 2017, Vol. 37
2017, Vol. 37文章信息
- 岳楷, 杨万勤, 张川, 彭艳, 黄春萍, 吴福忠
- YUE Kai, YANG Wanqin, ZHANG Chuan, PENG Yan, HUANG Chunping, WU Fuzhong.
- 高寒森林不同生境对凋落叶分解灰分动态的影响
- Effects of environmental conditions on ash dynamics during foliar litter decomposition in an alpine forest
- 生态学报. 2017, 37(9): 3096-3105
- Acta Ecologica Sinica. 2017, 37(9): 3096-3105
- http://dx.doi.org/10.5846/stxb201601290213
-
文章历史
- 收稿日期: 2016-01-29
- 网络出版日期: 2016-12-19





2. 长江上游生态安全协同创新中心, 成都 611130;
3. 四川师范大学生命科学学院, 成都 610101
2. Collaborative Innovation Center for Ecological Security in the Upper Reaches of the Yangtze River, Chengdu 611130, China;
3. College of Life Science, Sichuan Normal University, Chengdu 610101, China
凋落叶是森林土壤和溪流生态系统中异养生物物质和能量的主要来源, 同时也是陆地生态系统物质循环和能量流动的一个重要环节[1]。灰分是凋落叶的重要组成部分, 其成分主要包括硅酸盐及钙 (Ca)、镁 (Mg)、钾 (K)、磷 (P) 等营养元素[1]。灰分动态不仅与凋落叶的分解过程相关, 且对研究凋落叶分解过程中有机物质的动态具有重要影响。传统上一般认为灰分浓度在凋落叶的分解过程中比较稳定或有少量增加, 因此采用凋落叶初始浓度值来计算凋落叶分解过程中无灰分质量损失 (ash free mass loss) 的方法被广泛用于凋落叶分解实验中用以排除外来物质对质量损失测定的干扰。然而, 有关凋落叶分解过程中灰分浓度稳定的报道主要来自于松属凋落叶, 且有报道表明阔叶树种凋落叶灰分浓度随凋落叶的分解而增加。糖枫 (sugar maple) 凋落叶经过1a的分解, 灰分浓度由最初的11.3%增加到19.5%, 且继续增加到分解第10年时的26.6%[1]。由此可见, 有关凋落叶分解过程中灰分浓度保持稳定的认识可能并不准确, 而导致无灰分质量损失的测定方法存在一定的不准确性。因此研究凋落叶分解过程中灰分动态特征显得十分必要。
森林生态系统中, 溪流在碳 (C) 和营养元素的跨生态系统运输过程中扮演了重要的角色[2-3]。凋落叶分解在水生态系统和陆地生态系统具有相似的影响因素, 包括气候条件、环境营养元素的有效性[4-5]、分解者的群落结构[6]以及凋落叶的质量 (quality) 特征[7]。环境条件 (如温度、湿度和营养有效性) 不仅能直接调控凋落叶的分解, 也可通过调控分解者的群落结构而间接影响凋落叶的分解过程[8]。然而, 由于这些影响因素在不同生境下差异很大, 因此在凋落叶分解过程中可能起到了不同的作用, 且对灰分的动态可能产生不同的影响。目前有关凋落叶分解过程的研究主要集中于单一的生态系统, 同时关注不同生境下凋落叶分解过程的研究相对较少, 而有关凋落叶分解过程中灰分浓度的动态过程的研究鲜有报道。高寒森林具有典型的季节动态, 冬季雪被的形成、覆盖、消融及伴随的土壤冻融循环过程显著地影响了环境特征, 且不同环境条件下存在较大的差异[9]。作为重要的水源涵养地, 岷江上游高寒森林生态系统的结构和功能直接关系到岷江流域乃至整个长江上游的生态安全[10]。因此, 以岷江上游高寒森林为研究区域, 选取森林地表、溪流和河岸带作为研究样地, 采用凋落物袋分解法研究了该区域典型植物康定柳 (Salix paraplesia)、高山杜鹃 (Rhododendron lapponicum)、方枝柏 (Sabina saltuaria) 及四川红杉 (Larix mastersiana) 凋落叶在不同生境下分解过程中灰分的动态特征。根据现有报道, 假设灰分浓度在凋落叶分解过程中呈现增加的趋势, 且受物种类型、分解时期和环境条件的调控。本研究以期为认识不同生境下凋落叶分解过程中灰分动态及目前广泛采用的无灰分质量损失方法提供一定的参考。
1 材料和方法 1.1 研究地概况本研究在位于四川省阿坝藏族羌族自治州理县的高山森林生态系统定位研究站 (102°53′—102°57′E, 31°14′—31°19′N, 2458—4619 m a.s.l.) 进行。该区域地处青藏高原东缘与四川盆地的过渡带, 是长江上游的典型地带。年平均气温约3℃, 最低温-18℃, 最高温23℃, 年平均降水量约850 mm。研究区的森林植被沿海拔由低到高依次为针阔混交林、高寒针叶林、高山灌丛和草甸。研究点位于海拔约3600 m的典型高寒森林。乔木层主要为四川红杉、方枝柏和岷江冷杉 (Abies faxoniana), 树龄约130a。林下灌丛主要为康定柳、高山杜鹃、三颗针 (Berberis sargentiana)、扁刺蔷薇 (Rose sweginzowii) 等, 草本植物主要有高山冷蕨 (Cystopteris montana)、薹草属 (Carex) 和莎草属 (Cyperus) 植物等。基于前期的研究和调查, 于研究区域选取3个25 m×25 m的样方 (31°14′ N, 102°53′ E, 3580 m a.s.l.), 每个样方相距约1 km左右。同时, 在邻近林下样地的区域分别选取3条典型的森林溪流和3个河岸带作为研究样方。
