 2017, Vol. 37
2017, Vol. 37文章信息
- 石耀辉, 周广胜, 蒋延玲, 王慧, 许振柱, 麻雪艳
- SHI Yaohui, ZHOU Guangsheng, JIANG Yanling, WANG Hui, XU Zhenzhu, MA Xueyan.
- 贝加尔针茅响应降水变化敏感指标及关键阈值
- Thresholds of Stipa baicalensis sensitive indicators response to precipitation change
- 生态学报. 2017, 37(8): 2620-2630
- Acta Ecologica Sinica. 2017, 37(8): 2620-2630
- http://dx.doi.org/10.5846/stxb201601270198
-
文章历史
- 收稿日期: 2016-01-27
- 网络出版日期: 2016-10-29





2. 中国科学院植物研究所植被与环境变化国家重点实验室, 北京 100093
2. State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
北方温带草原是我国草地的主体, 具有重要的生态和生产功能, 主要分布于干旱-半干旱气候区, 这一区域是受降水影响的敏感和脆弱地带[1-5]。降水变化会对牧草的产量和质量产生直接影响, 是影响草原群落结构和生产力的关键因子, 草原干旱是我国北方牧区最为严重的气象灾害之一, 发生次数多、分布范围广, 持续时间长, 影响程度尤为强烈[6-8]。历史记录表明, 自20世纪70年代以来, 中国北方干旱事件发生增加, 极端干旱频率升高, 半干旱边界已经向东南移动[9-11]。未来气候变化情景下, 干旱、强降水等极端天气气候事件频发, 且来势早、强度大, 并有加剧的趋势[12-13], 使得降水变化对草原的影响与应对越来越受到关注。
干旱指标是确定干旱是否发生及其严重程度的标准, 是干旱研究分析的基础。国内外已就干旱指标开展了大量研究, 不同行业从各自的角度出发, 得出一系列干旱指标。有代表性的气象指标、土壤墒情指标和植物生理生态指标分别从大气干燥程度、土壤供水能力和植物耐旱能力方面描述了植物的缺水程度。各类干旱指标都具有一定的时间与空间尺度范围, 有其不同的优缺点和适用性[7, 10, 14-17]。气象指标可以大致反映干旱发生趋势但很难直接表示植物遭受干旱的影响程度和机理;土壤墒情指标, 能较好指示植物对水分胁迫响应的状况但有其不确定性, 不能很好评估大尺度干旱问题[7, 15, 17];植物生理生态指标是植物耐旱能力的具体体现, 直接反映植物受旱程度, 主要包括植物生理指标和植物形态指标两类[17-19]。研究发现, 植株高度与水分呈正相关[20-21], 干旱胁迫下单叶面积减小, 叶数降低, 植株总叶面积减小[4]。随着水分减少多数植物比叶面积减小, 叶片增厚, 形态生长受到抑制[22-24]。在受到干旱胁迫时叶片含水量显著下降[25]。Siddique等[26]研究发现叶水势是反映植物对水分胁迫响应较好的指标。叶片叶绿素含量随水分变化的增减物种间差异较大[27-29]。以往研究为认识植物对干旱的响应提供了支持, 但植物响应干旱的生理生态指标多, 变异大, 且常为定性指标, 主观性强, 缺乏定量分析, 给应用带来不便[15, 17, 30-32]。
本研究针对目前草原植物干旱指标研究中存在的上述问题, 以贝加尔针茅 (Stipa baicalensis) 为研究材料, 基于不同降水变化模拟实验, 尝试从植物个体水平出发量化形态、生理和生物量特征对降水变化的敏感性, 筛选敏感指标, 确定敏感指标发生显著变化的关键阈值, 为草原植物干旱的监测、预测和干旱的解除提供参考。
1 研究方法 1.1 实验设计实验于2011年4月至9月在中国科学院植物研究所进行。贝加尔针茅种子于实验前1年采集于中国科学院植物研究所乌兰察布草地生态研究站 (41°43′N, 111°52′E, 海拔1420—1500 m), 经0.5%高锰酸钾溶液消毒8min, 用水冲洗后备播。土壤为内蒙古草原典型栗钙土, 土壤有机碳含量、总氮含量和有效氮含量分别为 (12.31±0.19) g/kg、(1.45±0.02) g/kg和 (81.61±0.71) mg/kg, 将土壤过筛去除其中的残根、石块等分装入聚乙烯塑料盆中。以贝加尔针茅种源地内蒙古乌兰察布盟四子王旗6、7和8月的30年 (1978—2007) 月均降水量为对照设置5个降水量梯度:-30%、-15%、对照、+15%和+30%, 每个梯度6个重复。根据塑料盆尺寸 (上口10.9 cm, 下底8.5 cm, 高9.5 cm) 将月均降水量换算为各处理的总浇水量, 每3天浇灌1次。具体降水处理梯度数据见表 1。
| 月份Month | 降水量Precipitation/mm | ||||
