 2017, Vol. 37
2017, Vol. 37文章信息
- 刘莹, 李鹏, 沈冰, 冯朝红, 刘琦, 张祎
- LIU Ying, LI Peng, SHEN Bing, FENG Zhaohong, LIU Qi, ZHANG Yi.
- 采用稳定碳同位素法分析白羊草在不同干旱胁迫下的水分利用效率
- Effects of drought stress on Bothriochloa ischaemum water-use efficiency based on stable carbon isotope
- 生态学报. 2017, 37(9): 3055-3064
- Acta Ecologica Sinica. 2017, 37(9): 3055-3064
- http://dx.doi.org/10.5846/stxb201601180112
-
文章历史
- 收稿日期: 2016-01-18
- 网络出版日期: 2016-12-19





黄土丘陵半干旱区由于降雨量少且年、季度分配不均匀导致当地干旱频繁发生, 影响植物的生长[1-2]并制约当地的退耕还草工程的建设。植物水分利用效率 (WUE) 是植物生理活动过程中光合碳同化率与蒸腾速率的比值, 反映植物的耗水性和对干旱的适应性[3-4]。高水分利用效率高是植物对干旱的响应[5], 也是植物适应水分亏缺环境的一种反映表现[6]。该指标的测定主要方法是用气体交换法和稳定碳同位素方法[7]。在对青藏高原草本及灌木树种[8-10]、荒漠植物[11-12]、东北草原区植物[13]以及热带亚热带植物[14-15]的相关研究表明:植物组织稳定碳同位素分辨率Δ13C和稳定碳同位素比率δ13C与C3植物的瞬时WUE或者长期WUE有很强的相关性[16-17], 可以指示C3植物对干旱的适应情况。但是稳定碳同位素特征是否也与黄土丘陵半干旱区C4草本植物的WUE具有强相关性, 能否做为指示植物长期WUE的测定指标的研究尚未见报道, 该方法能否解释C4植物对干旱胁迫的响应机制也还未可知。
白羊草 (Bothriochloa ischaemum), C4植物, 具有再生能力强、耐踩踏、耐旱、根系发达、具有较高的生产力等特点, 是黄土丘陵半干旱区天然优质牧草[18-19], 是该地区退耕还草工程人工草地建设的主要草种, 也是黄土高原退化草地恢复和C存储的重要植物之一[20]。许多学者发现白羊草在不同土壤养分和水分[21-22]、CO2浓度[23]以及和其他植物混交播种[24]条件下, 白羊草在土壤水分含量较低时, 虽然净光合速率较小但蒸腾速率也减小, 白羊草水分利用效率不变[21]或者反而提高[25]。同时这也说明白羊草水分利用效率在指示植物干旱程度时, 也受到养分、水分、CO2浓度等环境因子的影响, 且每个因子所占的比重和敏感性也是有区别的。黄土丘陵半干旱区环境条件复杂多变, 常见研究主要探讨了单个环境因子与白羊草水分利用效率之间的关系[23-25], 但究竟哪个环境因子是主导因子还不明确。阐明在不同土壤水分条件下影响白羊草水分利用效率的主导因子, 为今后调控主导环境因子进而提高白羊草水分利用效率的研究工作提供参考依据。
本文通过盆栽实验, 研究了白羊草在不同土壤水分条件下的光合生理特性, 分析干旱胁迫下生物量积累和分配的模式, 瞬时水分利用效率与δ13C及Δ13C值之间的关系, 以及干旱胁迫下影响水分利用效率的环境因子, 以阐明白羊草对干旱胁迫的响应机制, 为黄土高原丘陵区白羊草人工草地的建设与管理提供理论支撑。
1 实验材料与方法 1.1 实验材料与设计实验于2014年6月在西安理工大学进行, 采用自制长方体有机透明玻璃容器 (尺寸19×4×27cm) 控制实验。实验用土壤采于陕北绥德王茂沟以白羊草为优势种的草地, 去除表层腐殖质和枯落物, 取0—30cm土壤均匀混合装袋, 自然风干后过2mm筛, 同时测定其容重。玻璃容器每盆装土2.5Kg容重与陕北采土样地容重相同, 为1.2g/cm3。实验用土全氮0.69g/Kg, 土壤田间持水量22%。
实验用白羊草种子与2013年10月采于陕北安塞天然草地, 采收后装在纸袋内自然状态下实验室储藏。种子于2014年5月1日播种, 每个容器撒播5穴, 出苗后每穴保留生长最旺盛的一株幼苗, 未出苗的种穴进行补植。定苗后于2014年7月1日开始控水实验处理, 设3个土壤水分梯度, 供水充分 (80%饱和田间持水量, 即80%FC, CK), 轻度胁迫 (60%饱和田间持水量, 60%FC, MS), 重度胁迫 (40%饱和田间持水量, 40%FC, SS);每个水分梯度10盆重复。
盆栽土壤含水量控制采用称重法, 每天傍晚18:00称重控制浇水, 直至2014年11月11日实验结束。每盆白羊草生物量在控水处理前计入本底值重量, 由于幼苗的初始生物量0.867g与实验结束时每盆白羊草湿生物量最大值5.812g差值仅占整体盆栽控水总重量3880g的0.1%, 且控水时间较短, 因此忽略白羊草生长重量增加对水分处理控制的影响。
1.2 实验方法与测定 1.2.1 白羊草光合特性和瞬时WUE的测定实验于2014年8月3、10、13日晴朗无云的天气条件下于9:00—11:00进行 (此时白羊草生长期为开花前期), 采用Li-6400便携式光合作用测定系统 (Li-Cor, USA) 的2×3cm透明叶室, 测定自然条件下 (自然光源, 未控制CO2浓度和湿度) 白羊草光合特性指标及环境因子参数:叶片净光合速率Pn、蒸腾速率Tr, 、气孔导度Gs、胞间CO2浓度Ci、光合有效辐射PAR、大气温度Ta、大气CO2浓度Ca、相对湿度RH、叶片大气水汽压亏缺VPD。测定选用充分展开的叶片, 每个点稳定2分钟后读数。根据上述测定参数计算叶片水分利用效率WUE=Pn/Tr。
