 2017, Vol. 37
2017, Vol. 37文章信息
- 梅军林, 庄枫红, 马姜明, 覃扬浍, 梁士楚, 姜勇
- MEI Junlin, ZHUANG Fenghong, MA Jiangming, QIN Yanghui, LIANG Shichu, JIANG Yong.
- 桂林喀斯特地区克隆生长红背山麻杆种群的点格局分析
- Spatial point pattern analysis of Alchornea trewioides population clonal growth in the karst area of Guilin
- 生态学报. 2017, 37(9): 3164-3171
- Acta Ecologica Sinica. 2017, 37(9): 3164-3171
- http://dx.doi.org/10.5846/stxb201601240162
-
文章历史
- 收稿日期: 2016-01-24
- 网络出版日期: 2016-12-19





2. 珍稀濒危动植物生态与环境保护教育部重点实验室, 桂林 541004;
3. 岩溶生态与环境变化研究广西高校重点实验室, 桂林 541004;
4. 广西珍稀濒危动物生态学重点实验室, 桂林 541004
2. Key Laboratory of Ecology of Rare and Endangered Species and Environmental Protection, Ministry of Education, China 541004, China;
3. University Key Laboratory of Karst Ecology and Environmental Change of Guangxi Province (Guangxi Normal University), Guilin 541004, China;
4. Guangxi Key Laboratory of Rare and Endangered Animal Ecology, GuangxiNormalUniversity, Guilin 541004, China
种群空间分布格局能在很大程度上反映该种群与生境的关系及其在群落中的地位和作用[1]。它与尺度有很大的关系, 这个问题早就受到许多学者的关注, 传统的分析方法在特定空间尺度上得到的认识不能推广或者应用到其它尺度上去[2]。点格局分析方法是由Ripiey[3]首先提出, 后经Diggle等的发展逐步形成一种种群格局分析的新方法, 能够最大限度地利用空间点信息来分析植物种群由小到大一系列连续尺度上的空间分布格局[4]。它是以植物种的个体在空间的坐标为基本数据, 每个个体都可以视为二维空间的一个点, 这样所有个体就组成了空间分布的点图, 以点图为基础进行格局分析, 它可以分析各种尺度下的种群格局和种间关系, 在拟合分析的过程中最大限度的利用了坐标图的信息, 因而检验能力较强[5]。开展种群空间分布格局的研究对于了解物种的特性, 种群间相互关系以及种群与环境之间的关系具有重要意义, 是研究植物群落空间结构的基本组成要素[6]。目前, 关于种群空间格局的研究主要包括不同的尺度, 如个体生长状况所造成的分布格局、植物种群对环境变化直接反应而产生的格局、植物间的相互关系或植物和动物间相互关系而产生的群落格局、大的植被类型以及分类单位在全球范围内的景观格局[7]。一般来说, 植物种群在群落中的分布有3种类型, 即随机分布、集群分布和分散分布。空间关联性为群落的重要特征之一, 是群落形成、演化的基础和重要的数量、结构指标, 也是种间或种内关系的一种表现形式和群落分类的依据[8-9]。对群落关联性分析是研究群落内部镶嵌结构的重要方法, 因为一个种斑块的形成、变化都影响着整个群落结构的变化[10]。而种内联结用来表示同一物种不同龄级在空间分布上的相互关联性, 是对一定时期内植物种群组成个体之间相互关系的静态描述, 这种关系不仅包括空间分布关系, 也包含着个体间的功能关系, 揭示了种群的现状, 并能反映种群内个体间的相互作用[11-12]。