1.2 实验设计与样品处理2013年10月初在研究区域内收集当年自然凋落的康定柳、高山杜鹃、方枝柏和四川红杉凋落叶, 带回实验室自然风干。分别准确称取每种凋落叶风干样10 g各3份, 于65℃下烘干至恒重, 计算凋落叶初始含水量, 然后研磨过60目筛用以测定其初始化学计量 (表 1)。每个物种准确称取相当于烘干重10 g的风干样装入20 cm × 20 cm的尼龙网袋中 (孔径0.5 mm), 共1080袋 (4物种× 3样地× 3重复× 10次采样× 3生境)。2013年11月13日将装有样品的凋落物袋分别铺设于林下、溪流和河岸带的样地中。铺设时, 将凋落物袋系于安全绳后放置到溪流中, 并将安全绳一端固定于溪流岸边灌木的主干上, 每条溪流铺设3条系有样品的安全绳, 共120袋 (4物种× 3重复× 10次采样)。同样, 分别在林下和河岸带的每个样方内放置同样数量的凋落物袋。在每个处理的任一样方内随机选取一个凋落物袋, 各放置1个纽扣式温度计 (iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, USA), 同步监测不同环境条件下温度的动态特征。温度计读取时间设定为每2 h记录1次。
| 物种 Species |
全碳C/ (g/kg) |
全氮N/ (g/kg) |
全磷P/ (g/kg) |
碳氮比 C/N |
碳磷比 C/P |
氮磷比 N/P |
灰分Ash/% |
| 康定柳S. paraplesia | 337.59±0.71a | 26.69±1.08a | 1.75±0.03a | 12.68±0.51a | 192.89±3.88a | 15.23±0.47a | 10.83±0.21a |
| 高山杜鹃R. lapponicum | 382.11±11.07a | 6.59±0.86b | 0.97±0.11b | 60.44±9.74b | 403.01±35.70b | 7.13±1.46b | 4.33±0.14b |
| 方枝柏S. saltuaria | 462.99±14.99c | 10.42±0.51c | 1.54±0.03c | 44.79±3.82c | 301.35±4.59c | 6.80±0.44b | 4.58±0.09c |
| 四川红杉L. mastersiana | 377.94±1.05b | 15.70±1.01d | 1.25±0.02d | 24.28±1.61a | 302.85±5.19d | 12.58±0.87a | 7.33±0.09d |
| 同一列不同小写字母表示不同物种同一指标差异显著 (P < 0.05) | |||||||
根据前期研究及研究区气候状况, 以雪被形成、覆盖和融化特征为基础, 将1a划分为5个不同时期, 即冻结初期 (PP)、冻结期 (FP)、融化期 (TP)、生长季节 (GS) 和生长季节后期 (LGS)。埋样后, 分别于每个时期末进行采样, 每次于不同生境样地中采集不同物种凋落物袋各9袋 (3样地× 3重复), 同时读取温度数据, 并计算各个时期温度的平均温度、正积温和负积温 (表 2)。将采集的凋落叶自然风干后, 小心剔除凋落叶表面的泥沙和杂物, 于65℃下烘干后测定凋落叶的质量残留量。将称量后的凋落叶放入100 mL坩埚中, 置于550℃的马弗炉中5h使其充分燃烧。然后称量坩埚中剩下的部分, 即为灰分量 (g)。凋落叶初始全C含量采用氧化外加热法测定, 全氮 (N) 采用半微量凯氏定氮法测定, 全磷 (P) 采用钼锑抗比色法测定[11]。
| 时期 Period |
林下Forest floor | 溪流Stream | 河岸带Riparian zone | ||||||
| MT/℃ | PAT/℃ | NAT/℃ | MT/℃ | PAT/℃ | NAT/℃ | MT/℃ | PAT/℃ | NAT/℃ | |
| PF1 | -4.3 | 0.4 | -177.8 | 4.1 | 167.0 | 0 | 0.8 | 36.2 | -1.7 |
| FP1 | -5.01 | 2.71 | -388.6 | 3.4 | 258.7 | 0 | 0.8 | 62.6 | 0 |
| TP1 | 2.1 | 110.8 | -15.8 | 3.8 | 170.9 | 0 | 5.3 | 236.6 | 0 |
| GS1 | 8.9 | 911.3 | 0 | 5 | 510.0 | 0 | 9.0 | 914.3 | 0 |
| LGS1 | -6.5 | 653.9 | -1.8 | 6.1 | 618.7 | 0 | 8.1 | 816.8 | 0 |
| PF2 | -1.9 | 0.1 | -65.3 | 4.0 | 170.5 | 0 | 1.6 | 55.3 | 0 |
| FP2 | -3.4 | 12.9 | -298.6 | 2.9 | 239.1 | 0 | 0.7 | 59.8 | 0 |
| TP2 | 1.9 | 85.9 | -3.9 | 3.9 | 170.5 | 0 | 5.0 | 220.6 | 0 |
