志恒, 唐艳鸿, 方精云. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系. 生物多样性, 2004, 12(1): 200–205. |