红背山麻杆 (Alchornea trewioides (Benth.) Muell. Arg.) 又名红背娘、红背叶, 属于大戟科 (Euphorbiaceae) 山麻杆属 (Alchornea Sw.) 植物, 为桂林喀斯特地区常见的先锋灌木树种, 通常在一些皆伐地段或者废弃的采石场等开阔地段出现, 能以克隆繁殖为主兼具有性繁殖进行快速覆盖裸露岩石并形成红背山麻杆群落。近年来, 对红背山麻杆的生物量[13]、水分生理特征、光合作用[14]、红背山麻杆叶的化学成分研究[15-18], 以及红背山麻杆在喀斯特地区中元素迁移的作用[19]等方面进行了研究。但对于以克隆繁殖为主红背山麻杆种群的空间分布格局尚未见报道, 尤其是对红背山麻杆各个径级在空间上的相互关系等方面缺乏充分的认识。研究喀斯特地区先锋物种红背山麻杆种群空间格局及其不同径级之间的关联, 有助于深化对喀斯特地区克隆植物种群组成、结构与动态的认识, 对喀斯特地区石漠化的重建与恢复具有重要意义。
1 研究区概况与方法 1.1 研究区概况研究样地设在桂林境内的典型喀斯特石山, 地理坐标为109°45′—104°40′ E, 24°18′—25°41′ N。桂林地处广西东北部, 平均海拔160 m, 属于中亚热带湿润季风气候, 气候温和, 雨量充沛, 年平均气温18.9 ℃。年均日照总时数1553 h, 全年无霜期300 d, 年均降雨量1949.5 mm, 降雨量分配不均, 秋、冬季干燥少雨;年平均蒸发量1490—1905 m。研究样地内植被因人为干扰程度和地形因子的复合影响而发生次生演替, 现存的红背山麻杆群落中其他伴生种主要有紫薇 (Lagerstroemia indica)、龙须藤 (Bauhinia championii) 和黄荆 (Vitex negundo) 等。
1.2 调查方法2014年7月, 在红背山麻杆群落样地内选择有代表性的6个样方, 通常灌木样方为5 m×5 m, 考虑到做点格局分析及岩溶石山地势条件的特殊性, 将样方适当扩大为10 m×10 m, 并以此面积进行点格局分析。调查时将每个样方划分为4个5 m×5 m的小样方, 采用每木调查法, 测量样地内所有体植株地径、坐标、株高、冠幅等数量特征。同时记录群落所处海拔、盖度、岩石裸露率、坡度、坡向、坡位、郁闭度等群落综合特征和生态学指标。本研究依据样方内红背山麻杆整体地径大小情况, 将红背山麻杆地径划分为径级Ⅰ(地径≤0.5 cm)、径级Ⅱ(0.5 cm<地径≤1.0 cm)、径级Ⅲ(地径>1.0 cm)3个不同径级。运用Ripley′s K函数对6个样方红背山麻杆进行分布格局及3个径级之间的空间关联性进行研究。
1.3 数据分析 1.3.1 空间格局分析本研究主要以Ripley′s K函数为基础, 应用L(r) 和g(r) 函数两种方法, 分别以样方中每个红背山麻杆个体的空间分布坐标点图为基础, 分析红背山麻杆在不同尺度范围内的格局。其中, K函数为:

|
(1) |
式中, r表示空间尺度, A为样方面积, n为样方内个体数, wij为边界效应修正, uij指点i和点j间的距离, I为指示函数。当uij≤r时, I=1.0, 当uij>r时, I=0。为了用一种更为直接而简捷的函数反映空间格局, Besag[20]提出用L(r) 取代K(r), 并对K(r) 作开平方的线性变换, 以保持方差稳定。L(r) 的表达式为:
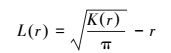
|
(2) |
L(r)=0说明个体为完全空间随机分布, L(r)>0为聚集分布, L(r)<0为分散分布。
二阶函数分析 (即Ripley′s K函数) 是较为常用和有效的空间点格局分析方法, 然而它是一个累积分布函数, 其在大尺度上的格局分析受到小尺度上累积效应的影响[21-22], 从而使格局分析复杂化[23]。g(r) 函数是从K函数的基础上衍生出来的, 主要以环代替K函数中的圆, 计算过程没有累积效应[24]。它是利用点间的距离, 计算任一点为圆心、半径为r、指定宽度的圆环区域内的点的数量来进行空间点格局分析[25], 它与K函数之间存在着函数关系, 即:
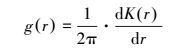
|
(3) |
式中, 当g(r)=1.0时, 个体分布显示为完全空间随机分布, g(r)>1.0时为聚集分布, g(r)<1.0时为分散分布。鉴于g(r) 函数能有效剔除K函数小尺度上的累积效应、更直观、更准确[26]。本文使用该函数描述具体空间尺度上的种群分布格局。
1.3.2 不同径级红背山麻杆空间关联性分析分析两个不同物种之间空间关系的点格局分析被称为二元点格局分析。本研究以K函数为基础, 利用两个不同物种的点格局分析方法来反映同一物种不同径级间的空间关系。分析不同径级红背山麻杆植株之间的空间关联:
