 2017, Vol. 37
2017, Vol. 37文章信息
- 冯秋园, 万祎, 刘学勤, 刘永
- FENG Qiuyuan, WAN Yi, LIU Xueqin, LIU Yong.
- 持久性有机污染物在水生食物网中的传递行为
- Environmental behavior of persistent organic pollutants in aquatic food web
- 生态学报. 2017, 37(9): 2845-2857
- Acta Ecologica Sinica. 2017, 37(9): 2845-2857
- http://dx.doi.org/10.5846/stxb201602040256
-
文章历史
- 收稿日期: 2016-02-04
- 网络出版日期: 2016-12-19

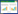




2. 北京大学城市与环境学院, 地球表面过程教育部重点实验室, 北京 100871;
3. 中国科学院水生生物研究所, 武汉 430072
2. College of Urban and Environmental Science, MOE Laboratory for Earth Surface Process, Peking University, Beijing 100871, China;
3. Institute of Hydrobiology Chinese Academy of Sciences, Wuhan 430072, China
持久性有机污染物 (POPs) 具有持久性、生物蓄积性、高毒性、半挥发性和远距离传输的特点, 会对人类健康和生态安全产生严重危害。POPs通过大气干湿沉降、地面径流和水-气界面直接交换进入水生生态系统, 是POPs在生态圈中传递的一个重要的“汇”。POPs进入水体后, 被生物体通过被动扩散和摄食作用吸收, 并沿食物网传递, 成为POPs在水生生态系统中非常重要的环境行为, 同时使顶级消费者受到高水平的POPs暴露风险, 因此研究POPs在水生生态系统食物网中的环境行为成为国内外研究的重点和热点[1-3], 但当前研究的重点仍集中在POPs在浮游食物网的传递, 对在底栖及底栖-浮游耦合食物网和微食物网的研究仍不够充分[4-5], 对各种内、外源环境压力下, 食物网变化对POPs环境行为影响的研究也相对缺乏[6-9]。鉴于此, 本文从食物网的主要特征与POPs环境行为的关系, POPs在底栖及底栖-浮游耦合食物网和微食物网中的环境行为及影响因素, 以及目前较为关注的几种内外源压力下食物网变化对POPs环境行为的影响等4个方面, 阐述了POPs在水生食物网中的环境行为, 为揭示POPs在水生生态系统中的归趋机制, 判断水生生物的POPs暴露风险, 制定科学的环境标准提供参考。
1 食物网主要特征与POPs环境行为的关系生物体所处的营养级和食物链长度是食物网的主要特征, 是研究POPs在食物网中生物富集或生物放大时的重点考虑因素。在不同的生态系统中, 其影响往往是不同的, 了解特定生态系统中营养级和食物链长度与POPs生物富集或生物放大之间的相关关系, 并探索影响这种相关关系的主要因素一直以来是生态学的热点问题。
1.1 营养级与POPs生物富集性的关系生物放大作用是营养级对POPs生物富集性影响的直接体现。一般情况下, 营养级越高, POPs生物放大效应越明显。研究发现, 除PFOA外, 爬行动物、哺乳动物等高营养级水生动物对PFCs (PFNA、PFDA、PFUA、PFDOA和PFOS) 的生物富集 (BAFs) 系数大于5000, 显著高于无脊椎动物和鱼类, 存在明显的生物放大现象[10-12]。诸多研究发现[13-14]生物营养级 (15N值) 和PCBs浓度之间显著正相关, 生物放大因子大于1。通常认为POPs生物富集性随营养级升高而增大的一个重要原因是脂质含量增加, 但实际上并不是所有食物网中生物体的脂质含量都随15N的升高而增加。另有研究表明, 单位脂质POPs含量与营养级之间仍有非常显著的正相关关系, 说明营养级增加本身对POPs生物富集也具有重要影响, 但营养级和脂质含量的相对贡献大小较难评价[15-16]。
但污染物并不总是随营养级的升高而显示生物放大效应。Neff等[17]、Varanasi等[18]和Broman等[19]在波罗的海的食物链中没有发现PAHs的生物放大现象, 推测是由于营养级越高, 生物体对PAHs的分解活性越高, 与其它地区的类似研究结果一致[20-22]。Mizukawa等[23]通过污染物浓度与生物体15N之间的相关关系, 分析了日本东京湾中20种PBEDs和PCBs的生物放大现象, 发现PCBs和部分PBDEs存在生物放大现象, 另几种PBDE同系物的浓度则随营养级升高而下降, 其原因可能是在生物体内被转化或代谢分解了。Nfon等[24]分析了具有不同理化性质的POPs在波罗的海浮游和底栖食物网中的传递, 发现所有PCBs同系物和OCPs存在显著的食物链放大作用 (FCMFs>1), 但由于生物体的代谢分解作用, PAHs和PCNs的食物链放大系数 (FCMFs) 小于1, 是营养稀释作用。上述研究说明, POPs的食物链放大作用受到生物转化和代谢分解的限制, 特别是在较高营养级中[25-26]。另有研究发现, 一般低营养级浮游食物网不存在污染物的生物放大或很低, 在高营养级消费者体内存在明显的生物放大, 因为低营养级生物对能量的需求量较低, 因此食物的摄入量相对更低, 而食物摄入是POPs富集的一个重要途径[27-29]。Lundgren等[30]发现在亚北极地区波的尼亚湾的底栖食物网中大多数高氯代PCNs没有生物放大现象, 推测其原因是由于高氯代PCNs体积较大, 阻碍了其在生物体内的扩散和吸收。Evenest研究了几种POPs在斯的巴特尔群岛的孔斯峡湾底栖食物网中的生物富集情况, 发现除了顺式九氯, 没有观测到POPs的生物放大现象, 几种无脊椎动物体内的POPs浓度等于甚至超过了鱼类或绒鸭体内的POPs浓度, 原因是这些底栖无脊椎动物是食腐性的, POPs的生物转化和排出能力低, 且直接暴露在POPs浓度较高的沉积物间隙水中[31]。
营养级对POPs生物富集性的影响通常是通过生物放大或营养级放大来分析, 一般情况下, POPs的生物富集性随营养级的升高而增强, 但也有例外, 因为其它各种生态和生理性质, 如猎物的丰度、生长速率、生物与沉积物的接触程度、摄食POM的量、微生物环导致的高15N和低POPs浓度等都不同程度地控制着POPs的生物富集, 使POPs的实际浓度偏离了根据营养级预测得到的值[31-33]。另外, 目前生物所处营养级一般用15N来表示, 生物放大性通过15N与POPs浓度的相关关系来分析, 但是对于底栖食物网, 单纯地利用15N有时并不能很好地指示底栖动物所处的实际营养级, 因为某些生物或所摄食的生物具有同位素分馏作用, 例如, 一些摄食POM的生物, 由于细菌环 (bacteria loop) 的分解和同位素分馏作用, POPs暴露风险降低, 但是15N的值却提高了[34];其次, 15N基准值的季节性变化也会影响15N和POPs相关关系的斜率;再者, 很多底栖无脊椎动物是杂食性/食腐性, 因此导致POPs传递的再循环, 使POPs的负荷较高, 但是单纯地分析15N可能并不能反映出这种循环。
目前, 有关POPs在水生食物网中的生物放大作用具有很大的争议, 如Bruner等[35]、Losser等[36]和Gobas等[37]等认为生物放大在整个食物网都存在, 但Borga等[38]认为在低营养级中, 由于生物体摄食吸收的POPs可以忽略, 所以不存在生物放大作用。Guruge等[39]认为生物放大只存在于直接呼吸空气的生物体内。当前, 对营养级升高造成的POPs生物富集性增强, 主要还是从脂质含量增加来解释[40], 但实际上, 营养级升高本身也是一个重要的影响因素, 但是其具体影响机制纷繁复杂, 存在很多疑问和不确定性。Gobas等[31]推测一是因为大部分生物量在沿营养级传递过程中转化成了能量被耗散, 造成污染物的浓缩, 二是由于食物在胃肠道中被消化, 提高了化学物质的逸度。综上发现, 营养级与POPs在食物网中生物富集性之间的关系, 及潜在的具体机制仍存在很多疑问。在具体的研究中, 需要根据生物体的生理特征, 结合POPs理化性质和生态系统的特点综合考虑, 才能得到较为准确的预测结果。
1.2 食物链长度与POPs生物放大效应的关系目前研究认为, 食物链延长会增加顶级消费者体内的污染物浓度[2, 41-42]。Kidd等[43]发现加拿大拉柏吉湖中, 鱼体内的毒杀酚浓度异常高, 明显高于其它湖泊的同种鱼类。通过对比湖泊的污染输入、营养级结构等发现, 拉柏吉湖比其它湖泊的食物链更长, 同种鱼在拉柏吉湖中的N15要显著高于其他湖泊, 从而导致其体内有异常高的污染物浓度, 由此揭开了食物链长度会提高POPs生物放大效应的认识。Whittle等[44]通过研究苏必利尔湖、安大略湖、休伦湖、伊利湖的食物链长度和生物放大系数发现食物链长度和生物放大系数呈正相关, 进一步证明食物链延长会提高POPs的生物放大作用。另有诸多研究发现食物链延长会导致生物体脂质含量增加, 那POPs生物富集的提高是否只是由于食物链延长引起的脂质含量增加而导致的?根据是否存在糠虾和草食性鱼类, Rasmussen等[2]对北美五大湖区的81个湖泊划分为3类, 通过对比顶级消费者湖红点鲑 (Salvelinus namaycush) 体内的PCBs浓度发现, 增加1个营养级, 湖红点鲑 (Salvelinus namaycush) 体内的PCBs浓度增加3.5倍, 但脂质含量只增加了1.5倍, 因此认为食物链延长本身也会导致POPs在顶级消费者体内生物富集性的提高。Bentzen等[40]进一步分析其原因得到:① 糠虾具有昼夜迁移行为, 会摄食部分沉积物, 导致其体内的POPs浓度比其它同营养级浮游生物更高;② 捕食浮游动物的鱼类通过捕食降低了浮游动物的生物量, 降低了生物量稀释作用, 提高了POPs浓度;③ 顶级消费者由捕食浮游动物改为捕食体型较大且营养级相对较高的糠虾, 摄食能耗降低, 脂质含量增加[45], POPs浓度升高。
2 底栖及底栖-浮游耦合食物网中POPs的环境行为目前, 有关POPs在食物网中传递的研究多集中在浮游食物网, 但底栖食物网是生态系统中物质循环和能量流动的重要环节, 并通过与浮游食物网相耦合, 共同影响POPs在水生生态系统中的传递。据研究:① 底栖初级和次级生产力的贡献非常大, 大多鱼类对底栖初级、次级生产力的依赖程度甚至高于浮游初级和次级生产力[46-48]。② 底栖大型无脊椎动物在营养盐的循环和输入方面起到非常重要的作用[49]。③ 底栖细菌的生产力往往高于浮游细菌[50]。另外, 底栖生境是POPs在水生生态系统中循环流动的“汇”, 底栖生物通过摄食和被动吸收从沉积物中获取POPs, 并通过底栖食物网向上传递到顶级消费者体内, 是POPs循环的关键部分。但POPs在浮游和底栖食物网中的传递特征具有一定的差异。Kidd等[16]在马拉维湖研究发现, 同一污染物在浮游食物网中生物富集程度大于底栖食物网, 其原因是同一营养级的底栖 (藻类和腹足类等) 生物比浮游动物、毛翅类昆虫、腹足类生物的脂质含量低, 且底栖藻类的生长率更高, 产生“生长稀释”作用。Nfon等[24]发现PCBs在底栖食物链中的食物链放大系数要低于浮游食物链, 推测是由于浮游生物缺少分解PCBs的酶。但Campfens等[51]认为沉积物中POPs的逸度通常远大于水体, 底栖生物受到的污染会更严重。目前, 已普遍认识到底栖食物网对POPs环境行为的重要影响, 但底栖食物网在POPs传递过程中所起的作用仍莫衷一是, 传递机制尚不清晰, 未来还需要进一步的深入研究。