| -30% | -15% | 对照Control | +15% | +30% | |
| 6 | 36 | 44 | 51 | 59 | 67 |
| 7 | 62 | 75 | 88 | 101 | 114 |
| 8 | 70 | 85 | 100 | 115 | 130 |
| 总量Total | 168 | 204 | 240 | 275 | 311 |
水分控制前先进行育苗、间苗及定苗。(1) 育苗:4月18日播种, 播种后放入温室育苗 (最大光强约1000 μmol m-2 s-1, 白天和夜间温度分别为26—28℃和18—20℃), 定期浇水。(2) 间苗及定苗:在出苗至第1片叶完全展开后进行间苗, 直到幼苗第3片叶完全展开时进行定苗, 每盆选取长势较好且一致的幼苗保留4株。2011年5月31日开始控制水分。控水开始前对每盆样品进行称重、补水以保证初始水分条件一致。控制实验开始后每3天在16:00后浇水1次。
1.2 指标测定(1) 形态特征
在9月2日对植株进行收获, 统计各处理下每盆植株叶片数, 测定株高 (cm), 利用WinFOLIA (Régent Instruments Inc., Quebec, Canada) 植物叶片分析系统测定植株总叶面积 (cm2)。单叶面积 (cm2)=总叶面积/叶数。比叶重为单位叶面积重量, 比叶重=叶干重/叶面积, 为比叶面积倒数 (mg/cm2)。
(2) 生理特征
叶含水量 (%) 叶片含水量=(鲜重-干重)/鲜重×100%。
叶C、N含量 (%) 为单位重量含量。生物量测定后的烘干叶片样品采用MM400混合研磨仪 (Retsch GmbH, Haan, Germany) 研磨后, 称取8 mg样品, 用锡箔纸包好放入Vario EL Ⅲ元素分析仪 (Elementar Analysensysteme GmbH, Hanau, Germany) 孔槽内, 在950℃加热条件下燃烧, 自动测定样品C和N, 每处理3个重复。
叶丙二醛 (MDA) 含量 (nmol/g):参考Hernández and Almansa[33]方法, 收获时采用双组分分光光度法测定, 每处理3个重复。
叶绿素含量 (mg/g) 测定采用乙醇浸提、分光光度法。经10mL 95%的酒精抽提、采用紫外分光光度计测定663 nm和645 nm处的光密度, 进而计算叶绿素含量, 每处理3个重复。
叶水势 (MPa) 植物收获时, 采用WP4-T露点水势仪 (Decagon Devices, Inc., Pullman, WA, USA) 进行叶片水势测定, 每处理3个重复。
(3) 生物量采用收获法进行测定, 测定每盆植株的地上和地下 (根) 的生物量 (65℃下烘干至恒重后采用感量0.01 g电子天平称量)。
1.3 数据分析采用统计分析软件SPSS 16.0的“单因素方差分析”分析降水变化对观测指标影响的显著性, 若影响显著, 则采用Duncan多重比较在0.05水平上分析降水处理间差异, 从而确定各指标对降水变化的敏感性。敏感指标对降水变化的阈值是指标受到降水变化影响而发生显著性变化的点。本研究引入正态总体统计容忍区间及容忍限的估算方法, 根据有限的观测样本推算其所在总体的上限 (μU) 和下限 (μD) 范围, 从而确定干旱敏感指标的受旱临界点, 统计容忍区间 (statistical tolerance interval) 是由随机样本确定的、以规定的概率至少包含抽样总体规定比例的区间, 实质是利用样本数据估算指定比例的总体的上下限[34]。《中华人民共和国国家标准 (GBT 3359—2009) 数据的统计处理和解释-统计容忍区间的确定》中提供了不同类型连续分布总体的容忍区间和容忍限的计算方法。在此, 采用总体均值和方差未知时正态总体容忍限的计算方法:
容忍上限计算式
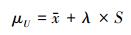
|
(1) |
容忍下限计算式
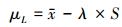
|
(2) |
式中, x为样本均值, S为样本方差, 利用提供的附表D4查找相应的λ值。
为便于指标间比较, 阈值计算时采用“极差标准化法”将实测值 (xi) 标准化为xi′, 消除量纲:
当指标与降水呈正相关时采用式
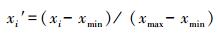
|
(3) |
当指标与降水呈负相关时采用式
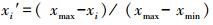
|
(4) |