| GS2 | 9.1 | 941.4 | 0 | 5.1 | 519.7 | 0 | 9.0 | 925.3 | 0 |
| LGS2 | 6.3 | 620.0 | 0 | 14.4 | 591.1 | 0 | 7.8 | 774.3 | 0 |
| MT:平均温度; PAT:正积温; NAT:负积温; PP:冻结初期; FP:冻结期; TP:融化期; GS:生长季节; LGS:生长季节后期; 1:第1年; 2:第2年 | |||||||||
每次采样即时测定溪流与河岸带水体的pH (pH 320, WTW GmbH, Weilheim, Germany) 和流速 (flow velocity; Martin Marten Z30, Current Meter)。同时, 用聚乙烯塑料瓶采集溪流和河岸带每个样方水样各1份, 带回实验室后及时用0.7 μm孔径的玻璃纤滤器 (GF/F glass fibre filter; Whatman International, Florham Park, NJ, USA) 过滤备用。采用双指示剂中和法测定碳酸氢根 (HCO3-) 的含量, 紫外分光光度计 (UV-2600, Unico Instruments Co., Ltd., Shanghai, China) 测定磷酸根 (PO43-) 的含量, 离子色谱 (ICS-90, Dionex Corp., Sunnyvale, CA, YSA) 测定铵态氮 (NH4+-N) 和硝态氮 (NO3--N) 的含量 (表 3)[12]。
| 时期Period | 流速 Flow velocity (m/s) |
pH | HCO3-/ (mg/L) |
PO43-/ (μg/L) |
NH4+-N/ (mg/L) |
NO3--N/ (mg/L) |
|
| 溪流Stream | |||||||
| PF1 | 0.59±0.03 | 6.85±0.04 | 15.45±0.33 | 7.8±0.20 | 0.13±0.01 | 0.32±0.03 | |
| FP1 | 0.30±0.02 | 6.68±0.06 | 10.71±0.08 | 7.2±0.15 | 0.01±0.01 | 0.27±0.02 | |
| TP1 | 0.72±0.02 | 6.81±0.02 | 14.53±0.31 | 8.3±0.10 | 0.13±0.03 | 0.31±0.03 | |
| GS1 | 0.72±0.04 | 5.44±0.24 | 13.87±0.09 | 8.1±0.10 | 0.07±0.01 | 0.24±0.03 | |
| LGS1 | 0.61±0.03 | 6.67±0.02 | 15.50±0.16 | 7.8±0.10 | 0.15±0.02 | 0.35±0.03 | |
| PF2 | 0.54±0.02 | 6.56±0.08 | 15.11±0.05 | 7.7±0.04 | 0.08±0.01 | 0.17±0.01 | |
| FP2 | 0.29±0.01 | 6.69±0.04 | 10.93±0.15 | 7.4±0.09 | 0.05±0.01 | 0.26±0.03 | |
| TP2 | 0.65±0.04 | 6.70±0.02 | 14.61±0.10 | 8.1±0.03 | 0.09±0.01 | 0.22±0.02 | |
| GS2 | 0.72±0.02 | 6.54±0.02 | 14.24±0.03 | 8.1±0.03 | 0.08±0.00 | 0.24±0.01 | |
| LGS2 | 0.51±0.03 | 6.75±0.02 | 14.48±0.15 | 7.8±0.14 | 0.16±0.02 | 0.36±0.03 | |
| 河岸带Riparian zone | |||||||
| PF1 | 0.05 ± 0.00 | 6.74 ± 0.07 | 21.05 ± 0.11 | 7.9 ± 0.31 | 0.02 ± 0.01 | 0.31 ± 0.01 | |
| FP1 | 0.04 ± 0.00 | 7.13 ± 0.04 | 21.06 ± 0.09 | 7.4 ± 0.20 | 0.01 ± 0.00 | 0.40 ± 0.02 | |
| TP1 | 0.05 ± 0.01 | 6.92 ± 0.05 | 18.41 ± 0.21 | 8.1 ± 0.12 | 0.04 ± 0.01 | 0.36 ± 0.04 | |
| GS1 | 0.06 ± 0.01 | 6.47 ± 0.14 | 18.57 ± 0.33 | 8.2 ± 0.15 | 0.04 ± 0.01 | 0.24 ± 0.03 | |
| LGS1 | 0.05 ± 0.00 | 6.70 ± 0.05 | 19.53 ± 0.03 | 7.4 ± 0.06 | 0.05 ± 0.01 | 0.35 ± 0.03 | |
| PF2 | 0.05 ± 0.00 | 7.08 ± 0.08 | 19.23 ± 0.02 | 7.6 ± 0.04 | 0.03 ± 0.00 | 0.35 ± 0.01 | |
| FP2 | 0.04 ± 0.00 | 7.14 ± 0.02 | 21.05 ± 0.09 | 7.4 ± 0.05 | 0.02 ± 0.01 | 0.42 ± 0.03 | |
| TP2 | 0.05 ± 0.01 | 7.03 ± 0.02 | 19.06 ± 0.05 | 7.6 ± 0.04 | 0.03 ± 0.01 | 0.26 ± 0.02 | |