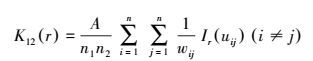
|
(4) |
式中, n1、n2表示两个不同径级红背山麻杆的植株数量;i和j分别代表两个不同径级的植株。同样, 对K(r) 进行平方根转换, 得到L函数[27], 即:
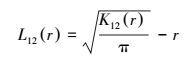
|
(5) |
在分析植物竞争时, 选择L(r) 函数能够更好地表现生态过程[28]。在L12(r)>1.0, 则两个径级在距离r处显著正相关;L12(r)<1.0, 则两个径级在距离r处显著负相关;若L12(r)=1.0, 则表明两个径级之间相互独立或没有显著相关性。用Monte-Carlo拟合, 确定上下包迹线, 即置信区间的范围。本文拟合次数为100次, 取95%的置信水平。
本文空间点格局分析以及关联性统计分析均采用R 3.2.2软件中的spatstat程序包完成。若实际分布的g(r) 值落在包迹线内, 则红背山麻杆符合随机分布;若在包迹线以上, 则红背山麻杆为聚集分布;若落在包迹线以下, 则呈分散分布。若L12(r) 值落在包迹线内, 不同径级间相互独立, 无空间相关性;若在包迹线以上, 两径级间为空间正相关, 说明两个龄级之间对环境资源需求的相似性或互补性;若落在包迹线以下, 两径级间为空间负相关, 说明两个龄级之间相互排斥或者对环境需求的竞争性[29]
2 结果与分析 2.1 径级构成6个样方中红背山麻杆远远多于其他物种, 虽然6个样方中红背山麻杆个体密度差别较大, 但基本上都是呈聚集分布 (图 1)。6个样方中红背山麻杆3个径级所占比例差距较大, 均是Ⅰ径级>Ⅱ径级>Ⅲ径级 (表 1)。Ⅰ径级株数在D样方中最多, 在A样方中最少;Ⅱ径级株数在E样方中最多, 在B样方中最少;Ⅲ径级株数在A样方中最多, 在F样方中最少 (表 1)。

|
| 图 1 红背山麻杆点分布图 Fig. 1 Point patterns of Alchornea trewioides population A~F代表样方 |
| 样方 Quadrat |
组成Compositions | 合计 Total |
|||||
| Ⅰ | % | Ⅱ | % | Ⅲ | % | ||
| A | 582 | 58.85 | 319 | 32.25 | 88 | 8.90 | 989 |
| B | 1337 | 87.10 | 165 | 10.75 | 33 | 2.15 | 1535 |
| C | 792 | 75.64 | 221 | 21.11 | 34 | 3.25 | 1047 |
| D | 1355 | 84.69 | 227 | 14.19 | 18 | 1.12 | 1600 |
| E | 951 | 71.50 | 353 | 26.54 | 26 | 1.95 | 1330 |
| F | 1303 | 82.47 | 262 | 16.58 | 15 | 0.95 | 1580 |
| 合计Total | 6320 | 78.21 | 1547 | 19.14 | 214 | 2.65 | 8081 |
对红背山麻杆群落6个样方的g(r) 函数分析结果表明, A样方内红背山麻杆在0—1.2 m和2.8—4.4 m尺度内表现为聚集分布, 在1.2—2.8 m和4.4—7 m尺度上表现为随机分布;B样方内背山麻杆在0—2 m尺度内表现为聚集分布, 在2—2.7 m和6.5—7 m尺度上表现为随机分布, 在2.7—6.5 m表现为分散分布;C样方内红背山麻杆在0—2.5 m尺度内表现为聚集分布, 在2.5—4.7 m尺度上表现为随机分布, 在4.7—7 m表现为分散分布;D样方内红背山麻杆在0—2.3 m尺度内表现为聚集分布, 在2.3—7 m尺度上基本表现为随机分布;E, F两个样方内红背山麻杆在0—1.4 m尺度内表现为聚集分布, 在1.4—7 m尺度上基本表现为随机分布 (图 2)。总体来看, 红背山麻杆在小尺度 (小于2 m) 上表现为聚集分布, 在大尺度 (2—7 m) 上表现为随机分布, 少数样方内出现分散分布。

|
| 图 2 红背山麻杆植株的空间点格局分析 Fig. 2 The point patter analysis of Alchornea trewioides on basis of g(r) function ―gobs为g(r) 函数值;阴影为拟合的95%的置信区间 |