降水量变化对株高、分蘖数、叶数、总叶面积和单叶面积都产生了显著影响, 但对比叶重的影响未达显著水平 (图 1, 表 2)。随着降水量的由多到少, 株高、分蘖数、叶数、总叶面积和单叶面积明显降低, 但不同的形态指标对降水量变化的敏感性差别明显。株高和总叶面积变化在处理组间变化最显著, 在降水从311 mm降到167 mm之间, 有3次显著变化, 株高分别出现在311—239、275—203 mm和239—167 mm之间, 总叶面积分别出现在311—275、275—239 mm和239—203 mm之间。分蘖数和叶数次之, 有两次显著变化, 分蘖数分别出现在311—239 mm和275—203 mm之间, 叶数分别出现在275—239 mm和239—203 mm之间。单叶面积仅在311—275 mm之间有显著变化, 其他降水水平之间差异不显著。

|
| 图 1 降水量变化对贝加尔针茅形态特征的影响 Fig. 1 Effects of precipitation change on morphological characters of S. baicalensis |
| 形态特征Morphological characteristics | df | F | P |
| 株高Plant heights | 4 | 18.522 | 0.000 |
| 分蘖数Tiller number | 4 | 5.797 | 0.011 |
| 叶数Leaf number | 4 | 34.677 | 0.000 |
| 总叶面积Total leaf area | 4 | 40.896 | 0.000 |
| 单叶面积Single leaf area | 4 | 7.296 | 0.005 |
| 比叶重Specific leaf weight | 4 | 0.760 | 0.575 |
降水量变化对叶水势、叶含水量、MDA含量、叶N含量、叶C含量和叶绿素含量都有显著影响 (图 2, 表 3)。

|
| 图 2 降水量变化对贝加尔针茅生理特征的影响 Fig. 2 Effects of precipitation change on physiological characters of S. baicalensis |
| 生理特征Physiological characteristics | df | F | P |
| 叶水势Leaf water potential | 4 | 15.056 | 0.000 |
| 叶含水量Leaf water content | 4 | 8.424 | 0.003 |
| MDA含量Malondialdehyde | 4 | 41.460 | 0.000 |
| 叶N含量Leaf N | 4 | 6.232 | 0.009 |
| 叶绿素Chlorophyll | 4 | 22.995 | 0.000 |
| 叶C含量Leaf C | 4 | 3.275 | 0.058 |
随着降水量的由多到少, 叶水势、叶含水量和叶C含量显著降低;MDA含量、叶N和叶绿素含量与降水量呈负相关关系。MDA含量和叶绿素含量处理组间变化最显著, 在降水从311 mm降到167 mm之间, 有3次显著变化, MDA含量分别出现在311—239、275—203 mm和203—167 mm之间, 叶绿素含量分别出现在311—275、275—239 mm和239—203 mm之间。叶水势、叶含水量和叶N含量分别出现两次显著变化, 叶水势分别出现在311—275 mm和275—167 mm之间, 叶含水量和叶N含量都是出现在311—203 mm和239—167 mm之间。叶C含量仅在275—167 mm之间有一个显著变化点, 其他降水水平之间差异不显著。
2.3 生物量特征随着降水量减少地上和地下生物量都呈现显著降低 (图 3, 表 4)。地上生物量变化比地下生物量变化更为敏感, 在311—275 mm和275—203 mm之间都出现了显著变化, 地下生物量在311—167 mm之间仅出现一次显著变化。

|
| 图 3 降水量变化对贝加尔针茅生物量的影响 Fig. 3 Effects of precipitation change on biomass accumulation of S. baicalensis |
| 生物量Biomass | df | F | P |
| 地上生物量Aboveground biomass | 4 | 42.971 | 0.000 |
| 地下生物量Belowground biomass | 4 | 4.328 | 0.027 |
总叶面积、单叶面积、株高、叶数、分蘖数、叶C含量、叶水势、叶含水量、地上和地下生物量与降水量呈正相关, 依据式 (3) 分别对其进行数据标准化;叶绿素含量、叶N含量、MDA含量与降水量呈负相关, 依据式 (4) 分别对其进行数据标准化。本研究是要确定干旱敏感指标的受旱临界点, 确定随降水量减少时指标值分布范围的容忍下限, 因此依据公式 (2) 对产生变化阈值的降水范围内数据进行计算, 采用95%的置信区间, 可得表 5中各个指标阈值。依据各指标与降水量的回归方程可求得对应的降水量。阈值是指敏感指标随降水量降低首次出现显著变化的点, 是判定植物受到胁迫的开始点, 按降水量阈值对各指标进行排序:总叶面积>地上生物量>叶水势=叶绿素含量>单叶面积>MDA含量>叶数>株高>分蘖数>叶含水量>叶N含量>地下生物量>叶C含量。形态特征方面总叶面积可以较好的反映降水量变化;生理特征可以通过测定水势 (或叶绿素含量) 反映植物水分状态;在生物量积累方面地上生物量对降水量变化的敏感性远大于地下生物量。