1.2.2 白羊草生物量的测定及各器官生物量比的计算2014年11月11日, 将白羊草地上部分刈割, 分新叶、老叶、茎编号保存。盆栽中的土壤连同根系倒在塑料布中, 立即将根系从土壤中挑出, 同时区分主根 (粗根) 和枝毛根 (细根), 放入2—3℃的去离子水槽中冲洗干净, 随后根系及地上样品冲洗干净后放入105℃杀青30min, 之后在65℃下烘至恒重后用天平称重 (精确到0.001g)。
对上述各器官生物量比进行了计算:式中MNL, MOL, MS, MFR, MCR分别表示白羊草新叶、老叶、茎、细根、粗根的干物质的量 (g)。

|
白羊草各部分器官的干物质用研体研磨后过80目筛备用, 土壤样品过0.149mm筛后备用。取0.1g土壤样品或者0.005—0.006g植物样品在MultiN/C3100德国耶拿总有机碳分/总氮析仪 (Analytik Jena AG, Germany) 固体燃烧室中1050℃高温充分燃烧后生成CO2(同时记录样品TOC值g/Kg), 然后用CCIA-36d-EP二氧化碳同位素质谱仪 (Los Gatos Research, USA), 检测样品的δ13C值。植物和土壤的δ13C值的测定以PDB (Pee Dee Belemnite) 为标准, 稳定碳同位素比值据下式计算[26]:
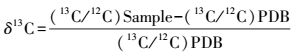
|
式中, (13C/12C) PDB为标准物质PDB的13C/12C, δ13C表示样品13C/12C与标准样品偏离的千分率;
同位素分辨率Δ13C的计算方法如下式:
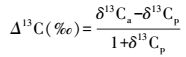
|
式中, δ13Ca表示大气CO2的碳同位素比率, 假设为-8‰[16, 27];δ13Cp表示植物或者土壤样品中CO2的碳同位素比率。
1.3 数据分析方法试验数据均采用Microsoft Excel 2013和Sigma plot10.0绘图, SPSS21.0统计分析软件处理。对瞬时WUE与δ13C和Δ13C的关系进行相关分析, 瞬时WUE与环境因子的关系进行相关分析、通径分析, 并用LSD法分析各指标的显著性, 显著性水平α=0.05。对生物量及各器官比值进行显著性检验、并用单因素方差分析 (one-way ANONA) 和Duncan法对比上述指标和比值的差异显著性水平 (α=0.05)。
2 结果与分析 2.1 干旱胁迫对白羊草光合特性及瞬时WUE的影响由表 1可知, 除胞间CO2浓度Ci外, 其余光合特性参数均随着土壤水分的减小而减小, 其中净光合速率Pn、蒸腾速率Tr、气孔导度Gs在充分供水条件下的显著高于两种水分胁迫条件。轻度干旱胁迫的水分利用效率WUE显著高于重度胁迫情况, 但与充分供水处理无显著性差异。对比CK及MS处理的净光合速率、蒸腾速率及气孔导度可以发现, 虽然MS处理的Pn和Tr均显著低于CK, 但由于其气孔导度Gs仅为0.037 mmol m-2 s-1, 说明植物此时气孔关闭, 虽然光合速率受到气孔影响而下降, 但气孔关闭也降低了蒸腾速率, 最终MS的水分利用效率显著高于CK处理。但当重度干旱胁迫时, 白羊草光合速率下降, 表明水分利用效率较低。胞间CO2浓度Ci随着干旱胁迫的加剧呈上升趋势, 但在3个水分处理之间并无显著性差异, 这与前人对大豆[28]等C3植物光合速率的研究成果相反, 但与徐伟洲对C4植物白羊草光合生理特征对干旱的响应及张文丽对C4植物玉米幼苗光合特征对干旱的响应研究结果相同[21, 29]。这可能与C3和C4植物的光合途径差异有关, 具体原因还有待后续的研究证明。
| 水分处理 Level of water stress | 充分供水 Control | 轻度胁迫 Mild stress | 重度胁迫 Serious stress |
| 净光合速率Net photosynthetic rate/(μmol m-2 s-1) | 8.756±0.582 a | 6.973±0.424 b | 5.888±0.412 b |
| 蒸腾速率Transpirations rate/(mmol m-2 s-1) | 3.907±0.216a | 2.831±0.122 b | 3.045±0.202 b |
| 气孔导度Stomatal conductance/(mmol m-2 s-1) | 0.087±0.004a | 0.073±0.003b | 0.070±0.003b |
| 胞间CO2浓度Intercellular CO2 concentration/(μmol/mol) | 202.992±4.512 a | 209.882±4.187 a | 213.686±4.704 a |
| 气孔限制值Stomatal limitation value | 0.546±0.010 a | 0.529±0.009 a | 0.517±0.011 a |
| 水分利用效率Water use efficiency/(μmol/mol) | 2.234±0.130 ab | 2.321±0.098 a | 1.992±0.095 b |
| CK:80% FC;MS:60% FC;SS:40% FC; 小写字母表示不同水分处理间差异显著 (P<0.5) | |||
由图 1可知, 白羊草总生物量随土壤水分的降低显著减少, 对于地上部分的影响显著大于对其地下部分的影响。对于地上部分, 干旱胁迫对新叶生物量无显著影响, 却显著降低白羊草老叶的生物量, 茎生物量在重度水分胁迫为0.049g, 显著低于水分充足条件0.071g。对于地下部分, 虽然根系总体生物量在3个水分处理下差异不显著, 但是进一步区分主根 (粗根) 和须根 (细根) 对土壤水分的响应, 其表现完全不同。粗根生物量在水分充足时重量为0.024g, 大于细根生物量0.014g, 随着土壤水分逐步减少, 粗根生物量减小, 细根生物量增大。重度水分胁迫时, 粗根生物量仅为0.009g, 细根生物量为0.049g, 是粗根生物量的5.444倍。

|