早期的研究中, 底栖食物网和浮游食物网被认为是互相独立的, 但是随着研究的深入, 逐渐认识到二者是通过各种方式耦合在一起的, 通过物质和能量的互相传递和交换共同支持生态系统的功能[52], POPs在此过程中随物质循环、能流流动出现耦合。一般情况下, 浮游-底栖食物网主要通过以下方式耦合:悬浮颗粒从水柱中沉降, 实现浮游生境对底栖动物的营养供给[53-56];底栖生物降解颗粒有机质为营养盐, 并再循环进入水柱[57-58], 底栖沉积物的再悬浮加强浮游-底栖耦合过程[59];杂食性鱼类捕食生境多变, 能广泛捕食浮游、底栖生物及一些陆地的无脊椎动物[47], 从而实现不同生境之间物质和能量的传递与交换[60-61]。在上述过程中, POPs随之一起实现浮游-底栖食物网的耦合, 因此只分析一种POPs传递路径, 不能准确地判断顶级捕食者体内POPs的来源和暴露风险。例如, 通过底栖-浮游食物网的耦合, 沉积物中的有机碳可以再循环进入到浮游生物体内, 同时将沉积物中与有机碳相络合的POPs通过食物网向上传递, 沉积物中积累多年的POPs使得水柱中捕食者体内的污染物浓度维持在一定的水平[15, 62]。糠虾等白天在湖泊底部摄食底栖生物, 晚上迁移到上层水柱中, 捕食浮游生物, 实现能量和POPs的浮游-底栖耦合。通过模拟糠虾的3种生活情景:① 只生活在浮游生境;② 部分时间生活在浮游生境, 部分时间生活在底栖生境;③ 只生活在底栖生境。模拟结果显示情景① 中糠虾体内的PCBs浓度最小, 情景③ 最大, 情景② 介于两者之间, 且其模拟结果与实测值最相符[92]。这说明了浮游-底栖食物网的耦合, 及耦合提高了浮游动物和鱼类的POPs暴露风险。
在过去, 底栖生境往往只作为污染物在水生生态系统中传递的“汇”, 且由于采样困难等原因, POPs在底栖食物网中环境行为的研究相对较少, 但了解POPs在底栖食物网中的环境行为是解析POPs在水生生态系统中归趋机制必不可少的内容, 需进一步加强研究。另外, 底栖食物网通过多种过程和浮游食物网耦合, 对POPs的环境行为产生复杂影响, 但目前对POPs在底栖-浮游耦合食物网中的归趋机制知之甚少, 是未来探索的一个重点。
3 微食物网对POPs环境行为的作用微生物, 包括病毒、细菌、鞭毛虫、纤毛虫、浮游植物和微型的浮游动物, 在生态系统中具有非常重要的作用, 这些生物形成一个非常复杂的微食物网。微食物网通过以下机制影响食物网的结构、功能:① 矿化分解有机质为浮游植物提供营养盐;② 有机碎屑消耗殆尽时, 细菌会与浮游植物竞争无机营养盐[63-64]。③ 细菌可以替代小型浮游动物, 缓解初级生产者的被捕食压力。④ 沉积物中的细菌是大型无脊椎动物的食物来源, 但有关这一点存在较大的争议[65-66]。研究表明, 微食物网在POPs的传递过程中也起到重要作用, 微生物吸附是POPs传递的一个重要途径, 可以通过微食物网向高营养级生物传递。微生物丰度高、体型小、周转速率快、具有最大的比表面积, 在水生生态系统中异养细菌通常占到了生物表面积的80%, 因此成为一个非常重要的POPs吸附体。且由于微生物体型较小, POPs在微生物与水相之间很快达到平衡, 几乎不受“生长稀释”的影响, 因此微生物成为比浮游植物更重要的POPs吸收介质[67]。另外, 颗粒物沉降过程中有机质被不断分解, 在一个较短的时间内, 细菌分解不会影响PCBs在颗粒物与水之间的分配, 导致PCBs浓度不断升高[68]。同时, 矿化分解提高了颗粒物的表面积, 增加了PCBs的吸附[69-70]。当颗粒物沉降到沉积物表层时, 微生物丰度较大, 有机质被大量分解, 然后大部分PCBs再次进入到水柱中, 进行再循环。由此可见, 微食物网对POPs在水生生态系统中的传递和再循环起到了非常重要的作用。另外, 众所周知, 微生物分解是POPs离开水生生态系统的重要途径, 但由于微食物网采集及分析存在诸多困难, 其相关研究还非常缺乏。
4 主要内外源压力下的食物网变化及对POPs环境行为的影响食物网是POPs在水生生态系统中传递的关键路径, 在受到一系列的内源驱动和外源压力时, 食物网的组成、结构和功能会发生很大的变化, 从而使食物网的动力学特征发生较大的变化。其中, 季节变化会使水生生态系统发生规律性变化, 探索由于季节演替导致的POPs在环境介质与食物网之间, 及食物网内部环境行为的规律性变化是生态学研究的重点之一;另外, 富营养化和外来物种入侵或引进导致的生态和环境灾变是目前国际上水生生态系统普遍存在的两个问题, 在这种外源压力下, 水生生态系统的结构和功能会发生显著变化, 甚至发生稳态转换, 从而使POPs在食物网中的环境行为和归趋机制发生变化。因此, 本文从季节变化、富营养化和外来物种入侵或引进这3个较为常见和普遍关注的内、外源压力入手, 探讨了食物网变化对POPs环境行为的影响。
4.1 食物网的季节演替对POPs传递的影响光照、温度、风和径流输入等的季节变化会导致水生生态系统的理化性质, 生物体的生理特征和种群结构发生周期性变化, 进而引起食物网结构和功能的变化, 使POPs的环境行为发生变化。
研究认为, 温度、光照、营养盐和水动力条件的变化使浮游植物的生物量及物种组成发生季节演替, 浮游植物是食物网中物质循环和能量流动的起点, 通过“上行控制”作用使高营养级消费者的栖息环境、生长率、食性等发生较大的变化, 进而引起整个食物网的变化。在诸多地区的浮游和底栖食物网中都观察到了明显的季节变化[31, 71-73]。研究表明, 水生生态系统中POPs浓度具有明显的季节变化, 7月份最低、9月底最高, 之后又不断下降, 其原因主要是由于藻类生产力的季节变化造成的[74-75]。Nizzetto[8]研究发现, 浮游动物体内的PCBs浓度在水华后期剧烈下降, 生物富集系数 (BAF) 最低, 不存在生物放大作用, 在水华爆发前期和爆发期存在明显的生物放大作用。因为在水华期间, 浮游动、植物快速生长, 生物量剧烈变化, 通过“生长稀释”和“生物量稀释”改变了PCBs的暴露风险。另外, 浮游动物物种组成、捕食者和生理学特征的季节性变化也会影响POPs的生物富集性。鱼类的生长率和种群结构与初级生产力和浮游生物的密度密切相关, 浮游动、植物的季节性演替, 使得鱼类的摄食习性也有周期性的变化, 进而导致鱼类POPs生物富集性的季节变化[44]。另外, 各种不同鱼类其单位脂质POPs浓度的季节变化不一样, 其原因目前还不是十分明确, 推测可能是食性不同和迁移方式不同造成的[27, 31]。
此外, 浮游植物的季节性生长会影响有机质的沉降, 有研究发现一年中大约有70%脂质沉降发生在春季水华时期, 与此同时POPs沉降量也最大, 底栖生境的POPs暴露风险增加[76], 而在冬季, 脂质的沉降量较低, POPs主要在浮游食物网中富集[67]。底栖群落的能量很大程度上依靠浮游生境的浮游碎屑[53], 同位素分析显示浮游植物是底栖无脊椎动物的主要食物来源[77]。浮游碎屑沉降量的显著季节变化, 使底栖群落的生存环境和食物资源发生显著的季节变化[65], 进而使底栖大型无脊椎动物的脂质含量发生季节变化[78], 而脂质含量是影响POPs生物富集的主要因素;另外, 摄食作用是生物体POPs的主要来源之一, 因此浮游生态系统中污染物的季节性变化会反映到底栖群落中, 但其变化趋势和幅度可能会存在差异, 因为很多底栖动物是杂食性的, 能够缓解季节变化的影响, 而且新沉降的物质能够很快地与老的沉积物混合 (生物扰动), 使得POPs分布特征发生混合。总体上, 底栖食物网生物富集性的季节性变化不如浮游食物网显著。另外, 不同底栖种群之间的季节变化趋势和幅度也各不相同, 因为它们的生态特征 (摄食模式、死亡率) 和生理特征 (氧吸收速率、生物转化能力、新陈代谢等) 不同, 这些都会影响污染物吸收、传递、排泄[31]。Hummel等[79]和Capuzzo等[80]的研究发现, 底栖软体动物体内的PCBs浓度受到季节性产卵和生殖的影响, 会发生明显的季节变化, 生殖会减少软体动物体内50%-66%的PCBs含量[81]。
环境理化性质和生物因素的季节变化造成POPs在环境和食物网中的分配和行为发生变化。如, 夏季水温升高, 底栖生物和微生物活性增强, 表层水流变慢和热分层出现, 加剧了底部的缺氧, 促进DDT厌氧分解, DDD浓度升高[82]。另外, 夏季水温升高改变生物的呼吸率和膜的通透性, POPs吸收速率加快, 生物富集性增强, POPs暴露风险升高[83]。
季节演替通过温度、风速、光照、DO等因素的周期性变化, 驱动食物网发生规律性变动, 造成POPs在食物网中的环境行为的季节性变化。环境介质、POPs的性质与食物网变化互相影响, 共同作用于POPs的环境行为, 但目前往往只考虑某一方面对POPs环境行为的影响, 对三者之间的联系关注不够。另外, 大多数研究只集中在食物网的某一环节, 如低营养级浮游食物网, 缺乏从整体角度的探索, 且研究的时间尺度较短, 多集中在某年的特定季节, 难以发掘其规律性变化。因此, 未来的研究需要从多角度、整体性和长时间尺度上深入研究POPs在水生食物网中环境行为的季节性变化。
4.2 富营养化条件下食物网变化对POPs环境行为的影响富营养化和POPs污染是水生态系统目前遇到的两个重要环境问题, 这两者之间往往是同时发生、互相联系的。传统上, 富营养化和POPs污染的研究通常是分开的, 但后来的研究发现, 营养水平能够影响污染物的循环和可生物利用水平[8](图 1)。瑞典环境保护局于1995年开展了揭示白令海附近海域及湖泊的富营养与污染物 (POPs, 微量金属) 之间互相联系的5年研究计划 (EUCON)[6]。

|
| 图 1 富营养化条件下食物网变化对POPs环境行为的影响 Fig. 1 Influences of foodweb changes caused by eutrophication on environmental behaviors of POPs |
富营养化会促进水-气界面的POPs输入[74], 或减少挥发[4], 改变食物网组成、多样性和优势种以及生态作用机制和能流传递效率、方向, 直接或间接地影响POPs的环境行为和传递机制[6, 84-85]。对于初级生产者, 富营养化促使浮游植物的优势种由蓝、绿藻取代硅藻、金藻等, 初级生产力大大提高, 生物量增加, 大型水生植物和底栖藻类死亡, 初级生产力从底栖为主导变为浮游占主导。由于蓝藻的食物品质较差, 使消费者的食物类型从浮游向底栖转变[86-87], 从而改变了食物网中物质流的传递方向, 同时改变了POPs沿食物网的流动路径。浮游生物POPs浓度降低, 其原因在于:相较于硅藻和金藻等, 蓝藻的脂质含量较低[88-89];且生物量大大增加导致“生物量稀释”, 生长速率较快, 发生“生长稀释”[90-93];另外, 在富营养条件下, 藻类的脂质含量相较于贫营养条件下更低[16, 67]。
对于浮游和底栖动物, 富营养化导致透光度下降, 附生藻类和大型水生植物死亡, ① 使得浮游和底栖动物的栖息、避难生境减少, 增强了鱼类的捕食作用[94-95], 大型无脊椎动物减少, 且小型化;② 蓝藻爆发, 可摄食的食物资源减少, 质量变差, 导致初级消费者丰度下降、优势种改变, 且食性发生变化。因此POPs在食物网中的传递路径发生变化;③ 藻类爆发导致有机质的沉降量增加。首先, 有机质大量分解造成沉积物-水界面缺氧[97], 底栖生物大量死亡, 优势种变为耐受低氧种[6, 87, 95-96], 使得底栖食物网发生较大的变化, POPs沿食物网的传递发生改变。其次, POPs随颗粒物沉降的通量增加[67, 97], 大部分POPs从水柱中移除, 降低了浮游生境的POPs暴露风险[98-99], 多项研究表明浮游食物链中POPs浓度与富营养化水平呈负相关[92, 100]。同时沉积物中POPs浓度升高[4, 89], 底栖食物网的暴露风险提高[75, 101-102]。
对于鱼类等高营养级生物, 富营养化会减少物种进化过程中对环境变化的生态响应, 导致物种灭绝, 降低多样性[103], 另外浮游和底栖群落的变化, 使鱼类的摄食食性、生长率和脂质含量都会发生较大的, 影响POPs的生物富集和传递效率[93, 104-105]。例如, Lasson等[98]对瑞典的61个湖泊研究发现, 鱼类体内PCBs生物富集系数 (BAF) 与湖泊生产力呈显著负相关, 因为富营养化条件下生产力提高, 鱼类生长加快, 发生“生长稀释”。另外, 鱼类等高营养级的生物, 其摄食吸收的POPs所占比例较大, 低营养级POPs浓度的变化会沿食物链影响鱼类体内的POPs浓度[93, 106]。但是上述几个不同的影响因素, 在不同的生态系统中或是不同的条件下, 起主导作用的因素不同。