| 敏感指标 Sensitive indicators |
方程 Equation |
R2 | P | 阈值Threshold | 敏感程度 (Si) Sensitivity |
|
| 指标 Indicator |
降水量/mm Precipitation |
|||||
| MDA含量Malondialdehyde | y=-2.95E-05x2+0.0197x-2.308 | 0.91 | <0.01 | 0.84 | 264 | 0.003 |
| 总叶面积Total leaf area | y=2.79E-05x2-0.0074x+0.539 | 0.94 | <0.01 | 0.68 | 283 | 0.011 |
| 株高Plant heights | y=-2.15E-05x2+0.0156x-1.9262 | 0.87 | <0.01 | 0.61 | 247 | 0.006 |
| 叶绿素含量Chlorophyll | y=4.38E-05x2-0.0161x+1.6356 | 0.88 | <0.01 | 0.53 | 276 | 0.013 |
| 地上生物量Aboveground biomass | y=0.0058x-0.8855 | 0.89 | <0.01 | 0.74 | 280 | 0.008 |
| 叶水势Leaf water potential | y=3.00E-06x2+0.0034x-0.4952 | 0.83 | <0.01 | 0.67 | 276 | 0.009 |
| 叶数Leaf number | y=0.0061x-1.0054 | 0.90 | <0.01 | 0.57 | 258 | 0.008 |
| 叶片含水量Leaf water content | y=0.0048x-0.6222 | 0.77 | <0.01 | 0.47 | 227 | 0.006 |
| 分蘖数Tiller number | y=1.53E-05x2-0.0032x + 0.265 | 0.69 | <0.01 | 0.39 | 243 | 0.009 |
| 叶N含量Leaf N | y=0.0047x-0.6628 | 0.67 | <0.01 | 0.34 | 213 | 0.007 |
| 单叶面积Single leaf area | y=6.33E-05x2-0.0267x+2.9617 | 0.59 | <0.01 | 0.37 | 270 | 0.015 |
| 地下生物量Belowground biomass | y=0.004x-0.4599 | 0.52 | 0.01 | 0.37 | 207 | 0.006 |
| 叶C含量Leaf C | y=-3.39E-05x2+ 0.0197x-2.2259 | 0.50 | 0.01 | 0.27 | 186 | 0.006 |
指标敏感程度 (Si) 以出现降水胁迫开始点 (阈值) 时单位降水量导致该指标变化程度确定:
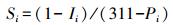
|
(5) |
式中Si为指标i对降水变化的敏感程度, Ii为指标i开始出现显著变化的值, Pi为指标i开始出现显著变化时对应的降水量, 311为降水增加30%对应的降水量。由表 5数据可知, 各指标的敏感程度由大到小依次为:单叶面积>叶绿素>总叶面积>水势=分蘖数>地上生物量>叶数>叶N含量>叶片含水量=地下生物量=株高=叶C含量>MDA含量。
2.5 敏感指标间关系降水量变化对本研究测定的形态、生理和生物量特征几乎都有显著的影响。指标间相关性分析显示叶N含量与叶绿素含量之间有着较强的相关关系 (图 4)。丙二醛 (MDA), 叶片含水量和叶水势这三者之间有着较好的相关关系 (图 4)。单叶面积和叶数的增多都可以显著的增加植株总叶面积 (图 4), 植株总叶面积的增加可以通过增加单叶面积和叶数两个途径来实现。利用通径分析方法可得植株总叶面积与叶数和单叶面积的回归方程:

|
| 图 4 贝加尔针茅响应降水量变化敏感指标间关系 Fig. 4 Relationships among indicators of S. baicalensis response to precipitation change |
总叶面积=-32.281+0.551叶数+59.244单叶面积
其相关系数和通径系数如表 6所示, 由表 6数据可知, 总叶面积的改变主要是叶数的变化引起。
| 自变量 Independent variable |
与总叶面积的相关关系 Correlation with total leaf area |
直接通径系数 Direct coefficient |