由表 2可知, 3个径级红背山麻杆的空间关联性在6个样方中的表现不一致。6个样方整体来看, 径级Ⅰ和Ⅱ在0—3 m尺度下呈正相关, 在3—7 m尺度表现为不相关 (表 2);径级Ⅱ和Ⅲ在6个样方中0—1 m尺度下普遍呈正相关, 随着尺度的增加多整体表现为不相关, 少数负相关 (表 2);径级Ⅰ和Ⅲ在0—1 m尺度下呈正相关或不相关, 随着尺度的增大空间关联性表现为不相关或负相关 (表 2)。
| 样方 Quadrat |
径级 Diameter classes |
尺度Scale/m | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||
| A | Ⅰ | Ⅱ | + | + | + | + | + | + | + |
| Ⅱ | Ⅲ | + | r | - | - | r | r | r | |
| Ⅰ | Ⅲ | + | r | r | r | r | r | r | |
| B | Ⅰ | Ⅱ | + | + | + | r | - | - | - |
| Ⅱ | Ⅲ | + | + | r | r | r | r | r | |
| Ⅰ | Ⅲ | r | r | - | - | - | - | - | |
| C | Ⅰ | Ⅱ | + | + | + | r | r | r | r |
| Ⅱ | Ⅲ | + | r | r | - | - | - | r | |
| Ⅰ | Ⅲ | + | + | r | r | r | r | r | |
| D | Ⅰ | Ⅱ | + | + | + | r | r | r | r |
| Ⅱ | Ⅲ | r | - | - | r | r | r | r | |
| Ⅰ | Ⅲ | r | - | - | r | r | r | r | |
| E | Ⅰ | Ⅱ | + | r | - | - | - | - | - |
| Ⅱ | Ⅲ | + | r | r | r | r | r | r | |
| Ⅰ | Ⅲ | + | r | r | r | r | r | r | |
| F | Ⅰ | Ⅱ | + | r | - | - | r | r | r |
| Ⅱ | Ⅲ | + | + | + | r | r | r | r | |
| Ⅰ | Ⅲ | r | - | - | - | - | - | - | |
| Ⅰ代表地径≤0.5 cm;Ⅱ代表 0.5 cm<地径≤1.0 cm;Ⅲ代表地径>1.0 cm. “+”表示正相关;“-”表示负相关;“r”表示不相关 | |||||||||
本研究红背山麻杆径级Ⅰ的个体数在582—1355株/100 m2, 径级Ⅱ的个体数在165—353株/100 m2, 径级Ⅲ的个体数在15—88株/100 m2。是由于桂林喀斯特地区土层薄, 土壤瘠薄, 土壤涵养水源能力弱, 环境恶劣不利于种子的萌发生长, 从而导致红背山麻杆林下实生苗匮乏[30]。所以红背山麻杆选择克隆繁殖方式, 使其有较强的适应环境压力和抗扰动能力, 还具有很强的水平扩展能力, 通过个体之间克隆整合作用, 能有效地利用资源, 迅速开拓新生境, 扩大种群, 形成单优势种的立地[31-32], 红背山麻杆形成一棵较大径级克隆出几棵较小径级的克隆株系。故6个样方内红背山麻杆均表现出随着地径的增大植株个体逐渐减少。
在群落演替中, 种群的分布格局主要取决于种群生物学特征, 反映了种群在水平空间上的配置格局或分布情况, 它是由种群特性、种间关系和环境条件的综合影响所决定[33-34]。
本研究以克隆繁殖为主的红背山麻杆群落在较小尺度 (小于2 m) 上表现为聚集分布, 但随尺度的增大呈现为随机分布。是由于种群的空间分布格局与密度之间有着密切的关系[35], 而6个样方中红背山麻杆的密度均较大 (图 1);加之样方中没有乔木, 也少有其他灌木物种与红背山麻杆竞争, 对于红背山麻杆来说, 有充足的光照和水分条件使其能快速克隆繁殖, 且克隆繁殖母株与多个分株之间相隔较近, 整个克隆基株表现为集群分布, 故红背山麻杆在小尺度 (小于2 m) 呈集群分布。当尺度变大时, 就会出现多个红背山麻杆克隆基株, 这种一个或者少数几个克隆基株的不同分株之间的聚集就可以逐渐被忽略, 而不同基株的随机分布却更为明显。
3.2 空间关联性本研究中红背山麻杆3个径级个体之间在小尺度上均表现为正关联。在较大尺度上径级Ⅰ与径级Ⅱ也呈较明显的正关联, 但径级Ⅱ与径级Ⅲ则以相互独立的空间关系为主, 而与径级Ⅰ与径级Ⅱ及径级Ⅱ与径级Ⅲ相比, 径级Ⅰ与径级Ⅲ的空间关联性呈较强的负关联。影响空间关联性的因素有很多, 如繁殖方式、植株大小等。