| GS2 | 0.06 ± 0.01 | 6.80 ± 0.03 | 20.12 ± 0.04 | 8.1 ± 0.03 | 0.06 ± 0.00 | 0.30 ± 0.05 | |
| LGS2 | 0.05 ± 0.00 | 6.71 ± 0.03 | 19.48 ± 0.03 | 7.5 ± 0.05 | 0.05 ± 0.00 | 0.35 ± 0.04 | |
| PP:冻结初期pre-freezing period; FP:冻结期freezing period; TP:融化期thawing period; GS:生长季节growing season; LGS:生长季节后期late growing season; 1:第1年; 2:第2年 | |||||||
不同时期凋落叶灰分浓度Ct (%) = Mt/M′t × 100%, 式中Mt为第t次采样灰分的残留量, M′t为第t次采样凋落叶的残留量 (t = 0, 1, 2, 4, 5, 6, 7, 8, 9, 10);凋落叶灰分浓度在各个时期的变异量Ct (%) = (Ct-Ct-1)/C0 × 100%, 式中Ct-1和Ct分别为第t-1和第t次采样凋落叶灰分的浓度 (t = 1, 2, 3, 4, 5, 6, 7, 8, 9, 10), C0为凋落叶灰分的初始浓度;凋落叶灰分浓度在整个2年的变异量C (%) = (C10-C0)/C0 × 100%, 式中C10为第10次采样凋落叶灰分的浓度。
采用单因素方差分析 (one-way ANOVA) 比较不同物种初始质量指标的差异性, 同一物种在相同环境条件下不同分解时期灰分量和浓度的差异性;采用双因素方差分析 (two-way ANOVA) 检验分解时期和物种对凋落叶灰分量的影响;采用逐步回归分析 (stepwise regression analysis) 来检验凋落叶初始质量对凋落叶灰分变异量的影响;采用Pearson相关分析检验环境因子对凋落叶灰分浓度变异量的相关性。数据统计和分析采用SPSS 19.0(SPSS Inc., Chicago, IL, USA) 完成。
2 结果 2.1 凋落叶灰分量的动态特征随着凋落叶的分解, 不同生境下不同物种凋落叶中灰分量整体呈现了下降的趋势, 但不同生境、物种和分解时期之间存在一定的差异 (图 1)。就生境而言, 各个物种的灰分量在溪流中减少最快, 河岸带次之, 林下最慢。就物种而言, 康定柳凋落叶灰分量减少最多, 经过两年的分解由最初的1.08 g减少到0.12—0.39 g (图 1)。其次为四川红杉和方枝柏凋落叶, 而高山杜鹃凋落叶灰分量减少相对缓慢 (图 1)。在不同分解时期, 凋落叶灰分量存在显著 (P < 0.05) 差异。康定柳凋落叶灰分量在第一年的生长季节减少最为显著, 而高山杜鹃凋落叶灰分量仅在溪流中的不同时期差异较为显著。对方枝柏和四川红杉凋落叶而言, 灰分量虽然在各个时期存在差异, 但变异在凋落叶的分解过程中相对较小 (图 1, 图 2)。

|
| 图 1 不同生境下康定柳, 高山杜鹃, 方枝柏, 四川红杉凋落叶分解过程中灰分量的动态特征 (平均值±标准偏差, n=9) Fig. 1 Dynamics of ash content in decomposing litters of Salix paraplesia, Rhododendron lapponicum, Sabina saltuaria, and Larix mastersiana under different environmental conditions (mean ± SD, n=9) Ⅳ:初始值; PP:冻结初期; FP:冻结期; TP:融化期; GS:生长季节; LGS:生长季节后期; 1:第1年; 2:第2年; 不同小写字母表示同一物种在同一环境条件下的不同分解时期灰分量差异显著 (P < 0.05) |

|
| 图 2 不同生境下康定柳, 高山杜鹃, 方枝柏, 四川红杉凋落叶分解过程中灰分浓度的动态特征 (平均值±标准偏差, n=9) Fig. 2 Dynamics of ash concentration in decomposing litters of Salix paraplesia, Rhododendron lapponicum, Sabina saltuaria, and Larix mastersiana under different environmental conditions (mean ± SD, n=9) Ⅳ:初始值; PP:冻结初期; FP:冻结期; TP:融化期; GS:生长季节; LGS:生长季节后期; 1:第一年; 2:第二年; 不同小写字母表示同一物种在同一环境条件下的不同分解时期灰分量差异显著 (P < 0.05) |
相对凋落叶灰分量而言, 灰分浓度在凋落叶分解过程中表现出了缓慢增加的趋势 (图 2)。灰分浓度随物种的不同差异很大, 表现为康定柳>四川红杉>方枝柏>高山杜鹃。同一物种凋落叶灰分浓度在不同环境条件下差异不明显, 但在不同分解时期其浓度在一定程度上存在显著差异 (P < 0.05)。在康定柳凋落叶分解初期, 灰分浓度呈现出明显的上升, 之后表现出先下降再上升的规律 (图 2)。经过两年的分解, 康定柳凋落叶灰分浓度与初始浓度差异不显著 (P > 0.05)。对于其他3种凋落叶灰分浓度而言, 虽然在凋落叶分解的过程中存在波动, 但整体呈现出缓慢上升的趋势 (图 2)。