间接通径系数Indirect coefficient | |
| 叶数 Leaf number |
单叶面积 Single leaf area |
|||
| 叶数Leaf number | 0.96 | 0.78 | — | 0.18 |
| 单叶面积Single leaf area | 0.76 | 0.32 | 0.45 | — |
贝加尔针茅 (S. baicalensis) 广泛分布于中国松嫩平原、内蒙古高原东部, 向西南一直延伸到甘肃陇东高原和西秦岭山地, 是我国草甸草原的主要建群物种之一, 其分布区域气候为温凉半湿润型, 年降水量350—450 mm[2]。以往关于贝加尔针茅或草甸草原对降水 (干旱) 响应的阈值研究较少。侯琼等[6]基于SPAC (Soil-Plant-Atmosphere Continuum) 原理计算的草甸草原生长季 (6—8月) 需水量为270—290 mm。穆少杰等[35]运用BIOME-BGC模型计算贝加尔针茅草原年降水量为313.92 mm时NEP (net ecosystem productivity) 为0, 大于该值时NEP多为正值, 小于该值时NEP多为负值。周广胜等[36]基于MaxEnt模型计算得到贝加尔针茅潜在分布区域年降水量下限为308 mm。北方牧区草原干旱等级国家标准 (GB/T 29366—2012)[37]确定了判定温性草甸草原干旱的指标依次为土壤相对湿度、降水距平百分率和相对蒸降差, 并提供了相应的等级范围。这些研究大多运用模型的方法[35-36], 指标多采用年降水量[35-37], 草原年降水量时间分布不均, 多集中于生长季, 可达全年80%以上;评价多对于生物量或NEP[6, 35, 37], 其他性状多为定性分析, 阈值研究较少, 植物生长过程受到干旱胁迫会发生一系列变化, 首先反应到生理, 然后才是形态结构, 最终影响生物量。本研究以贝加尔针茅为材料, 从生理、形态和生物量3个方面筛选得到水势 (或叶绿素含量)、植株总叶面积和地上生物量4个干旱敏感指标, 受到胁迫时的降水量 (6、7和8月3个月份总降水量) 阈值分别是276 (276)、283 mm和280 mm, 与贝加尔针茅实际分布区生长季降水量下限较一致。针茅属植物 (Stipa L.) 受气候和土壤条件的地带性分异影响, 在各类针茅草原群落内部土壤养分分布、土壤水分分配等具空间异质性, 如半干旱草原土壤氮素具有小尺度格局, 这种异质性也会影响植物对降水变化的响应, 某一时期土壤水分含量通常难以与间隔距离较远区域比较, 但其土壤水分分布的结构特征主要受土壤性状的影响[38]。贝加尔针茅能适应多种基质的土壤, 黑钙土和栗钙土土壤都有其广泛分布[2], 本研究土壤为内蒙古草原典型栗钙土, 因此阈值适用方面应注意其土壤异质性。干旱指标有其相应的时空尺度, 在不同生长期指标及其阈值会存在变化[7, 15, 17, 39]。本研究探究植物对降水量变化的响应主要在个体水平进行, 缺少合理的机理解释, 且没有反应干旱累积效应。动态评估干旱发生时间, 揭示反映累积效应的机制, 以准确反映干旱累积效应, 提高对干旱的预测精度和及时性今后应给予重点关注[15, 17]。同时个体研究所得规律是否与自然生态系统相符, 有待于野外控制实验和长期定位观测数据的验证。
植物响应降水变化的生理生态指标多, 变异大, 给应用带来不便。降水量变化对本研究测定的形态、生理和生物量特征几乎都有显著的影响, 对这些指标间的相关性进行分析有利于指标的筛选和整合。本实验结果显示叶N含量与叶绿素含量之间有着较强的相关关系, N是组成叶绿素的必要成分, 较高的叶N含量对应较高的叶绿素含量[21, 40-41]。丙二醛 (MDA) 是植物体内脂质过氧化最终产物之一, 植物器官衰老或在逆境下遭受伤害, 往往发生膜脂过氧化作用, MDA含量可以反映植物遭受逆境伤害的程度[5]。干旱胁迫下叶片含水量减小, 叶水势降低, 同时会使活性氧累积, 从而导致膜脂过氧化反应增强, 膜脂遭到活性氧攻击后严重受损, 释放更多的MDA, 这3个指标间有着较好的相关关系[4, 5, 25, 28, 42-43]。植株总叶面积大小与叶数和单叶面积密切相关[4], 因此植株总叶面积的增加可以通过增加单叶面积和叶数两个途径来实现。利用通径分析方法发现总叶面积的改变主要是叶数的变化引起。从形态结构因素来看, 植株总叶面积、比叶重 (即单位叶面积重量) 都会导致生物量发生变化, 本研究结果显示降水量变化对贝加尔针茅比叶重没有产生显著影响, 所以降水量变化主要通过改变叶数引起植株总叶面积变化进而导致生物量变化。