| 图 1 不同水分处理下白羊草各器官生物量及其比值 Fig. 1 The biomass and ratio of different parts under different water stresses |
对比图 1中SS处理的根叶比是0.386, 显著大于其他两个处理, 而SS处理的细根粗根比为6.123, 分别是MS和CK处理的5.649倍和8.056倍;但该处理的叶茎比却显著减小, 最终导致SS处理的根冠比显著大其他两个处理, 即SS处理的根冠比的显著增大受到于细根显著增加和叶片显著减少的共同作用。
2.3 干旱胁迫对白羊草各器官稳定碳同位素特征的影响表 2从干旱胁迫和白羊草各个器官两个角度阐述了稳定碳同位素比值和同位素分辨率的变化。白羊草各器官δ13C在-13.942—-11.213‰之间, 总体均值为-12.753‰。随着干旱胁迫加剧, 白羊草各器官的δ13C均呈上升趋势, 两个干旱胁迫MS和SS处理的新叶δ13C显著大于充分供水处理, SS处理茎的δ13C显著大于其他两个处理 (P<0.05)。从不同器官的δ13C值来看, 当重度干旱胁迫时各器官δ13C没有显著差异, 轻度干旱胁迫MS各器官δ13C均值表现为δ13CFR>δ13CNL>δ13COL>δ13CCR>δ13CS, 充分供水CK处理各器官δ13C均值表现为δ13CFR>δ13CCR>δ13COL>δ13CNL>δ13CS, 且δ13CFR、δ13CNL、δ13CS之间具有显著性差异。
白羊草碳同位素分辨率Δ13C的值在3.032—4.946‰之间, 不同水分处理的Δ13C的值随着干旱胁迫的增加而减小, 但只有CK处理新叶的Δ13C值差异达到显著水平, 其余处理的叶、茎、根的差异不明显。对比各处理器官的Δ13C的值发现, Δ13C的值总体呈Δ13CR<Δ13CL<Δ13CS, SS处理各器官的Δ13C的值差异不明显, CK和MS处理根系的Δ13C与叶片和茎均存在显著性差异, 叶片和茎的Δ13C之间差异不显著。
| 水分处理 Level of water stress | 充分供水 Control | 轻度胁迫 Mild stress | 重度胁迫 Serious stress | |
| 碳同位素值 | δ13CNL | -13.942±0.504 b C | -11.838±0.514 a AB | -11.922±0.776 a A |
| Carbon isotope ratio δ13C | δ13COL | -13.455±0.678 a BC | -13.331±0.269 a BC | -12.953±0.131 a A |
| δ13CS | -14.265±0.179 b C | -13.757±0.422 b C | -12.653±0.324 a A | |
| δ13CFR | -12.125±0.202 a A | -11.213±1.104 a A | -11.660±0.583 a A | |
| δ13CCR | -12.570±0.080 a AB | -13.338±0.598 a BC | -12.270±0.216 a A | |
| 碳同位素分辨率 | Δ13CNL | 4.784±0.203a A | 3.760±0.308 b AB | 3.706±0.448 b A |
| Carbon isotope discrimination Δ13C | Δ13COL | 4.540±0.294 a AB | 4.553±0.122 a A | 4.391±0.062 a A |
| Δ13CS | 4.946±0.066 a A | 4.716±0.184 a A | 4.229±0.155 a A | |
| Δ13CFR | 3.968±0.107 a C | 3.032±0.755 a B | 3.642±0.331 a A | |
| Δ13CCR | 4.208±0.040 a BC | 4.507±0.259 a A | 4.044±0.115 a A | |
| 水分利用效率Water use efficiency (WUE) | 2.234±0.130 ab | 2.321±0.098 a | 1.992±0.095 b | |
| δ13CNL、δ13COL、δ13CS、δ13CFR、δ13CCR(Δ13CNL、Δ13COL、Δ13CS、Δ13CFR、Δ13CCR) 分别表示新叶、老叶、茎、细根、粗根的碳同位素值 (分辨率).小写字母表示不同水分处理间差异显著, 大写字母表示不同器官之间差异显著 (P<0.5) | ||||
白羊草不同器官 (新叶、老叶、茎、细根、粗根) 的稳定碳同位素值、分辨率、瞬时水分利用效率WUE的相关分析结果表明 (表 3):δ13CNL、δ13COL、δ13CS、δ13CFR、δ13CCR与Δ13CNL、Δ13COL、Δ13CS、Δ13CFR、Δ13CCR分别呈极显著负相关 (相关系数均超过0.989)。δ13CNL、δ13COL、δ13CS与WUE呈显著正相关, Δ13CNL、Δ13COL、Δ13CS、Δ13CFR与WUE呈显著负相关 (P<0.05), 其中新叶的δ13CNL与WUE的相关系数最大为0.614, 其次是δ13COL为0.460, 新叶的Δ13CNL与WUE的负相关系数最大为-0.531。
| WUE | Δ13CNL | Δ13COL | Δ13CS | Δ13CFR | Δ13CCR | |
| δ13CNL | 0.614** | -0.989** | -0.581** | -0.353 | 0.307 | -0.081 |
| δ13COL | 0.460* | -0.554** | -0.996** | -0.150 | 0.250 | -0.301 |
| δ13CS | 0.406* | -0.321 | -0.110 | -0.998** | -0.384 | 0.279 |
| δ13CFR | 0.373 | 0.475** | 0.265 | -0.334 | -0.988** | 0.644** |