2.3 凋落叶灰分浓度变异量及其影响因素经过2a的分解, 4种凋落叶中灰分浓度整体表现为增加, 但物种之间存在明显差异, 表现为方枝柏>高山杜鹃>四川红杉>康定柳 (图 3)。双因素方差分析表明物种和分解时期及其交互作用显著影响了凋落叶分解过程中灰分浓度的变异量 (表 4)。同一环境条件下, 不同物种凋落叶灰分浓度变异量同样表现出了明显的差异 (图 3)。逐步回归分析表明, 凋落叶初始质量能显著影响其分解过程中灰分浓度的动态, 且主要与初始C含量和N/P相关, 其次与N含量显著相关 (表 5)。相关分析表明, 凋落叶分解过程中灰分变异量同样与环境因子存在紧密的关系, 但不同环境条件和不同物种之间存在一定的差异 (表 6)。

|
| 图 3 不同生境下康定柳, 高山杜鹃, 方枝柏, 四川红杉凋落叶分解过程中灰分浓度的变异量 (平均值±标准偏差, n=9) Fig. 3 Changes of ash concentration in decomposing litters of Salix paraplesia, Rhododendron lapponicum, Sabina saltuaria, and Larix mastersiana under different environmental conditions (mean ± SD, n=9) Ⅳ:初始值; PP:冻结初期; FP:冻结期; TP:融化期; GS:生长季节; LGS:生长季节后期; 1:第1年; 2:第2年; 不同小写字母表示同一物种在同一环境条件下的不同分解时期灰分量差异显著 (P < 0.05) |
| 因素Factor | 林下Forest floor | 溪流Stream | 河岸带Riparian zone | |||
| df | F | df | F | df | F | |
| 第1年Year 1 | ||||||
| 时期Period | 4 | 0.88 | 4 | 3.15* | 4 | 4.43** |
| 物种Species | 3 | 4.58** | 3 | 2.71* | 3 | 4.83** |
| 时期×物种Period × Species | 12 | 22.54*** | 12 | 4.88*** | 12 | 13.19*** |
| 第2年Year 2 | ||||||
| 时期Period | 4 | 15.76*** | 4 | 12.14*** | 4 | 7.05*** |
| 物种Species | 3 | 1.59 | 3 | 19.65*** | 3 | 11.65*** |
| 时期×物种Period × Species | 12 | 17.85*** | 12 | 18.89*** | 12 | 5.70*** |
| 全两年2 years | ||||||
| 时期Period | 9 | 9.49*** | 9 | 5.83*** | 9 | 5.44*** |
| 物种Species | 3 | 3.80* | 3 | 7.07*** | 3 | 7.94*** |
| 时期×物种Period × Species | 27 | 17.45*** | 27 | 8.35*** | 27 | 9.11*** |
| df自由度degree of freedom; *P < 0.05, **P < 0.01, ***P < 0.001 | ||||||
| 环境条件Environmental condition | 回归式Regression model | ||||
| a0 | a1X1 | a1X1 | a3X3 | ||
| 林下Forest floor | 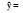 |
92.23 | -4.75 N/P (0.761) | -0.07 C/P (0.806) | |
| 溪流Stream | 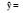 |
-68.70 | +0.36 C (0.869) | -7.73 N/P (0.893) | +2.41 N (0.933) |
| 河岸带Riparian zone | 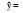 |
40.69 | +0.17 C (0.675) | -12.28 N/P (0.749) | +3.59 N (0.827) |
| C:全碳; N:全氮; C/P:碳磷比; N/P:氮磷比; 括号数字表示决定系数 (R2), n=36 | |||||
| 物种Species | MT | PAT | NAT | FV | pH | HCO3- | PO43- | NH4+-N | NO3--N |
| 林下Forest floor | |||||||||
| 康定柳S. paraplesia | -0.251* | 0.028 | 0.116 | ||||||
| 高山杜鹃R. lapponicum | -0.003 | -0.071 | 0.161 | ||||||
| 方枝柏S. saltuaria | 0.083 | 0.101 | 0.257* | ||||||
| 四川红杉L. mastersiana | 0.206 | 0.025 | -0.081 | ||||||
| 溪流Stream | |||||||||