刘振国等研究表明, 空间正关联或负关联主要发生在小尺度下, 空间关联性随尺度的增大而减弱, 并趋于无关联[36]。加之克隆繁殖的特殊性, 母株与分株间看作一个整体可以通过克隆整合进行资源传递[37], 也就是说整个克隆株系内无需竞争可以独占小斑块内资源。随着尺度的增大, 红背山麻杆克隆株系增多, 并有一定程度的生态位重叠, 3个径级个体之间的关联性由正关联转变为无关联或者负关联。所以本研究中红背山麻杆3个径级个体之间在小尺度上均表现为正关联。
与径级Ⅰ与径级Ⅱ及径级Ⅱ与径级Ⅲ相比, 径级Ⅰ与径级Ⅲ的空间关联性呈较强的负关联, 即随着两个径级大小差异增大, 它们的空间关联性逐渐呈无关联或负关联。这是由于植株的形体大小会影响种群空间格局[38], 高大个体对土壤水分、养分以及光照等资源的竞争会加剧, 独立性增强, 它们的正关联会著削弱, 同时伴随着聚集强度的降低[39]。据此推测, 较小植株, 对土壤水分、养分以及光照等资源的需求较少, 加之可能大部分植株属于克隆分株, 是由母株供给养分, 故总体来看径级Ⅰ与径级Ⅱ在大尺度仍表现为正关联。随着径级的增大, 植株对土壤水分、养分以及光照等资源的需求加剧, 这使得径级Ⅰ与径级Ⅲ, 以及径级Ⅱ与径级Ⅲ的空间关系逐渐呈无关联或负关联。相邻径级间的正关联较强, 而径级相差越大它们的正关联就越弱[40], 这种现象可能源于种内竞争的非对称作用, 即差异愈显著的植株, 竞争作用对弱小竞争者愈不利, 并可导致弱小竞争者的生长发育迟滞或死亡[41]。即, 红背山麻杆较大地径的母株对其他母株的克隆小分株产生较强的抑制作用。
致谢: 中国科学院广西植物研究所郭屹立和王斌老师对R软件使用的帮助。| [1] | 陈小荣, 李乐, 夏家天, 杨旭, 王伟, 丁炳扬. 百山祖亮叶水青冈种群结构和分布格局. 浙江农林大学学报, 2012, 29(5): 647–654. |
| [2] | 杨洪晓, 张金屯, 吴波, 李晓松, 张友炎. 毛乌素沙地油蒿种群点格局分析. 植物生态学报, 2006, 30(4): 563–570. |
| [3] | Ripley B D. The second-order analysis of stationary point processes. Journal of Applied Probability, 1976, 13(2): 255–266. DOI:10.1017/S0021900200094328 |
| [4] | 张金屯. 数量生态学. 北京: 科学出版社, 2004: 289–302. |
| [5] | 张金屯, 孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析. 生态学报, 2004, 24(1): 35–40. |
| [6] | 兰国玉, 雷瑞德. 植物种群空间分布格局研究方法概述. 西北林学院学报, 2003, 18(2): 17–21. |
| [7] | 董灵波, 刘兆刚, 张博, 袁野, 孙云霞. 基于Ripley L和O-ring函数的森林景观空间分布格局及其关联性. 应用生态学报, 2014, 25(12): 3429–3436. |
| [8] | 黄明钗, 史艳财, 韦霄, 吴林芳, 吴儒华, 潘子平, 曹洪麟. 珍稀濒危植物金花茶的点格局分析. 生态学杂志, 2013, 32(5): 1127–1134. |
| [9] | Perry G L W, Miller B P, Enright N J, Lamont B B. Stochastic geometry best explains spatial associations among species pairs and plant functional types in species-rich shrublands. Oikos, 2014, 123(1): 99–110. DOI:10.1111/more.2014.123.issue-1 |
| [10] | 缪宁, 刘世荣, 史作民, 薛泽冰, 喻泓, 何飞. 青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局. 生态学报, 2011, 31(1): 1–9. |
| [11] | 高福元, 赵成章, 卓马兰草. 高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性. 生态学报, 2014, 34(3): 605–612. |