对于微生物, 富营养化会直接或通过改变栖息生境的理化性质间接地影响微生物对POPs的分解速率, Graham等[107]研究认为富营养条件下微生物降解POPs的速率要高于寡营养水平, 但是目前这方面的研究相对较少。另如前文所述, 富营养化会扩大和加强底栖生境的缺氧程度, 促使微生物将DDT转化为DDD[82-83]。
4.3 物种入侵或引进对POPs环境行为的影响外来物种的入侵或引进是目前水生生态系统遇到的危机之一, 对食物网的组成、结构和动态变化产生了重要的影响, 例如, 生物体的食性、生长率、营养位等, 进而影响POPs的暴露风险, 及在食物网中的环境行为和传递机制。研究发现, 中营养级物种入侵会导致浮游动物和摄食浮游动物鱼类的营养位 (trophic position) 增加, 提高其捕食者体内的污染物浓度, 例如, 尾突蚤或胡瓜鱼等中营养级物种入侵后, 改变了浮游动物的群落结构, 优势种由枝角类变为桡足类, 浮游动物生物量降低[108-109], 提高了浮游动物和鱼类的营养位, 进而导致其体内的污染物浓度升高[9, 110]。另外, 在食物网中引入糠虾等新物种, 会延长食物链长度, 且由于糠虾的昼夜迁移行为, 会增加浮游-底栖食物网的耦合程度, 导致POPs在顶级消费者体内的浓度升高[2]。
但其它研究发现, 外来种入侵并不总是引起较高营养级生物污染物暴露风险的增加。在加拿大实验湖L227和L110中引入梭子鱼或肉食性白斑狗鱼后, 生态系统的群落结构、物种丰度和优势种都发生了改变, 很多鱼类的摄食习惯发生变化, 从主要摄食浮游动物转变为摄食底栖动物, 提高了营养位, 但鱼类体内的PCBs、DDT、HCH和Hg等污染物的浓度或是没有变化, 或是下降了, 推测这可能是由于营养级之间复杂的级联作用导致的[111]。例如, 中营养级物种入侵, 导致浮游动物减少, 浮游植物的生物量增加, 产生“生物量稀释”[109, 112-113], 或因为入侵的物种比传统猎物的营养位更高[110, 114]、密度更大[115], 消费者的捕食偏好改变, 觅食效率提高, 生长率更高[9], 从而导致“生长稀释”, 使污染物浓度下降。
物种入侵或是人工引入会通过营养级联作用对多个营养级产生不同程度的影响, 从而对食物网的结构、功能和生态过程产生影响, 进而导致POPs沿食物网的传递发生很大的变化, 但目前对这方面的研究还相对较少, 作用机制还不甚清晰。
5 结论与展望营养级和食物链长度是食物网的2个主要特征, 是研究POPs生物富集或生物放大时的重点考虑因素。一般情况下, POPs生物富集与营养级呈正相关, 但受到生物体各种生理、生态因素和POPs理化性质的影响, 有时会出现例外。各种影响因素纷繁复杂, 如何互相联系作用于营养级与POPs生物富集性之间的相关关系尚不十分明确, 需进一步探索。食物链长度与生物富集呈正相关, 除脂质的影响外, 其它具体作用机制还需深入研究。底栖食物网是POPs在水生生态系统中传递的一个重要途径, 但目前对POPs在底栖食物网中环境行为的认识莫衷一是, 尚不明确。另外, 底栖食物网通过各种过程与浮游食物网耦合, 对POPs的环境行为产生复杂的影响, 但相关研究甚少, 需要加强关注。微食物网对POPs环境行为的影响逐渐被关注, 目前相关研究主要集中在细菌对POPs的分解和吸附作用, 由于微生物体型小, 分析困难, 微食物网中POPs环境行为的动态研究和定量研究还相对缺乏。在内源驱动和外源压力的作用下, 比如季节演替、富营养化、物种入侵等, 使环境介质、食物网和POPs的结构、性质发生复杂的变化, 三者之间互相反馈导致POPs沿食物网的环境行为发生更为错综复杂的变化, 但是目前这方面的研究还不够全面和系统, 是研究的重点和难点。
目前, POPs在食物网中环境行为的研究主要集中在“classic” POPs, 包括PCBs、OCPs和PAHs等, 但在传统POPs被禁用多年后, 很多替代性的新型POPs被越来越多地使用, 并在世界范围内的不同环境中都有检出。例如, 在中国, 海洋哺乳动物和沉积物中传统PBDE开始逐渐被新型阻燃剂BFRs替代[116-117], 渤海地区更是遭到了新型POPs的严重污染[118]。在南、北极地区也分别都检测到了新型POPs, 并认为存在生物放大的可能。目前对于新型POPs可以获取的数据还很少[119-121], 对于其生物富集性的研究也多集中在少数地区的极少数大型哺乳动物和鱼类中。未来, 需要加大对新型POPs在水生食物网中的研究。
| [1] | Kelly B C, Ikonomou M G, Blair J D, Morin A E, Gobas F A P C. Food web-specific biomagnification of persistent organic pollutants. Science, 2007, 317(5835): 236–239. DOI:10.1126/science.1138275 |
| [2] | Rasmussen J B, Rowan D J, Lean D R S, Carey J H. Food chain structure in Ontario Lakes determines PCB levels in Lake Trout (salvelinus namaycush) and other pelagic fish. Canadian Journal of Fisheries and Aquatic Sciences, 1990, 47(10): 2030–2038. DOI:10.1139/f90-227 |
| [3] | Wan Y, Zhang K, Dong Z M, Hu J Y. Distribution is a major factor affecting bioaccumulation of decabrominated diphenyl ether: Chinese sturgeon (Acipenser sinensis) as an example. Environmental Science and Technology, 2013, 47(5): 2279–2286. DOI:10.1021/es304926r |
| [4] | Jeremiason J D, Eisenreich S J, Paterson M J, Beatly K G, Hecky R, Elser J J. Biogeochemical cycling of PCBs in lakes of variable trophic status: a paired-lake experiment. Limnology and Oceanography, 1999, 44(3): 889–902. |
| [5] | 李雪梅, 张庆华, 甘一萍, 周军, 戴家银, 曹宏, 许木启. 持久性有机污染物在食物链中积累与放大研究进展. 应用与环境生物学报, 2008, 13(6): 901–905. |
| [6] | Skei J, Larsson P, Rosenberg R, Jonsson P, Olsson M, Broman D. Eutrophication and contaminants in aquatic ecosystem. AMBIO, 2000, 29(4-5): 184–194. |
| [7] | Gunnarsson J, Björk M, Gilek M, Granberg M, Rosenberg R. Effects of eutrophication on contaminant cycling in marine benthic systems. AMBIO, 2000, 29(4-5): 252–259. |
| [8] | Nizzetto L, Gioia R, Li J, Borga K, Pomati F, Bettinetti R, Dachs J, Jones K C. Biological pump control of the fate and distribution of hydrophobic organic pollutants in water and plankton. Environmental Science and Technology, 2012, 46(6): 3204–3211. DOI:10.1021/es204176q |
| [9] | Rennie M D, Strecker A L, Palmer M E. Bythotrephes invasion elevates trophic position of zooplankton and fish: implications for contaminant biomagnification. Biological Invasions, 2011, 13(11): 2621–2634. DOI:10.1007/s10530-011-0081-0 |
| [10] | Houde M, Pacepavicius G, Wells R S, Fair P A, Letcher R J, Alaee M, Bossart G D, Hohn A A, Sweeney J, Solomon K R, Derek C G. Polychlorinated bipphenyls and hydroxylated polychlorinated biphenyls in plasma of bottlenose dolphins (Tursiops truncatus) from the Western Atlantic and the Gulf of Mexico. Environmental Science and Technology, 2006, 40(19): 5860–5866. DOI:10.1021/es060629n |
| [11] | Kannan K, Tao L, Sinclair E, Pastva S D, Jude D J, Giesy J P. Perfluorinated compounds in aquatic organisms at various trophic levels in a Great Lakes food chain. Archives of Environmental contamination and Toxicology, 2005, 48(4): 559–566. DOI:10.1007/s00244-004-0133-x |
| [12] | Martin J W, Mabury S A, Solomon K R, Muir D C G. Dietary accumulation of perfluorinated acids in juvenile rainbow trout (Oncorhynchus mykiss). Environmental Toxicology and Chemistry, 2003, 22(1): 189–195. DOI:10.1002/etc.v22:1 |
| [13] | Nfon E, Cousins I T. Interpreting time trends and biomagnification of PCBs in the Baltic region using the equilibrium lipid partitioning approach. Environmental Pollution, 2006, 144(3): 994–1000. DOI:10.1016/j.envpol.2006.01.042 |
| [14] | Hobson K A, Fisk A, Karnovsky N, Holst M, Gagnon J M, Fortier M. A stable isotope (δ13C, δ15N) model for the North Water food web: implications for evaluating trophodynamics and the flow of energy and contaminants. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2002, 49(22-23): 5131–5150. DOI:10.1016/S0967-0645(02)00182-0 |