| δ13CCR | -0.309 | -0.129 | -0.309 | 0.279 | 0.737** | -0.996** |
| Δ13CNL | -0.531** | 0.526** | 0.358 | -0.387 | 0.130 | |
| Δ13COL | -0.402* | 0.106 | -0.283 | 0.309 | ||
| Δ13CS | -0.398* | 0.345 | -0.264 | |||
| Δ13CFR | -0.446* | -0.707** | ||||
| Δ13CCR | 0.268 |
植物叶片水分利用效率WUE主要表征植物生存生长过程对环境的适应的重要因素, 是光合碳同化率 (净光合速率Pn) 与蒸腾速率 (Tr) 的比值, 而这2个因子又受到其他环境因子 (光合有效辐射PAR、大气CO2浓度Ca, 叶片温度Tl, 空气湿度RH等, 空气水分浓度Ha) 的影响。在干旱胁迫环境下, 植物可通过以下两种机制提高水分利用效率:1) 减低气孔导度, 使蒸腾速率减低高于光合速率的降低;2) 提高光合速率, 使光合速率的增加高于水分消耗的增加。本研究发现3种水分处理各个因子对WUE影响的通径分析 (表 4) 表明, 各因子对WUE的影响效应均不相同, CK处理影响效应为Tl>Ta>Ca>Ha>RH>PAR>D, MS为D>RH>PAR>Ha>Ca>Ta>Tl, SS为Ta>D>Ca>PAR>Ha>RH>Tl。CK处理的水分利用效率受到Tl的影响均最大, 为5.064, 主要来自于Tl的直接影响和D对其间接影响;MS受到D影响最大 (8.771), 分别来自自身直接影响和Tl间接影响;而SS处理主要来源Ta的直接影响。对比各个环境因子对WUE的通径分析发现CK和MS处理下, Gs、Tr与WUE极显著正相关;但在SS处理下却完全相反, 同时净光合速率也显著降低。这说明当水分胁迫时, 白羊草自身的水分已经不能维持其自身生长, 使气孔关闭, 光合速率下降, 导致水分利用效率降低。当水分能满足植物自身生长需要时, 气孔打开, 净光合速率增大, 虽然此时蒸腾速率也增大, 但显然净光合速率增大的幅度显著大于蒸腾速率, 水分利用效率提高。Lee K H等[30]认为干旱胁迫时植物通过改变合成ABA的关键酶, 9-顺式-环氧类胡萝卜双氧合酶 (9-cis-epoxycarotenoid dioxygenase, NCED) 调控干旱胁迫[31]。田伟丽等[32]对马铃薯的水分利用效率的研究表明, 在严重水分胁迫下 (土壤相对含水量25%), 植物激素脱落酸 (ABA) 含量增加33%。这可能也是导致白羊草表现出CK和MS条件下, 水分利用效率高, SS水分利用效率低的主要原因。
| 影响因子 Factors | 总效应 Total effect | 直接通径系数 Direct effect | 间接通径系数Indirect effect | ||||||||
| WUE | D | Ta | Tl | Ca | Ha | RH | PAR | ||||
| CK | D | -4.561 | -2.406** | 1.241 | - | -1.079 | -2.305 | 1.124 | -1.024 | 0.891 | 0.238 |
| Ta | 0.010 | -1.736 | 1.343 | -0.779 | - | -0.808 | 1.560 | -1.501 | 1.673 | 1.602 | |
| Tl | 5.065 | 2.525** | -0.910 | 2.419 | 1.175 | - | -1.259 | 1.321 | -0.883 | -0.234 | |
| Ca | -0.013 | -0.128 | -0.078 | 0.060 | 0.115 | 0.064 | - | 0.091 | -0.115 | -0.099 | |
| Ha | -0.084 | -0.128 | 0.075 | -0.054 | -0.111 | -0.067 | 0.091 | - | 0.090 | 0.096 | |
| RH | -0.253 | -0.568 | -0.437 | 0.210 | 0.547 | 0.199 | -0.511 | 0.398 | - | -0.528 | |
| PAR | -0.289 | -0.342 | -0.251 | 0.034 | 0.316 | 0.032 | -0.266 | 0.256 | -0.318 | - | |
| MS | D | 8.771 | 3.493 | 1.273 | - | 1.748 | 3.453 | 0.510 | -0.724 | -2.899 | 3.190 |
| Ta | -4.220 | -2.221 | 1.012 | -1.112 | - | -1.368 | 1.386 | -1.429 | 0.780 | -0.257 | |
| Tl | -14.915 | -5.670 | -1.395 | -5.605 | -3.493 | - | -0.075 | 0.337 | 4.427 | -4.836 | |
| Ca | -0.136 | 0.730** | 0.671 | 0.107 | -0.456 | 0.010 | - | -0.602 | -0.228 | 0.303 | |
| Ha | 2.273 | 4.126* | -3.103 | -0.855 | 2.654 | -0.246 | -3.404 | - | 2.017 | -2.020 | |
| RH | 4.728 | -3.099* | 1.340 | 2.572 | 1.088 | 2.419 | 0.968 | -1.515 | - | 2.294 | |
| PAR | 3.996 | 1.933 | 1.175 | 1.765 | 0.223 | 1.648 | 0.803 | -0.946 | -1.430 | - | |