| 康定柳S. paraplesia | 0.050 | -0.233* | -0.093 | 0.127 | 0.065 | -0.181 | 0.226* | -0.095 | 0.006 |
| 高山杜鹃R. lapponicum | -0.067 | -0.062 | 0.135 | 0.298** | 0.167 | 0.317** | 0.258* | 0.289** | 0.175 |
| 方枝柏S. saltuaria | 0.127 | 0.249* | -0.003 | -0.067 | 0.094 | 0.162 | -0.099 | 0.237* | 0.345** |
| 四川红杉L. mastersiana | -0.010 | -0.071 | -0.204 | -0.227* | -0.171 | -0.309** | -0.199 | -0.263* | -0.048 |
| 河岸带Riparian zone | |||||||||
| 康定柳S. paraplesia | -0.257* | -0.124 | 0.074 | -0.419** | 0.256* | 0.382** | -0.436** | -0.151 | 0.521** |
| 高山杜鹃R. lapponicum | 0.329** | 0.241* | 0.096 | 0.152 | -0.265* | -0.320** | 0.223* | 0.201 | -0.070 |
| 方枝柏S. saltuaria | 0.051 | 0.172 | -0.149 | -0.074 | -0.029 | 0.162** | -0.184 | 0.182 | 0.129 |
| 四川红杉L. mastersiana | -0.415** | -0.363** | -0.565** | -0.210* | -0.026 | 0.307* | 0.066 | -0.390** | 0.078 |
| MT:平均温度mean temperature;PAT:正积温positive accumulative temperature;NAT:负积温negative accumulative temperature;FV:流速flow velocity; n= 90, *P < 0.05, **P < 0.01 | |||||||||
与提出的科学假设一致, 本研究结果表明凋落叶在2a的分解过程中灰分量在不同环境条件下整体上均呈现了下降的趋势, 但不同物种之间存在明显的差异 (图 1)。相反, 凋落叶中灰分浓度在不同环境条件下整体表现为增加的趋势 (图 2), 但浓度变异量在不同时期差异明显, 且受到物种和环境因子的调控 (图 3, 表 4—表 6)。这一方面证明凋落叶分解过程中灰分浓度并不一定保持稳定[1], 另一方面也表明高寒森林季节性冻融循环对凋落叶分解过程中灰分浓度动态具有不同程度的影响, 且物种差异所代表的凋落叶质量对分解过程中灰分浓度变异量具有重要的影响。
凋落叶分解过程中, 随着有机物质的降解和凋落叶的失重, 各种元素也随之被释放出来, 且灰分量也随之减少。当凋落叶中某一元素或组分的释放速率低于凋落叶的失重率时, 即出现浓度上升的现象[13]。影响凋落叶分解过程中元素或组分动态的因素十分复杂, 其中以凋落叶质量特征、外界环境条件和元素或组分的本身性质为主[14]。本研究发现, 凋落叶分解过程中灰分浓度同样受到这些因素的调控。灰分一般被定义为凋落叶充分燃烧后剩下的组分, 即不包含有机C、有机N、可燃性组分和挥发性组分等[1]。近期研究表明, 由物种决定的基质质量是影响凋落叶分解过程的主导因素[7, 15], 而本研究发现凋落叶初始质量同样是分解过程中灰分动态的重要影响因子, 且主要受凋落叶初始C、N含量和N/P比的影响 (表 4, 表 5)。不同物种凋落叶不仅初始灰分浓度差异显著 (表 1), 且在同一生境下不同物种凋落叶灰分浓度在相同时期变异量差异明显 (图 3)。凋落叶初始质量不仅直接与分解过程中各组分动态相关, 同时影响其在分解过程中消费者和分解者的群落结构, 进而间接影响其分解过程和个组分的动态特征[16]。然而值得注意的是, 灰分量在某些分解时期出现了增加的现象。这可能是因为凋落叶分解过程中有新的灰分生成导致, 因为凋落叶的分解过程伴随着有机物转化为无机物的转化过程。当凋落叶中灰分的形成速率大于流失速率时, 灰分量即表现为增加。
前期研究表明, 高寒森林不同冻融循环时期显著影响了凋落叶的分解过程[17-18]。本研究发现, 凋落叶分解过程中灰分浓度在不同时期同样存在明显或显著的差异 (图 3, 表 4), 即环境条件显著影响了灰分的动态特征。对同一物种而言, 除溪流中康定柳凋落叶外, 虽然不同环境条件下凋落叶在整个分解过程中灰分浓度均表现为增加, 但在不同分解时期存在明显的差异 (图 3)。高寒森林具有典型的冻融循环现象, 不同分解时期环境因子存在明显的差异 (表 2, 表 3)。温度是影响凋落叶失重和元素释放过程的一个重要环境因子, 不同生境下由温度导致的诸如营养元素有效性和微生物群落结构及功能的差异也是影响凋落叶分解过程的一个主要因素[19]。由相关分析可知, 凋落叶分解过程中灰分浓度动态在一定程度上不仅与温度直接相关, 而且与其分解环境中营养元素有效性等因子显著相关 (表 6)。这些结果表明, 与凋落叶分解过程中其它组分或元素一样, 灰分浓度在凋落叶的分解过程中同样受到环境因子的影响。