| [15] | Kucklick J R, Baker J E. Organochlorines in Lake Superior's food web. Environmental Science and Technology, 1998, 32(9): 1192–1198. DOI:10.1021/es970794q |
| [16] | Kidd K A, Bootsma H A, Hesslein R H, Muir D C G, Hecky R E. Biomagnification of DDT through the benthic and pelagic food webs of Lake Malawi, East Africa: importance of trophic level and carbon source. Environmental Science and Technology, 2001, 35(1): 14–20. DOI:10.1021/es001119a |
| [17] | Neff J M. Bioaccumulation of organic micropollutants from sediments and suspended particulates by aquatic animals. Fresenius' Zeitschrift für Analytische Chemie, 1984, 319(2): 132–136. DOI:10.1007/BF00584674 |
| [18] | Varanasi U, Reichert W L, Stein J E, Brown D W, Sanborn H R. Bioavailability and biotransformation of aromatic hydrocarbons in benthic organisms exposed to sediment from an urban estuary. Environmental Science and Technology, 1985, 19(9): 836–841. DOI:10.1021/es00139a012 |
| [19] | Broman D, Näuf C, Lundbergh I, Zebühr Y. An in situ study on the distribution, biotransformation and flux of polycyclic aromatic hydrocarbons (PAHs) in an aquatic food chain (Seston-Mytilus edulis L.-Somateria mollissima L.) from the baltic: an ecotoxicological perspective. Environmental Toxicology and Chemistry, 1990, 9(4): 429–442. |
| [20] | Niimi A J, Oliver B G. Influence of molecular weight and molecular volume on dietary absorption efficiency of chemicals by fishes. Canadian Journal of Fisheries and Aquatic Sciences, 1988, 45(2): 222–227. DOI:10.1139/f88-026 |
| [21] | Selck H, Palmqvist A, Forbes V E. Biotransformation of dissolved and sediment-bound fluoranthene in the polychaete, Capitella Sp. I.. Environmental Toxicology and Chemistry, 2003, 22(10): 2364–2374. DOI:10.1897/02-272 |
| [22] | Gewurtz S B, Lazar R, Haffner G D. Comparison of polycyclic aromatic hydrocarbon and polychlorinated biphenyl dynamics in benthic invertebrates of Lake Erie, USA. Environmental Toxicology and Chemistry, 2000, 19(12): 2943–2950. DOI:10.1002/etc.v19:12 |
| [23] | Mizukawa K, Takada H, Takeuchi I, Ikemoto T, Omori K, Tsuchiya K. Bioconcentration and biomagnification of polybrominated diphenyl ethers (PBDEs) through lower-trophic-level coastal marine food web. Marine Pollution Bulletin, 2009, 58(8): 1217–1224. DOI:10.1016/j.marpolbul.2009.03.008 |
| [24] | Nfon E, Cousins I T, Broman D. Biomagnification of organic pollutants in benthic and pelagic marine food chains from the Baltic Sea. Science of the Total Environment, 2008, 397(1/3): 190–204. |
| [25] | Hanari N, Kannan K, Horii Y, Taniyasu S, Yamashita N, Jude D J, Berg M B. Polychlorinated naphthalenes and polychlorinated biphenyls in benthic organisms of a Great Lakes food chain. Archives of Environmental Contamination and Toxicology, 2004, 47(1): 84–93. |
| [26] | Corsilini S, Kannan K, Imagawa T, Focardi S, Giesy J P. Polychloronaphthalenes and other dioxin-like compounds in Arctic and Antarctic marine food webs. Environmental Science and Technology, 2002, 36(16): 3490–3496. DOI:10.1021/es025511v |
| [27] | Hallanger I G, Warner N A, Ruus A, Evenset A, Christensen G, Herzke D, Gabrielsen G W, Borgå K. Seasonality in contaminant accumulation in Arctic marine pelagic food webs using trophic magnification factor as a measure of bioaccumulation. Environmental Toxicology and Chemistry, 2011, 30(5): 1026–1035. DOI:10.1002/etc.v30.5 |
| [28] | Borgå K, Kidd K A, Muir D C G, Berglund O, Conder J M, Gobas F A P C, Kucklick J, Malm O, Powell D. Trophic magnification factors: considerations of ecology, ecosystems and study design. Integrated Environmental Assessment and Management, 2012, 8(1): 64–84. DOI:10.1002/ieam.244 |
| [29] | Binelli A, Provini A. The PCB pollution of Lake Iseo (N. Italy) and the role of biomagnification in the pelagic food web.. Chemosphere, 2003, 53(2): 143–151. DOI:10.1016/S0045-6535(03)00441-7 |
| [30] | Lundgren K, Tysklind M, Ishaq R, Broman D, Van Bavel B. Polychlorinated naphthalene levels, distribution, and biomagnification in a benthic food chain in the Baltic Sea. Environmental Science and Technology, 2002, 36(23): 5005–5013. DOI:10.1021/es0201146 |
| [31] | Evenset A, Hallanger I G, Tessmann M, Warner N, Ruus A, Borgå K, Gabrielsen G W, Christensen G, Renaud P E. Seasonal variation in accumulation of persistent organic pollutants in an Arctic marine benthic food web. Science of the Total Environment, 2016, 542: 108–120. DOI:10.1016/j.scitotenv.2015.10.092 |
| [32] | Paterson G, Ryder M, Drouillard K G, Haffner G D. Contrasting PCB bioaccumulation patterns among Lake Huron lake trout reflect basin-specific ecology. Environmental Toxicology and Chemistry, 2016, 35(1): 65–73. DOI:10.1002/etc.3179 |
| [33] | McLeod A M, Paterson G, Drouillard K G, Haffner G D. Ecological factors contributing to variability of persistent organic pollutant bioaccumulation within forage fish communities of the Detroit River, Ontario, Canada. Environmental Toxicology and Chemistry, 2014, 33(8): 1825–1831. DOI:10.1002/etc.2606 |