| [12] | 王磊, 孙启武, 郝朝运, 田胜尼, 张姗姗, 陈一锟, 张小平. 皖南山区南方红豆杉种群不同龄级立木的点格局分析. 应用生态学报, 2010, 21(2): 272–278. |
| [13] | 马姜明, 梁士楚, 梁月明, 潘复静. 桂林岩溶石山主要灌丛类型地上生物量及分配特征. 广西师范大学学报:自然科学版, 2009, 27(4): 95–98. |
| [14] | 邓彭艳, 陈洪松, 聂云鹏, 谭巍. 桂西北喀斯特地区菜豆树和红背山麻杆旱、雨季光合特性比较. 生态学杂志, 2010, 29(8): 1498–1504. |
| [15] | Huang Y L, Chen Y Y, Yan X J, Liu J L, Yang Z M, Li D P. Chemical constituents from the leaves of Alchornea trewioides(1): Phenolic acids and related compounds. Guihaia, 2014, 34(1): 126–129. |
| [16] | Huang Y L, Liu J L, Chen Y Y, Yang Z M, Yan X J, Li D P. Chemical constituents from the leaves of Alchornea trewioides(2): Flavonoids and phenylethanoid glycosides. Guihaia, 2014, 34(2): 143–147. |
| [17] | Huang Y L, Chen Y Y, Liu J L, Yang Z M, Wang L, Li D P. Chemical constituents from the leaves of Alchornea trewioides(3): Quinic acids. Guihaia, 2015, 35(1): 105–108. |
| [18] | Huang Y L, Li D P, Yang Z M. Chemical constituents from the leaves of Alchornea trewioides(4): Polyphenols. Guihaia, 2015, 34(4): 564–568. |
| [19] | 莫源富, 奚小双, 陈忠将. 岩石-土壤-植被 (红背山麻杆、裸花紫珠) 间的元素迁移——以广西巴马岩溶石山地区为例. 中国岩溶, 2010, 29(4): 440–444. |
| [20] | Besag J. Contribution to the discussion of Dr Ripley's paper. Journal of the Royal Statistical Society Series B, 1977, 39(2): 193–195. |
| [21] | Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414–1418. DOI:10.1126/science.288.5470.1414 |
| [22] | Getis A, Franklin J. Second-order neighborhood analysis of mapped point patterns. Ecology, 1987, 68(3): 473–477. DOI:10.2307/1938452 |
| [23] | 喻泓, 杨晓晖, 慈龙骏. 地表火对红花尔基沙地樟子松种群空间分布格局的影响. 植物生态学报, 2009, 33(1): 71–80. |
| [24] | Watson D M, Roshier D A, Wiegand T. Spatial ecology of a root parasite-from pattern to process. Austral Ecology, 2007, 32(4): 359–369. DOI:10.1111/aec.2007.32.issue-4 |
| [25] | Stoyan D, Stoyan H. Fractals, Random Shapes and Point Fields: Methods of Geometrical Statistics. Chichester: Wiley, 1994. |