4 结论本研究基于不同降水的模拟实验, 通过对贝加尔针茅形态、生理和生物量特征进行分析, 采用方差分析和正态总体统计容忍下限的方法找到了各敏感指标受降水胁迫的临界点, 并结合指标与降水量回归关系得到了对应的降水量阈值。按各指标开始受到降水变化胁迫的降水阈值对各指标进行排序为:总叶面积>地上生物量>叶水势=叶绿素含量>单叶面积>MDA含量>叶数>株高>分蘖数>叶含水量>叶N含量>地下生物量>叶C含量。研究表明, 在个体水平上, 植株总叶面积可做为贝加尔针茅形态特征方面响应降水变化的指示性指标, 其开始受降水胁迫的降水阈值为283 mm (6、7、8月3个月总降水量), 降水量变化导致总叶面积的改变主要与叶数而非单叶面积改变有关;叶水势或叶绿素含量可做为生理特征响应降水变化的指示性指标, 其开始受降水胁迫的降水阈值都为276 mm, 生物量特征方面地上生物量指示性高于地下生物量, 其开始受降水胁迫的降水阈值为280 mm。研究结果可为未来气候变化背景下草原植物对降水变化的响应与适应、监测与预测提供依据和参考。
致谢: 中国科学院植物研究所植被与环境变化国家重点实验室生态系统适应性研究组在实验过程中给予帮助, 特此致谢。| [1] | Bai Y F, Han X G, Wu J G, Chen Z Z, Li L H. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature, 2004, 431(7005): 181–184. DOI:10.1038/nature02850 |
| [2] | 张新时, 孙世洲, 雍世鹏. 中国植被及其地理格局——中华人民共和国植被图 (1 : 1000000) 说明书. 北京: 地质出版社, 2007. |
| [3] | 尹飞虎, 李晓兰, 董云社, 谢宗铭, 高志建, 何帅, 刘长勇. 干旱半干旱区CO2浓度升高对生态系统的影响及碳氮耦合研究进展. 地球科学进展, 2011, 26(2): 235–244. |
| [4] | 李永华, 卢琦, 吴波, 朱雅娟, 刘殿君, 张金鑫, 靳占虎. 干旱区叶片形态特征与植物响应和适应的关系. 植物生态学报, 2012, 36(1): 88–98. |
| [5] | Xu Z Z, Shimizu H, Ito S, Yagasaki Y, Zou C J, Zhou G S, Zheng Y R. Effects of elevated CO2, warming and precipitation change on plant growth, photosynthesis and peroxidation in dominant species from North China grassland. Planta, 2014, 239(2): 421–435. DOI:10.1007/s00425-013-1987-9 |
| [6] | 侯琼, 陈素华, 乌兰巴特尔. 基于SPAC原理建立内蒙古草原干旱指标. 中国沙漠, 2008, 28(2): 326–331. |
| [7] | 云文丽, 侯琼. 干旱指标在典型草原的适用性分析. 干旱区资源与环境, 2013, 27(8): 52–58. |
| [8] | 丁勇, 侯向阳, UbugunovL, 吴新宏, 尹燕亭, 李西良, 运向军. 温带草原区气候变化及其对植被影响的研究进展. 中国农学通报, 2012, 28(17): 310–316. DOI:10.11924/j.issn.1000-6850.2011-3571 |
| [9] | 马柱国, 符淙斌. 中国干旱和半干旱带的10年际演变特征. 地球物理学报, 2005, 48(3): 519–525. |
| [10] | Zhang L X, Zhou T J. Drought over East Asia: a review. Journal of Climate, 2015, 28(8): 3375–3399. DOI:10.1175/JCLI-D-14-00259.1 |
| [11] | Wu Z Y, Lu G H, Wen L, Lin C A. Reconstructing and analyzing China's fifty-nine year (1951-2009) drought history using hydrological model simulation. Hydrology and Earth System Sciences, 2011, 15(9): 2881–2894. DOI:10.5194/hess-15-2881-2011 |
| [12] | Battisti D S, Naylor R L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science, 2009, 323(5911): 240–244. DOI:10.1126/science.1164363 |