| SS | D | 0.860 | 0.795 | 0.270 | - | 0.240 | 0.789 | -0.694 | -0.279 | -0.656 | 0.665 |
| Ta | 0.977 | 0.587 | -0.396 | 0.177 | - | 0.242 | -0.044 | 0.430 | -0.263 | -0.153 | |
| Tl | -3.997 | -3.181 | -0.766 | -3.155 | -1.313 | - | 2.706 | 0.753 | 2.636 | -2.442 | |
| Ca | 0.494 | -0.732 | 0.392 | 0.639 | 0.055 | 0.623 | - | -0.292 | -0.400 | 0.601 | |
| Ha | 0.288 | 0.278 | -0.232 | -0.098 | 0.204 | -0.066 | 0.111 | - | 0.075 | -0.217 | |
| RH | 0.210 | -0.239 | 0.025 | 0.198 | 0.107 | 0.198 | -0.131 | -0.064 | - | 0.141 | |
| PAR | 0.329 | 2.132 | 1.553 | 1.784 | -0.555 | 1.636 | -1.751 | -1.660 | -1.257 | - | |
| 气孔导度Gs, 胞间CO2浓度Ci, 蒸腾速率Tr, 大气水汽压亏缺D, 叶片温度Tl, 空气CO2浓度Ca, 空气水分含量Ha, 空气相对湿度RH, 光合有效辐射PAR;*表示叶片净光合速率与对应影响因子相关性显著, **表示叶片净光合速率与对应影响因子相关性极显著. Gs: stomatal conductance, Ci: intercellular CO2 concentration, Tr: transpiration rate, D: Atmospheric water vapor pressure deficit, Tl: leaf temperature, Ca: atmospheric carbon dioxide concentration, Ha: atmospheric humidity, RH:relative humidity, PAR—photosynthetic active radiation; CK: Control, MS: mide stress, SS: serious stress | |||||||||||
白羊草的WUE表示为叶片进行光合作用过程中, 光合速率与蒸腾速率的比值。随着干旱胁迫加剧, 为了充分利用水分, 植物常常关闭气孔, 减小气孔导度, 降低蒸腾速率, 导致胞间CO2浓度降低, 此时植物能选择吸收的CO2减少, 最终导致δ13C值增大, Δ13C值减小[33-34]。腰政懋等[35]研究发现, 辽东栎碳同位素值δ13C随干旱胁迫的增加而增加;张辉[36]等对甘薯的研究得到其碳同位素分辨率Δ13C值随土壤含水量的降低;陈平等[37]研究表明, 干旱条件下随着水分亏缺的加剧, 菘蓝和决明子的δ13C显著增大、两种植物的Δ13C显著减小。白羊草是C4植物, 光合作用工程中的Δ13C和WUE的关系为:

|
式中, Δ13C为白羊草各个器官的碳同位素分辨率, a是CO2扩散过程中引起的碳同位素判别值, b1是气态CO2通过PEP羧化酶固定产生的判别值, b2为RuBP羧化酶对13C的判别值, ψ表示从维管束鞘中泄露到叶肉细胞中的CO2比例, Ca为大气中的CO2浓度, Ci为白羊草叶片的胞间CO2浓度, ν是蒸汽压亏缺, ν=μ(1-RH), μ为大气饱和蒸汽压, RH空气相对湿度, 1.6是气孔对大气和CO2传导率的转换系数。理论上WUE与Δ13C呈负相关, 本文研究发现白羊草WUE与叶片Δ13C的相关系数为-0.531, 呈现极显著负相关 (P<0.01), 同时WUE与叶片的δ13C也呈极显著正相关, 这与其他研究说明采用叶片Δ13C研究它与WUE关系更为适宜[38]的结果一致, 并且在其他C4植物的研究上也得到了证实[33]。因此可以通过测定C4植物叶片的δ13C或者Δ13C值来判断干旱条件下白羊草水分利用情况同样可行。此方法在黄土高原有限的水分条件下, 对选育该区高水分利用效率的植物提供快速简便的方法。
3.2 白羊草生物量分配对干旱胁迫的响应植物的叶片和根是植株的“源”器官和“库”器官, 源器官通过光合作用获得光合同化物 (碳水化合物) 通过茎向库器官运输, 使得植株整体协调生长。这种“源库”关系由于物种、CO2浓度、干旱胁迫强度、干旱胁迫时间等因素的变化而变化[39-40]。一些学者认为[25, 41]:干旱胁迫使植物根系生长追逐水源, 根冠竞争碳水化合物, 同化产物多分配向根系, 促进根冠比增大。本文结果证实了这一论断, 重度水分胁迫下的根冠比显著大于其他水分处理, 且根叶比是其他处理2—2.2倍, 与Fernández RJ[39]对C4草本植物和韦莉莉[42]对杉木的研究结果具有相同趋势, 这说明干旱胁迫加强光合产物从源器官向库器官的转运, 增加库器官的光合产物比例。本文进一步分析细根和粗根比, 发现重度干旱胁迫时, 从叶片“源”流入根系“汇”的碳并未在根系中均匀分配, 而是主要用于细根的生长, 这主要是因为细根对养分和水分的有效性的敏感程度大于粗根, 细根的主要生理功能是吸收养分和水分, 粗根主要起运输和支持作用[43], 这种“汇”碳分配的不均性, 遵循植物个体最优化分配原则[44], 最终导致细根生物量增加而粗根生物量减少的结果。