综上所述, 凋落叶分解过程中, 灰分量随凋落叶的失重而呈现逐渐减少的趋势, 且物种之间存在明显的差异。与之相反, 经过2a的分解, 除溪流中康定柳凋落叶灰分浓度略有下降外 (-0.99%), 林下和河岸带中康定柳凋落叶及其它物种凋落叶中灰分浓度均有不同程度的增加 (5.86%—72.15%)。整体而言, 凋落叶灰分浓度在不同环境条件下表现为增加的趋势, 且浓度的变异量受凋落叶初始质量特征和环境因子等因素的调控。这表明, 虽然不同物种和不同生境能显著的影响凋落叶分解过程中灰分浓度的动态特征, 但灰分浓度整体上呈现增加的趋势是一致的。因此, 传统上认为凋落叶分解过程中灰分浓度基本保持不变的观点并不准确, 且其浓度受凋落叶物种和分解生境的影响。此外, 本研究结果表明以凋落物灰分浓度不变的前提采用初始灰分浓度值计算凋落物分解过程中无灰分质量损失的方法并不适用于所有物种的凋落叶, 计算过程中应该慎用。本研究为认识凋落叶分解过程中灰分动态特征提供了一定的参考, 同时为凋落叶分解过程中质量损失计算方法的选择提供了一定的数据支持。
| [1] | Berg B, McClaugherty C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration. Berlin Heidelberg: Springer, 2014. |
| [2] | Gessner MO, Chauvet E, Dobson M. A perspective on leaf litter breakdown in streams. Oikos, 1999, 85(2): 377–384. DOI:10.2307/3546505 |
| [3] | Wallace JB, Eggert S L, Meyer JL, Webster J R. Effects of resource limitation on a detrital-based ecosystem. Ecological Monographs, 1999, 69(4): 409–442. DOI:10.1890/0012-9615(1999)069[0409:EORLOA]2.0.CO;2 |
| [4] | Boyero L, Pearson RG, Gessner MO, Barmuta LA, Ferreira V, GraçaM A S, Dudgeon D, Boulton A J, Callisto M, Chauvet E, Helson J E, Bruder A, Albariño R J, Yule C M, Arunachalam M, Davies J N, Figueroa R, Flecker A S, Ramírez A, Death R G, Iwata T, Mathooko J M, Mathuriau C, Gonçalves Jr J F, Moretti M S, Jinggut T, Lamothe S, M'Erimba C, Ratnarajah L, Schindler M H, Castela J, Buria L M, Cornejo A, Villanueva V D, WestD C. A global experiment suggests climate warming will not accelerate litter decomposition in streams but might reduce carbon sequestration. Ecology Letters, 2011, 14(3): 289–294. DOI:10.1111/ele.2011.14.issue-3 |
| [5] | García-Palacios P, Maestre FT, Kattge J, Wall DH. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecology Letters, 2013, 16(8): 1045–1053. DOI:10.1111/ele.2013.16.issue-8 |
| [6] | McKie BG, Petrin Z, Malmqvist B. Mitigation or disturbance? Effects of liming on macroinvertebrate assemblage structure and leaf-litter decomposition in the humic streams of northern Sweden. Journal of Applied Ecology, 2006, 43(4): 780–791. DOI:10.1111/j.1365-2664.2006.01196.x |
| [7] | Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie S E, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested H M, Santiago L S, Wardle D A, Wright I J, Aerts R, Allison S D, Van Bodegom P, Brovkin V, Chatain A, Callaghan T V, Díaz S, Garnier E, Gurvich D E, Kazakou E, Klein J A, Read J, Reich P B, Soudzilovskaia N A, Vaieretti M V, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letters, 2008, 11(10): 1065–1071. DOI:10.1111/ele.2008.11.issue-10 |