| [13] | IPCC. 2013: Summary for policymakers//Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M, eds. Climate Change 2013: The Physical Science Basis. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom: Cambridge University Press, 2013. |
| [14] | Heim Jr R R. Drought indices: a review//White D A, ed. Drought: A Global Assessment, Routledge Hazards and Disasters Series, Vol. 1. London: Routledge, 2000: 159-167. |
| [15] | 袁文平, 周广胜. 干旱指标的理论分析与研究展望. 地球科学进展, 2004, 19(6): 982–991. |
| [16] | Koleva E, Alexandrov V. Drought in the Bulgarian low regions during the 20th century. Theoretical and Applied Climatology, 2008, 92(1/2): 113–120. |
| [17] | 李柏贞, 周广胜. 干旱指标研究进展. 生态学报, 2014, 34(5): 1043–1052. |
| [18] | de Carvalho R C, Cunha A, da Silva J M. Photosynthesis by six Portuguese maize cultivars during drought stress and recovery. Acta Physiologiae Plantarum, 2011, 33(2): 359–374. DOI:10.1007/s11738-010-0555-1 |
| [19] | 张仁和, 郭东伟, 张兴华, 路海东, 刘建超, 李凤艳, 郝引川, 薛吉全. 吐丝期干旱胁迫对玉米生理特性和物质生产的影响. 作物学报, 2012, 38(10): 1884–1890. |
| [20] | Fonseca C R, Overton J M, Collins B, Westoby M. Shifts in trait-combinations along rainfall and phosphorus gradients. Journal of Ecology, 2000, 88(6): 964–977. DOI:10.1046/j.1365-2745.2000.00506.x |
| [21] | 侯彦会. 退化小针茅草原植物群落对水热变化的适应机理研究[D]. 北京: 中国科学院植物研究所, 2014. |
| [22] | Sampson D A, Allen H L. Regional influences of soil available water-holding capacity and climate, and leaf area index on simulated loblolly pine productivity. Forest Ecology and Management, 1999, 124(1): 1–12. DOI:10.1016/S0378-1127(99)00054-7 |
| [23] | Quétier F, Thébault A, Lavorel S. Plant traits in a state and transition framework as markers of ecosystem response to land-use change. Ecological Monographs, 2007, 77(1): 33–52. DOI:10.1890/06-0054 |
| [24] | 朱志梅, 杨持, 曹明明, 刘康. 草原沙漠化过程中植物叶面积的变化及其与土壤因子的关系. 生态学杂志, 2010, 29(12): 2384–2389. |
| [25] | 丁雷, 李英瑞, 李勇, 沈其荣, 郭世伟. 梯度干旱胁迫对水稻叶片光合和水分状况的影响. 中国水稻科学, 2014, 28(1): 65–70. |
| [26] | Siddique M R B, Hamid A, Islam M S. Drought stress effects on water relations of wheat. Botanical Bulletin of Academia Sinica, 2000, 41(1): 35–39. |
| [27] | 周婵, 杨允菲, 李建东. 松嫩平原两种趋异类型羊草对干旱胁迫的生理响应. 应用生态学报, 2002, 13(9): 1109–1112. |
| [28] | 李冬林, 向其柏. 土壤水分状况对浙江楠幼苗的影响. 南京林业大学学报:自然科学版, 2006, 30(5): 112–114. |