3.3 白羊草各器官碳同位素组成的差异许多研究学者通过对C4植物玉米[38]、C3植物甘薯[36]、旱稻[45]等不同器官的碳同位素分馏分析发现, C3植物各器官的δ13C值均存在显著性差异, C4植物叶片和茎的δ13C差异不显著, C3和C4植物的籽粒和根系均比叶片和茎更易富集13C。本研究方差分析结果显示, CK和MS不同器官的δ13C和Δ13C值差异达显著水平, 而SS处理各器官的差异不显著, 其原因可能是不同水分处理对白羊草13C分馏产生影响。总结各器官的差异表现为Δ13C值茎、新叶、老叶之间差异不显著, 结合图 2可以发现, 说明白羊草叶片和茎之间未发生13C分馏, 但茎和叶的Δ13C值均显著大于根, 说明根与茎叶之间已经产生了13C分馏。Brugnoli和Farquhar[46]以及Leavitt和Long[47]提出植物13C分馏有3个原因造成:1) 各个器官的化学组成成分不同导致13C分馏, 各种化学成分具有各自稳定的δ13C值, 器官的纤维素含量高则较容易13C富集, 木质素和油脂含量高则13C含量低;2) 植物器官生理生态特征差距大, 在合成碳水化合物过程及其通过韧皮部的输出、装载、下载的过程中以及光合同化物从“源”运输到“库”过程中都可能发生13C的二次分馏;3) 不同器官的呼吸特征具有很大差异, 植物器官在呼吸时优先利用12C, 从而使13C在组织中富集。根系是植物的“库”器官, 根系的碳在积累的过程中受到环境和代谢因素的综合作用, 从“源”到“库”的转换过程中都可能出现13C的二次分馏, 且根系和叶片虽然都作为呼吸器官, 但其呼吸速率显著不同, 这可能也是造成的其13C分馏的主要原因。因此, 在对不同植物的δ13C和Δ13C值进行比较时, 应根据具体植物的不同器官进行具体分析。
4 结论(1) 干旱胁迫显著降低白羊草的净光合速率、蒸腾速率、气孔导度, 但水分利用效率在适度干旱时最大, 重度干旱胁迫时最小。
(2) 重度干旱胁迫显著降低植物整体生物量, 但增加白羊草根冠比、根叶比和细根粗根比, 说明植物适应干旱环境时, 对地下的生物量分配增加, 且主要增加了细根生物量, 降低粗根生物量, 促进植物细根生长;
(3) 随着干旱胁迫加剧, 白羊草各器官的δ13C均呈上升趋势, MS和SS处理的新叶δ13C显著大于CK处理, Δ13C呈减小趋势, 且仅有CK处理新叶的Δ13C值差异达到显著水平;白羊草SS处理不同器官δ13C和Δ13C没有显著差异, CK和MS处理的各器官δ13C均值表现分别为细根>粗根>老叶>新叶>茎、细根>新叶>老叶>粗根>茎, CK和MS处理Δ13C的值总体呈现根<叶<茎。
(4) 3种水分处理下, 新叶的δ13CNL与WUE的相关系数最大为0.614, 其次是δ13COL为0.460, 新叶的Δ13CNL与WUE的负相关系数最大为-0.531, 说明利用稳定碳同位素方法测定白羊草水分利用效率具有可行性。
(5) CK水分处理的水分利用效率受到叶面温度的影响最大, 主要来源叶面温度的直接影响和大气水汽压亏缺对其的间接效应;MS处理受大气水汽压亏缺影响最大, 主要来源其直接影响和叶面温度的间接效应;SS处理受空气温度影响最大, 主要来源空气温度直接影响和空气水分含量对其的间接效应。
| [1] | 徐朗然, 张继敏, 丁士友. 黄土高原白羊草草原的基本特征及其地理学意义. 西北植物学报, 1997, 17(1): 88–93. |
| [2] | 魏胜林, 刘竞男, 王陶, 张婉, 余增亮. N+注入对甘草种子萌发和根发育效应及作用机制. 草业学报, 2004, 13(5): 112–115. |
| [3] | Martin B, Tauer C G, Lin R K. Carbon isotope discrimination as a tool to improve water-use efficiency in tomato. Crop Science, 1999, 39(6): 1775–1783. DOI:10.2135/cropsci1999.3961775x |
| [4] | Ray I M, Townsend M S, Muncy C M. Heritabilities and interrelationships of water-use efficiency and agronomic traits in irrigated alfalfa. Crop Science, 1999, 39(4): 1088–1092. DOI:10.2135/cropsci1999.0011183X003900040022x |
| [5] | 孙学凯, 范志平, 王红, 白洁, 张营, 邓东周. 科尔沁沙地复叶槭等3个阔叶树种的光合特性及其水分利用效率. 干旱区资源与环境, 2008, 22(10): 188–194. |
| [6] | Jaleel C A, Gopi R, Sankar B, Gomathinayagam M, Panneerselvam R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. Comptes Rendus Biologies, 2008, 331(1): 42–47. DOI:10.1016/j.crvi.2007.11.003 |
| [7] | 王云霓, 熊伟, 王彦辉, 于澎涛, 徐丽宏, 左海军, 曹恭祥, 孙浩. 干旱半干旱地区主要树种叶片水分利用效率研究综述. 世界林业研究, 2012, 25(2): 17–23. |
| [8] | 刘光琇, 陈拓, 安黎哲, 王勋陵, 冯虎元. 青藏高原北部植物叶片碳同位素组成特征的环境意义. 地球科学进展, 2004, 19(5): 749–753. |
| [9] | 周咏春, 樊江文, 钟华平, 张文彦. 青藏高原草地群落植物碳同位素组成与海拔梯度的关系. 中国科学:地球科学, 2013, 43(1): 120–130. |
| [10] | 邱权, 潘昕, 李吉跃, 王军辉, 董蕾, 马建伟, 杜坤. 青藏高原20种灌木幼苗生物量分配、水分利用效率及叶片δ13C比较. 西北林学院学报, 2014, 29(4): 8–14. |