| [34] | Jardine T D, Kidd K A, Fisk A T. Applications, considerations, and sources of uncertainty when using stable isotope analysis in ecotoxicology. Environmental Science and Technology, 2006, 40(24): 7501–7511. DOI:10.1021/es061263h |
| [35] | Bruner K A, Fisher S W, Landrum P F. The role of the zebra mussel, Dreissena polymorpha, in contaminant cycling: Ⅱ. Zebra Mussel contaminant accumulation from algae and suspended particles, and transfer to the benthic invertebrate, Gammarus fasciatus. Journal of Great Lakes Research, 1994, 20(4): 735–750. DOI:10.1016/S0380-1330(94)71191-6 |
| [36] | Looser R, Ballschmiter K. Biomagnification of polychlorinated biphenyls (PCBs) in freshwater fish. Fresenius' Journal of Analytical Chemistry, 1998, 360(7/8): 816–819. |
| [37] | Gobas F A P C, Wilcockson J B, Russell R W, Haffner G D. Mechanism of biomagnification in fish under laboratory and field conditions. Environmental Science and Technology, 1999, 33(1): 133–141. DOI:10.1021/es980681m |
| [38] | Borgå K, Gabrielsen G W, Skaare J U. Biomagnification of organochlorines along a Barents sea food chain. Environmental Pollution, 2001, 113(2): 187–198. DOI:10.1016/S0269-7491(00)00171-8 |
| [39] | Guruge K S, Tanabe S, Iwata H, Taksukawa R, Yamagishi S. Distribution, biomagnification, and elimination of butyltin compound residues in common cormorants (Phalacrocorax carbo) from Lake Biwa, Japan. Archives of Environmental Contamination and Toxicology, 1996, 31(2): 210–217. DOI:10.1007/BF00212368 |
| [40] | Bentzen E, Lean D R S, Taylor W D, Mackay D. Role of food web structure on lipid bioaccumulation of organic contaminants by lake trout (Salvelinus namaycush). Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53(11): 2397–2407. DOI:10.1139/f96-196 |
| [41] | Oliver B G, Niimi A J. Trophodynamic analysis of polychlorinated biphenyl congeners and other chlorinated hydrocarbons in the Lake Ontario ecosystem. Environmental Science and Technology, 1988, 22(4): 388–397. DOI:10.1021/es00169a005 |
| [42] | Evans M S, Noguchi G E, Rice C P. The biomagnification of polychlorinated biphenyls, toxaphene, and DDT compounds in a Lake Michigan offshore food web. Archives of Environmental Contamination and Toxicology, 1991, 20(1): 87–93. DOI:10.1007/BF01065333 |
| [43] | Kidd K A, Schindler D W, Muir D C G, Lockhart W L, Hesslein R H. High concentrations of toxaphene in fishes from a subarctic lake. Science, 1995, 269(5221): 240–242. DOI:10.1126/science.269.5221.240 |
| [44] | Whittle D M, Kiriluk R M, Carswell A A, Keir M J, MacEachen D C. Toxaphene congeners in the Canadian Great Lakes basin: temporal and spatial food web dynamics. Chemosphere, 2000, 40(9-11): 1221–1226. DOI:10.1016/S0045-6535(99)00372-0 |
| [45] | Kerr S R. A simulation model of lake trout growth. Journal of the Fisheries Research Board of Canada, 1971, 28(6): 815–819. DOI:10.1139/f71-122 |
| [46] | Hecky R E, Hesslein R H. Contributions of benthic algae to lake food webs as revealed by stable isotope analysis. Journal of the North American Benthological Society, 1995, 14(4): 631–653. DOI:10.2307/1467546 |
| [47] | Vander Zanden M J, Vadeboncoeur Y. Fishes as integrators of benthic and pelagic food webs in lakes. Ecology, 2002, 83(8): 2152–2161. DOI:10.1890/0012-9658(2002)083[2152:FAIOBA]2.0.CO;2 |
| [48] | Strayer D, Likens G E. An energy budget for the zoobenthos of Mirror Lake, New Hampshire. Ecology, 1986, 67(2): 303–313. DOI:10.2307/1938574 |
| [49] | Covich A P, Palmer M A, Crowl T A. The Role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling. BioScience, 1999, 49(2): 119–127. DOI:10.2307/1313537 |
| [50] | Theil-Nielsen J, Søndergaard M. Production of epiphytic bacteria and bacterioplankton in three shallow lakes. Oikos, 1999, 86(2): 283–292. DOI:10.2307/3546446 |
| [51] | Campfens J, Mackay D. Fugacity-based model of PCB bioaccumulation in complex aquatic food webs. Environmental Science and Technology, 1997, 31(2): 577–583. DOI:10.1021/es960478w |
| [52] | Vadeboncoeur Y, Vander Zanden M J, Lodge D M. Putting the lake back together: reintegrating benthic pathways into lake food web models. BioScience, 2002, 52(1): 44–54. DOI:10.1641/0006-3568(2002)052[0044:PTLBTR]2.0.CO;2 |
| [53] | Johnson R K, Wiederholm T. Pelagic-benthic coupling-the importance of diatom interannual variability for population oscillations of Monoporeia affinis. Limnology and Oceanography, 1992, 37(8): 1596–1607. DOI:10.4319/lo.1992.37.8.1596 |
| [54] | Jonasson P M. Ecology and production of the profundal benthos in relation to phytoplankton in Lake Esrom. Oikos, 1972, 14(Supplement): 1–148. |
| [55] | Johnson R K, Boström B, Van De Bund W. Interactions between Chironomus plumosus (L.) and the microbial community in surficial sediments of a shallow, eutrophic lake. Limnology and Oceanography, 1989, 34(6): 992–1003. DOI:10.4319/lo.1989.34.6.0992 |
| [56] | Rasmussen J B. Effects of density and microdetritus enrichment on the growth of chironomid larvae in a small pond. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(8): 1418–1422. DOI:10.1139/f85-177 |
| [57] | Kemp M W, Boynton W R. Spatial and temporal coupling of nutrient inputs to estuarine primary production: the role of particulate transport and decomposition. Bulletin of Marine Science, 1984, 35(3): 522–535. |