| [26] | 郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄俞淞, 黄甫昭, 李冬兴, 李先琨. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析. 生物多样性, 2015, 23(2): 183–191. DOI:10.17520/biods.2014126 |
| [27] | Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209–229. DOI:10.1111/oik.2004.104.issue-2 |
| [28] | Strand E K, Robinson A P, Bunting S C. Spatial patterns on the sagebrush steppe/Western juniper ecotone. Plant Ecology, 2007, 190(2): 159–173. DOI:10.1007/s11258-006-9198-0 |
| [29] | 李明辉, 何风华, 潘存德. 天山云杉天然林不同林层的空间格局和空间关联性. 生态学报, 2011, 31(3): 620–628. |
| [30] | 樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析. 生态学报, 2016, 36(2): 318–325. |
| [31] | 董鸣. 资源异质性环境中的植物克隆生长:觅食行为. 植物学报, 1996, 38(10): 828–835. |
| [32] | 蒋高明, 董鸣. 沿中国东北样带 (NECT) 分布的若干克隆植物与非克隆植物光合速率与水分利用效率的比较. 植物学报, 2000, 42(8): 855–863. |
| [33] | Muhamed H, Lingua E, Maalouf J P, Michalet R. Shrub-oak seedling spatial associations change in response to the functional composition of neighbouring shrubs in coastal dune forest communities. Annals of Forest Science, 2015, 72(2): 231–241. DOI:10.1007/s13595-014-0419-8 |
| [34] | Liu Y Y, Li F R, Jin G Z. Spatial patterns and associations of four species in an old-growth temperate forest. Journal of Plant Interactions, 2014, 9(1): 745–753. DOI:10.1080/17429145.2014.925146 |
| [35] | 宋于洋, 李园园, 张文辉. 梭梭种群不同发育阶段的空间格局与关联性分析. 生态学报, 2010, 30(16): 4317–4327. |
| [36] | 刘振国, 李镇清. 植物群落中物种小尺度空间结构研究. 植物生态学报, 2005, 29(6): 1020–1028. |
| [37] | 王长爱, 李德志, 朱志玲, 王绪平, 盛丽娟, 石强. 克隆植物生理整合作用的研究方法及其应用. 应用与环境生物学报, 2006, 12(4): 581–588. |
| [38] | 缪宁, 刘世荣, 史作民, 喻泓, 刘兴良. 川西亚高山红桦-岷江冷杉林优势种群的空间格局分析. 应用生态学报, 2009, 20(6): 1263–1270. |
| [39] | 李先琨, 向悟生, 欧祖兰, 苏宗明. 濒危植物南方红豆杉种群克隆生长空间格局与动态. 云南植物研究, 2003, 25(6): 625–632. |
| [40] | 缪宁, 史作民, 冯秋红, 刘兴良, 何飞. 川西亚高山岷江冷杉种群的空间格局分析. 林业科学, 2008, 44(12): 1–6. DOI:10.3321/j.issn:1001-7488.2008.12.001 |
| [41] | 侯继华, 黄建辉, 马克平. 东灵山辽东栎林主要树种种群11年动态变化. 植物生态学报, 2004, 28(5): 609–615. DOI:10.17521/cjpe.2004.0081 |