| [29] | 李金花. 内蒙古典型草原几种优势植物生态适应对策研究[D]. 兰州: 甘肃农业大学, 2002. |
| [30] | Jentsch A, Kreyling J, Boettcher-Treschkow J, Beierkuhnlein C. Beyond gradual warming: extreme weather events alter flower phenology of European grassland and heath species. Global Change Biology, 2009, 15(4): 837–849. DOI:10.1111/gcb.2009.15.issue-4 |
| [31] | Misson L, Degueldre D, Collin C, Rodriguez R, Rocheteau A, Ourcival J M, Rambal S. Phenological responses to extreme droughts in a Mediterranean forest. Global Change Biology, 2011, 17(2): 1036–1048. DOI:10.1111/gcb.2010.17.issue-2 |
| [32] | 牟成香, 孙庚, 罗鹏, 王志远, 罗光荣. 青藏高原高寒草甸植物开花物候对极端干旱的响应. 应用与环境生物学报, 2013, 19(2): 272–279. |
| [33] | Hernández J A, Almansa M S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiologia Plantarum, 2002, 115(2): 251–257. DOI:10.1034/j.1399-3054.2002.1150211.x |
| [34] | 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 3359-2009数据的统计处理和解释统计容忍区间的确定. 北京: 中国标准出版社, 2009. |
| [35] | 穆少杰, 周可新, 陈奕兆, 杨齐, 李建龙. 内蒙古典型草原不同群落净生态系统生产力的动态变化. 生态学杂志, 2014, 33(4): 885–895. |
| [36] | 周广胜, 何奇瑾, 殷晓洁. 中国植被/陆地生态系统对气候变化的适应性与脆弱性. 北京: 气象出版社, 2015. |
| [37] | 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 29366-2012北方牧区草原干旱等级. 北京: 中国标准出版社, 2013. |
| [38] | 白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落土壤水分和碳、氮分布的小尺度空间异质性. 生态学报, 2002, 22(8): 1215–1223. |
| [39] | 袁文平, 周广胜. 中国东北样带三种针茅草原群落初级生产力对降水季节分配的响应. 应用生态学报, 2005, 16(4): 605–609. |
| [40] | Karst A L, Lechowicz M J. Are correlations among foliar traits in ferns consistent with those in the seed plants?. New Phytologist, 2007, 173(2): 306–312. DOI:10.1111/j.1469-8137.2006.01914.x |
| [41] | 蒋高明. 植物生理生态学. 北京: 高等教育出版社, 2004. |
| [42] | Fu A H, Chen Y N, Li W H. Analysis on the change of water potential of Populus euphratica Oliv. and P. Russkii Jabl under different irrigation volumes in temperate desert zone. Chinese Science Bulletin, 2010, 55(10): 965–972. DOI:10.1007/s11434-009-0459-x |
| [43] | Sicher R C, Barnaby J Y. Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites to water stress. Physiologia Plantarum, 2012, 144(3): 238–253. DOI:10.1111/ppl.2012.144.issue-3 |