| [58] | Strayer D L. Perspectives on the size structure of lacustrine zoobenthos, its causes, and its consequences. Journal of the North American Benthological Society, 1991, 10(2): 210–221. DOI:10.2307/1467579 |
| [59] | Doering P H, Kelly J R, Oviatt C A, Sowers T. Effect of the hard clam Mercenaria mercenia on benthic fluxes of inorganic nutrients and gases. Marine Biology, 1987, 94(3): 377–383. DOI:10.1007/BF00428243 |
| [60] | Jeppesen E, Jensen J P, S∅ndergaard M, Lauridsen T, Pedersen L J, Jensen L. Top-down control in freshwater lakes: the role of nutrient state, submerged macrophytes and water depth. Hydrobiologia, 1997, 342-343: 151–164. DOI:10.1023/A:1017046130329 |
| [61] | Stein R A, DeVries D R, Dettmers J M. Food-web regulation by a planktivore: exploring the generality of the trophic cascade hypothesis. Canadian Journal of Fisheries and Aquatic Sciences, 1995, 52(11): 2518–2526. DOI:10.1139/f95-842 |
| [62] | Klump J V, Kaster J L, Sierszen M E. Mysis relicta assimilation of Hexachlorobiphenyl from sediments. Canadian Journal of Fisheries and Aquatic Sciences, 1991, 48(2): 284–289. DOI:10.1139/f91-039 |
| [63] | Danger M, Oumarou C, Benest D, Lacroix G. Bacteria can control stoichiometry and nutrient limitation of phytoplankton. Functional Ecology, 2007, 21(2): 202–210. DOI:10.1111/fec.2007.21.issue-2 |
| [64] | Joint I, Henriksen P, Fonnes G A, Bourne D, Thingstad T F, Riemann B. Competition for inorganic nutrients between phytoplankton and bacterioplankton in nutrient manipulated mesocosms. Aquatic Microbial Ecology, 2002, 29(2): 145–159. |
| [65] | Goedkoop W, Johnson R K. Pelagic-benthic coupling: profundal benthic community response to spring diatom deposition in mesotrophic Lake Erken. Limnology and Oceanography, 1996, 41(4): 636–647. DOI:10.4319/lo.1996.41.4.0636 |
| [66] | Goedkoop W, Johnson R K. Exploitation of sediment bacterial carbon by juveniles of the amphipod Monoporeia affinis. Freshwater Biology, 1994, 32(3): 553–563. DOI:10.1111/fwb.1994.32.issue-3 |
| [67] | Larsson P, Andersson A, Broman D, Nordbäck J, Lundberg E. Persistent organic pollutants (POPs) in pelagic systems. AMBIO, 2000, 29(4-5): 202–209. |
| [68] | Axelman J, Broman D, Näf C. Vertical flux and particulate/water dynamics of polychlorinated biphenyls (PCBs) in the open Baltic Sea. AMBIO, 2000, 29(4-5): 210–216. |
| [69] | Koelmans A A, Lijklema L, Jiménez C S. Sorption of chlorobenzenes to mineralizing phytoplankton. Environmental Toxicology and Chemistry, 1993, 12(8): 1425–1439. DOI:10.1002/etc.v12:8 |
| [70] | Koelmans A A, Anzion S F M, Lijklema L. Dynamics of organic micropollutant biosorption to cyanobacteria and detritus. Environmental Science and Technology, 1995, 29(4): 933–940. DOI:10.1021/es00004a013 |
| [71] | Fisk A T, Stern G A, Hobson K A, Strachan W J, Loewen M D, Norstrom R J. Persistent organic pollutants (POPs) in a small, herbivorous, Arctic marine zooplankton (Calanus hyperboreus): trends from April to July and the influence of lipids and trophic transfer. Marine Pollution Bulletin, 2001, 43(1/6): 93–101. |
| [72] | Hargrave B T, Phillips G A, Vass W P, Bruecker P, Welch H E, Siferd T D. Seasonality in bioaccumulation of organochlorines in lower trophic level Arctic marine biota. Environmental Science and Technology, 2000, 34(6): 980–987. DOI:10.1021/es990871c |
| [73] | Hallanger I G, Ruus A, Herzke D, Warner N A, Evenset A, Heimstad E S, Gabrielsen G W, Borgå K. Influence of season, location, and feeding strategy on bioaccumulation of halogenated organic contaminants in Arctic marine zooplankton. Environmental Toxicology and Chemistry, 2011, 30(1): 77–87. DOI:10.1002/etc.362 |
| [74] | Dachs J, Eisenreich S J, Baker J E, Ko F C, Jeremiason J D. Coupling of phytoplankton uptake and air-water exchange of persistent organic pollutants. Environmental Science and Technology, 1999, 33(20): 3653–3660. DOI:10.1021/es990168o |
| [75] | Söderström M, Nylund K, Järnberg U, Lithner G, Rosén G, Kylin H. Seasonal variations of DDT compounds and PCB in a eutrophic and an oligotrophic lake in relation to algal biomass. AMBIO, 2000, 29(4-5): 230–237. |
| [76] | Chi J, Wang Q Y, Huang J J, Huang G L. Sedimentation and seasonal variation of hexachlorocyclohexanes in sediments in a eutrophic lake, China. Journal of Environmental Science and Health, Part B: Pesticides, Food Contaminants, and Agricultural Wastes, 2008, 43(7): 611–616. DOI:10.1080/03601230802234724 |
| [77] | Renaud P E, Tessmann M, Evenset A, Christensen G N. Benthic food-web structure of an Arctic fjord (Kongsfjorden, Svalbard). Marine Biology Research, 2011, 7(1): 13–26. DOI:10.1080/17451001003671597 |
| [78] | Gardner W S, Nalepa T F, Frez W A, Cichocki E A, Landrum P F. Seasonal patterns in lipid content of lake michigan macroinvertebrates. Canadian Journal of Fisheries and Aquatic Sciences, 1985, 42(11): 1827–1832. DOI:10.1139/f85-229 |
| [79] | Hummel H, Bogaards R H, Nieuwenhuize J, De Wolf L, Van Liere J M. Spatial and seasonal differences in the PCB content of the mussel Mytilus edulis. Science of the Total Environment, 1990, 92: 155–163. DOI:10.1016/0048-9697(90)90327-Q |