| [8] | Waldrop MP, Zak DR, Sinsabaugh RL. Microbial community response to nitrogen deposition in northern forest ecosystems. Soil Biology and Biochemistry, 2004, 36(9): 1443–1451. DOI:10.1016/j.soilbio.2004.04.023 |
| [9] | 张川, 杨万勤, 岳楷, 黄春萍, 彭艳, 吴福忠. 高山森林溪流冬季不同时期凋落物分解中水溶性氮和磷的动态特征. 应用生态学报, 2015, 26(6): 1601–1608. |
| [10] | Zhang M F, Wei X H, Sun P S, Liu S R. The effect of forest harvesting and climatic variability on runoff in a large watershed: the case study in the upper Minjiang River of Yangtze River basin. Journal of Hydrology, 2012, 464-465: 1–11. DOI:10.1016/j.jhydrol.2012.05.050 |
| [11] | 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999. |
| [12] | American Public Health Association (Author), AWWA (American Water Works Association) (Author), Water Environment Federation. Standard Methods for the Examination of Water and Wastewater. 22nd ed. Washington, D.C.: American Public Health Association, 2012. |
| [13] | 何洁, 杨万勤, 倪祥银, 李晗, 徐李亚, 吴福忠. 雪被斑块对川西亚高山森林凋落物冬季分解过程中钾和钠动态的影响. 植物生态学报, 2014, 38(6): 550–561. |
| [14] | Berg B, Kjønaas O J, Johansson MB, Erhagen B, Åkerblom S. Late stage pine litter decomposition: relationship to litter N, Mn, and acid unhydrolyzable residue (AUR) concentrations and climatic factors. Forest Ecology and Management, 2015, 358: 41–47. DOI:10.1016/j.foreco.2015.08.032 |
| [15] | Makkonen M, Berg MP, Handa IT, Hättenschwiler S, Van Ruijven J, Van Bodegom P M, AertsR. Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecology Letters, 2012, 15(9): 1033–1041. DOI:10.1111/j.1461-0248.2012.01826.x |
| [16] | Gessner MO, Swan CM, Dang CK, McKie BG, Bardgett RD, Wall D H, Hättenschwiler S. Diversity meets decomposition. Trends in Ecology & Evolution, 2010, 25(6): 372–380. |
| [17] | 何伟, 吴福忠, 杨万勤, 武启骞, 何敏, 赵野逸. 雪被斑块对高山森林两种灌木凋落叶质量损失的影响. 植物生态学报, 2013, 37(4): 306–316. |
| [18] | 岳楷, 杨万勤, 彭艳, 张川, 黄春萍, 吴福忠. 岷江上游高山森林凋落叶在冬季河流中的质量损失特征. 长江流域资源与环境, 2015, 24(7): 1177–1184. |
| [19] | García-Palacios P, McKie BG, Handa IT, Frainer A, Hättenschwiler S. The importance of litter traits and decomposers for litter decomposition: a comparison of aquatic and terrestrial ecosystems within and across biomes. Functional Ecology, 2016, 30(5): 819–829. DOI:10.1111/1365-2435.12589 |