| [11] | 陈拓, 冯虎元, 徐世建, 强维亚, 安黎哲. 荒漠植物叶片碳同位素组成及其水分利用效率. 中国沙漠, 2002, 22(3): 288–291. |
| [12] | 苏培玺, 陈怀顺, 李启森. 河西走廊中部沙漠植物δ13C值的特点及其对水分利用效率的指示. 冰川冻土, 2003, 25(5): 597–602. |
| [13] | 苏波, 韩兴国, 李凌浩, 黄建辉, 白永飞, 渠春梅. 中国东北样带草原区植物δ13C值及水分利用效率对环境梯度的响应. 植物生态学报, 2000, 24(6): 648–655. |
| [14] | 林植芳, 林桂珠, 孔国辉, 张鸿彬. 生长光强对亚热带自然林两种木本植物稳定碳同位素比、细胞间CO2浓度和水分利用效率的影响. 热带亚热带植物学报, 1995, 3(2): 77–82. |
| [15] | 渠春梅, 韩兴国, 苏波, 黄建辉, 蒋高明. 云南西双版纳片断化热带雨林植物叶片δ13C值的特点及其对水分利用效率的指示. 植物学报, 2001, 43(2): 186–192. |
| [16] | Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 1989, 40(1): 503–537. DOI:10.1146/annurev.pp.40.060189.002443 |
| [17] | Farquhar G D, Richards R A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Australian Journal of Plant Physiology, 1984, 11(6): 539–552. DOI:10.1071/PP9840539 |
| [18] | 徐炳成, 山仑, 李凤民. 半干旱黄土丘陵区五种植物的生理生态特征比较. 应用生态学报, 2007, 18(5): 990–996. |
| [19] | 程杰, 呼天明, 程积民. 黄土高原白羊草种群分布格局对水热梯度的响应. 草地学报, 2010, 18(2): 167–171. DOI:10.11733/j.issn.1007-0435.2010.02.005 |
| [20] | 山仑, 陈国良. 黄土高原旱地农业的理论与实践. 北京: 科学出版社, 1993: 215–246. |
| [21] | 徐伟洲, 徐炳成, 段东平, 牛富荣. 不同水肥条件下白羊草光合生理生态特征研究Ⅰ.光合生理日变化. 草地学报, 2010, 18(5): 629–635. DOI:10.11733/j.issn.1007-0435.2010.05.004 |
| [22] | 徐伟洲, 徐炳成, 段东平, 牛富荣. 不同水肥条件下白羊草光合生理生态特征研究Ⅱ.光响应曲线. 草地学报, 2010, 18(6): 773–779. DOI:10.11733/j.issn.1007-0435.2010.06.005 |
| [23] | 张昌胜, 刘国彬, 薛萐, 冀智清, 张超. 干旱胁迫和CO2浓度升高条件下白羊草的光合特征. 应用生态学报, 2012, 23(11): 3009–3015. |
| [24] | 丁文利, 舒佳礼, 徐伟洲, 徐炳成. 水分胁迫和组合比例对白羊草与达乌里胡枝子叶绿素荧光参数的影响. 草地学报, 2014, 22(1): 94–100. DOI:10.11733/j.issn.1007-0435.2014.01.015 |
| [25] | 徐炳成, 山仑, 黄瑾, 黄占斌. 柳枝稷和白羊草苗期水分利用与根冠比的比较. 草业学报, 2003, 12(4): 73–77. |
| [26] | Farquhar G D, O'Leary M H, Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, 1982, 9(2): 121–137. DOI:10.1071/PP9820121 |
| [27] | Voltas J, Serrano L, Hernández M, Pemán J. Carbon isotope discrimination, gas exchange and stem growth of four Euramerican hybrid poplars under different watering regimes. New Forests, 2006, 31(3): 435–451. DOI:10.1007/s11056-005-0879-7 |
| [28] | 徐晨, 张鹏, 徐克章, 李大勇, 季平, 冯引娣, 赵兰坡. 干旱胁迫对不同大豆品种叶片光合及生理特性的影响. 中国油料作物学报, 2013, 35(6): 674–679. DOI:10.7505/j.issn.1007-9084.2013.06.009 |
| [29] | 张文丽, 张彤, 吴冬秀, 张岁岐, 山仑. 土壤逐渐干旱下玉米幼苗光合速率与蒸腾速率变化的研究. 中国生态农业学报, 2006, 14(2): 72–75. |
| [30] | Lee K H, Piao H L, Kim H Y, Choi S M, Jiang F, Hartung W, Hwang I, Kwak J M, Lee I J, Hwang I. Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell, 2006, 126(6): 1109–1120. DOI:10.1016/j.cell.2006.07.034 |