| [80] | Capuzzo J M, Farrington J W, Rantamaki P, Clifford C H, Lancaster B A, Leavitt D F, Jia Xiaoping. The relationship between lipid composition and seasonal differences in the distribution of PCBs in Mytilus Edulis L. Marine Environmental Research, 1989, 28(1/4): 259–264. |
| [81] | Person N J, Axelman J, Broman D. Validating possible effects of eutrophication using PCB concentrations in bivalves and sediment of the US musselwatch and benthic surveillance programs. AMBIO, 2000, 29(4-5): 246–251. |
| [82] | Kale S P, Murthy N B K, Raghu K, Sherkhane P D, Carvalho F P. Studies on degradation of 14C-DDT in the marine environment. Chemosphere, 1999, 39(6): 959–968. DOI:10.1016/S0045-6535(99)00027-2 |
| [83] | Sethajintanin D, Anderson K A. Temporal bioavailability of organochlorine pesticides and PCBs. Environmental Science and Technology, 2006, 40(12): 3689–3695. DOI:10.1021/es052427h |
| [84] | Horner-Devine M C, Leibold M A, Smith V H, Bohannan B J M. Bacterial diversity patterns along a gradient of primary productivity. Ecology Letters, 2003, 6(7): 613–622. DOI:10.1046/j.1461-0248.2003.00472.x |
| [85] | Smith V H. Microbial diversity-productivity relationships in aquatic ecosystems. FEMS Microbiology Ecology, 2007, 62(2): 181–186. DOI:10.1111/fem.2007.62.issue-2 |
| [86] | Carlier A, Riera P, Amouroux J M, Bodiou J Y, Desmalades M, Grémare A. Food web structure of two Mediterranean lagoons under varying degree of eutrophication. Journal of Sea Research, 2008, 60(4): 264–275. DOI:10.1016/j.seares.2008.10.006 |
| [87] | Vadeboncoeur Y, Jeppesen E, Vander Zanden M J, Schierup H H, Christoffersen K, Lodge D M. From Greenland to green lakes: cultural eutrophication and the loss of benthic pathways in lakes. Limnology and Oceanography, 2003, 48(4): 1408–1418. DOI:10.4319/lo.2003.48.4.1408 |
| [88] | Borgå K, Fisk A T, Hoekstra P F, Muir D C G. Biological and chemical factors of importance in the bioaccumulation and trophic transfer of persistent organochlorine contaminants in arctic marine food webs. Environmental Toxicology and Chemistry, 2004, 23(10): 2367–2385. DOI:10.1897/03-518 |
| [89] | Larsson P, Okla L, Cronberg G. Turnover of polychlorinated biphenyls in an oligotrophic and a eutrophic lake in relation to internal lake processes and atmospheric fallout. Canadian Journal of Fisheries and Aquatic Sciences, 1998, 55(8): 1926–1937. DOI:10.1139/f98-061 |
| [90] | Niimi A J, Cho C Y. Elimination of hexachlorobenzene (HCB) by rainbow trout (Salmo gairdneri), and an examination of its kinetics in lake Ontario Salmonids. Canadian Journal of Fisheries and Aquatic Sciences, 1981, 38(11): 1350–1356. DOI:10.1139/f81-181 |
| [91] | Axelman J, Broman D, Näf C. Field measurements of PCB partitioning between water and planktonic organisms: influence of growth, particle size, and solute-solvent interactions. Environmental Science and Technology, 1997, 31(3): 665–669. DOI:10.1021/es960088+ |
| [92] | Taylor W D, Carey J H, Lean D R S, McQueen D J. Organochlorine concentrations in the plankton of lakes in Southern Ontario and their relationship to plankton biomass. Canadian Journal of Fisheries and Aquatic Sciences, 1991, 48(10): 1960–1966. DOI:10.1139/f91-233 |
| [93] | Ryan M J, Stern G A, Kidd K A, Croft M V, Gewurtz S, Diamond M, Kinnear L, Roach P. Biotic interactions in temporal trends (1992-2010) of organochlorine contaminants in the aquatic food web of lake Laberge, Yukon Territory. Science of the Total Environment, 2013, 443: 80–92. DOI:10.1016/j.scitotenv.2012.10.055 |
| [94] | Jeppesen E, Jensen J P, Søndergaard M, Lauridsen T. Trophic dynamics in turbid and clearwater lakes with special emphasis on the role of zooplankton for water clarity. Hydrobiologia, 1999, 408-409: 217–231. DOI:10.1023/A:1017071600486 |
| [95] | Rasmussen J B, Kalff J. Empirical models for zoobenthic biomass in lakes. Canadian Journal of Fisheries and Aquatic Sciences, 1987, 44(5): 990–1001. DOI:10.1139/f87-117 |
| [96] | Kemp W M, Sampou P A, Garber J, Tuttle J, Boynton W R. Seasonal depletion of oxygen from bottom waters of Chesapeake Bay: roles of benthic and planktonic respiration and physical exchange processes. Marine Ecology Progress Series, 1992, 85: 137–152. DOI:10.3354/meps085137 |
| [97] | Auer B, Elzer U, Arndt H. Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: influence of resource and predation. Journal of Plankton Research, 2004, 26(6): 697–709. DOI:10.1093/plankt/fbh058 |
| [98] | Larsson P, Collvin L, Okla L, Meyer G. Lake productivity and water chemistry as governors of the uptake of persistent pollutants in fish. Environmental Science and Technology, 1992, 26(2): 346–352. DOI:10.1021/es00026a016 |
| [99] | Millard E S, Minns C K, Charlton C C, Halfon E. Effect of primary productivity and vertical mixing on pcb dynamics in planktonic model ecosystems. Environmental Toxicology and Chemistry, 1993, 12(5): 931–946. DOI:10.1002/etc.v12:5 |
| [100] | Berglund O, Larsson P, Ewald G, Okla L. The effect of lake trophy on lipid content and PCB concentrations in planktonic food webs. Ecology, 2001, 82(4): 1078–1088. DOI:10.1890/0012-9658(2001)082[1078:TEOLTO]2.0.CO;2 |
| [101] | Gunnarsson J S, Schaanning M T, Hylland K, Sköld M, Eriksen D Ø, Berge J A, Skei J. Interactions between eutrophication and contaminants. Ⅲ. Mobilization and bioaccumulation of benzo (a) pyrene from marine sediments. Marine Pollution Bulletin, 1996, 33(1/6): 80–89. |
| [102] | Franz J M S, Hauss H, Sommer U, Dittmar T, Riebesell U. Production, partitioning and stoichiometry of organic matter under variable nutrient supply during mesocosm experiments in the tropical Pacific and Atlantic Ocean. Biogeosciences, 2012, 9(11): 4629–4643. DOI:10.5194/bg-9-4629-2012 |
| [103] | Vonlanthen P, Bittner D, Hudson A G, Young K A, Müller R, Lundsgaard-Hansen B, Roy D, Di Piazza S, Largiader C R, Seehausen O. Eutrophication causes speciation reversal in whitefish adaptive radiations. Nature, 2012, 482(7385): 357–362. DOI:10.1038/nature10824 |
| [104] | Thomann R V. Bioaccumulation model of organic chemical distribution in aquatic food chains. Environmental Science and Technology, 1989, 23(6): 699–707. DOI:10.1021/es00064a008 |
| [105] | Hammar J, Larsson P, Klavins M. Accumulation of persistent pollutants in normal and dwarfed Arctic Char (Salvelinus alpinus sp. complex). Canadian Journal of Fisheries and Aquatic Sciences, 1993, 50(12): 2574–2580. DOI:10.1139/f93-281 |
| [106] | Smith D W. Synchronous response of hydrophobic chemicals in herring gull eggs from the Great Lakes. Environmental Science and Technology, 1995, 29(3): 740–750. DOI:10.1021/es00003a023 |
| [107] | Graham D W, Miles D, De Noyelles F, Smith V H. Development of small outdoor microcosms for studying contaminant transformation rates and mechanisms under various water column conditions. Environmental Toxicology and Chemistry, 1999, 18(6): 1124–1132. DOI:10.1002/etc.v18:6 |
| [108] | Fernandez R J, Rennie M D, Sprules W G. Changes in nearshore zooplankton associated with species invasions and potential effects on larval lake whitefish (Coregonus clupeaformis). International Review of Hydrobiology, 2009, 94(2): 226–243. DOI:10.1002/iroh.v94:2 |
| [109] | Strecker A L, Arnott S E. Invasive predator, Bythotrephes, has varied effects on ecosystem function in freshwater lakes. Ecosystems, 2008, 11(3): 490–503. DOI:10.1007/s10021-008-9137-0 |
| [110] | Vander Zanden M J, Rasmussen J B. A trophic position model of pelagic food webs: impact on contaminant bioaccumulation in lake trout. Ecological Monographs, 1996, 66(4): 451–477. DOI:10.2307/2963490 |
| [111] | Kidd K A, Paterson M J, Hesslein R H, Muir D C G, Hecky R E. Effects of northern pike (Esox lucius) additions on pollutant accumulation and food web structure, as determined by δ13C and δ15N, in a eutrophic and an oligotrophic lake. Canadian Journal of Fisheries and Aquatic Sciences, 1999, 56(11): 2193–2202. DOI:10.1139/f99-148 |
| [112] | Swanson H K, Johnston T A, Schindler D W, Bodaly R A, Whittle D W. Mercury bioaccumulation in forage fish communities invaded by rainbow smelt (Osmerus mordax). Environmental Science and Technology, 2006, 40(5): 1439–1446. DOI:10.1021/es0510156 |
| [113] | Hogan L S, Marschall E, Folt C, Stein R A. How non-native species in Lake Erie influence trophic transfer of mercury and lead to top predators. Journal of Great Lakes Research, 2007, 33(1): 46–61. DOI:10.3394/0380-1330(2007)33[46:HNSILE]2.0.CO;2 |
| [114] | Campbell L M, Thacker R, Barton D, Muir D C G, Greenwood D, Hecky R E. Re engineering the eastern Lake Erie littoral food web: the trophic function of non-indigenous Ponto-Caspian species. Journal of Great Lakes Research, 2009, 35(2): 224–231. DOI:10.1016/j.jglr.2009.02.002 |
| [115] | Coulas R A, Macisaac H J, Dunlop W. Selective predation on an introduced zooplankter (Bythotrephes cederstroemi) by lake herring (Coregonus artedii) in Harp Lake, Ontario. Freshwater Biology, 1998, 40(2): 343–355. DOI:10.1046/j.1365-2427.1998.00340.x |
| [116] | Chen S J, Feng A H, He M J, Chen M Y, Luo X J, Mai B X. Current levels and composition profiles of PBDEs and alternative flame retardants in surface sediments from the Pearl River Delta, southern China: comparison with historical data. Science of the Total Environment, 2013, 444: 205–211. DOI:10.1016/j.scitotenv.2012.11.104 |
| [117] | Zhu B Q, Lai N L S, Wai T C, Chan L L, Lam J C W, Lam P K S. Changes of accumulation profiles from PBDEs to brominated and chlorinated alternatives in marine mammals from the South China Sea. Environment International, 2014, 66: 65–70. DOI:10.1016/j.envint.2014.01.023 |
| [118] | Li X M, Gao Y, Wang Y W, Pan Y Y. Emerging persistent organic pollutants in Chinese Bohai Sea and its coastal regions. The Scientific World Journal, 2014, 2014: 1–10. |
| [119] | Kim J T, Son M H, Kang J H, Kim J H, Jung J W, Chang Y S. Occurrence of legacy and new persistent organic pollutants in avian tissues from King George Island, Antarctica. Environmental Science and Technology, 2015, 49(22): 13628–13638. DOI:10.1021/acs.est.5b03181 |
| [120] | Vorkamp K, Bossi R, Rigét F F, Skov H, Sonne C, Dietz R. Novel brominated flame retardants and dechlorane plus in Greenland air and biota. Environmental Pollution, 2015, 196: 284–291. DOI:10.1016/j.envpol.2014.10.007 |
| [121] | Wolschke H, Meng X Z, Xie Z Y Ebinghaus R, Cai M H. Novel flame retardants (N-FRs), polybrominated diphenyl ethers (PBDEs) and dioxin-like polychlorinated biphenyls (DL-PCBs) in fish, penguin, and skua from King George Island, Antarctica. Marine Pollution Bulletin, 2015, 96(1/2): 513–518. |