| [31] | Qin X Q, Zeevaart J A D. The 9-cis-epoxycarotenoid cleavage reaction is the key regulatory step of abscisic acid biosynthesis in water-stressed bean. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(26): 15354–15361. DOI:10.1073/pnas.96.26.15354 |
| [32] | 田伟丽, 王亚路, 梅旭荣, 李玉中, 郭家选. 水分胁迫对设施马铃薯叶片脱落酸和水分利用效率的影响研究. 作物杂志, 2015(1): 103–108. |
| [33] | Wingate L, Ogée J, Burlett R, Bosc A, Devaux M, Grace J, Loustau D, Gessler A. Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration. New Phytologist, 2010, 188(2): 576–589. DOI:10.1111/j.1469-8137.2010.03384.x |
| [34] | Chaves M M, Pereira J S, Maroco J, Rodrigues M L, Ricardo C P P, Osório M L, Carvalho I, Faria T, Pinheiro C. How plants cope with water stress in the field? Photosynthesis and growth. Annals of Botany, 2002, 89(7): 907–916. DOI:10.1093/aob/mcf105 |
| [35] | 腰政懋, 柴源, 冯博, 徐程扬. 水分胁迫下7个种源辽东冷杉幼苗水分利用效率差异. 北京林业大学学报, 2015, 37(6): 27–34. |
| [36] | 张辉, 朱绿丹, 宁运旺, 张丛志, 张永春. 土壤水分条件对甘薯水分利用效率和稳定性碳同位素影响. 土壤, 2014, 46(5): 806–813. |
| [37] | 陈平, 孟平, 张劲松, 何春霞, 贾长荣, 李建中. 两种药用植物生长和水分利用效率对干旱胁迫的响应. 应用生态学报, 2014, 25(5): 1300–1306. |
| [38] | 张丛志, 张佳宝, 赵炳梓, 张辉, 黄平, 李晓鹏, 朱强根. 玉米水分利用效率、碳稳定同位素判别值和比叶面积之间的关系. 作物学报, 2009, 35(6): 1115–1121. |
| [39] | Fernández R J, Wang M B, Reynolds J F. Do morphological changes mediate plant responses to water stress? A steady-state experiment with two C4 grasses. New Phytologist, 2002, 155(1): 79–88. DOI:10.1046/j.1469-8137.2002.00438.x |
| [40] | 许振柱, 周广胜, 肖春旺, 王玉辉. CO2浓度倍增和土壤干旱对两种幼龄沙生灌木碳分配的影响. 植物生态学报, 2005, 29(2): 281–288. DOI:10.17521/cjpe.2005.0036 |
| [41] | 冯广龙, 刘昌明, 王立. 土壤水分对作物根系生长及分布的调控作用. 生态农业研究, 1996, 4(3): 5–9. |
| [42] | 韦莉莉, 张小全, 侯振宏, 徐德应, 余雪标. 杉木苗木光合作用及其产物分配对水分胁迫的响应. 植物生态学报, 2005, 29(3): 394–402. DOI:10.17521/cjpe.2005.0052 |
| [43] | 梅莉, 王政权, 韩有志, 谷加存, 王向荣, 程云环, 张秀娟. 水曲柳根系生物量、比根长和根长密度的分布格局. 应用生态学报, 2006, 17(1): 1–4. |
| [44] | Niklas K J. A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categories. New Phytologist, 2006, 171(1): 27–40. DOI:10.1111/nph.2006.171.issue-1 |
| [45] | Zhao B Z, Kondo M, Maeda M, Ozaki Y, Zhang J B. Water-use efficiency and carbon isotope discrimination in two cultivars of upland rice during different developmental stages under three water regimes. Plant and Soil, 2004, 261(1/2): 61–75. DOI:10.1023/B:PLSO.0000035562.79099.55 |
| [46] | Brugnoli E, Farquhar G D. Photosynthetic fractionation of carbon isotopes//Leegood R C, Sharkey T D, von Caemmerer S, eds. Photosynthesis: Physiology and Metabolism. Netherlands: Springer, 2000: 352-434. |
| [47] | Leavitt S W, Long A. Stable-carbon isotope variability in tree foliage and wood. Ecology, 1986, 67(4): 1002–1010. DOI:10.2307/1939823 |