 2017, Vol. 37
2017, Vol. 37文章信息
- 陈云, 冯佳伟, 牛帅, 许宁, 韩军旺, 叶永忠, 袁志良
- CHEN Yun, FENG Jiawei, NIU Shuai, XU Ning, HAN Junwang, YE Yongzhong, YUAN Zhiliang.
- 小秦岭自然保护区苔藓植物群落数量分类、排序及多样性垂直格局
- Numerical classification, ordination, and species diversity of bryophytes along elevation gradients in Xiaoqinling
- 生态学报. 2017, 37(8): 2653-2664
- Acta Ecologica Sinica. 2017, 37(8): 2653-2664
- http://dx.doi.org/10.5846/stxb201512242568
-
文章历史
- 收稿日期: 2015-12-24
- 网络出版日期: 2016-10-19





2. 河南农业大生命科学学院, 郑州 450002;
3. 河南小秦岭国家级自然保护区管理局, 灵宝 472500
2. College of Life Sciences, Henan Agricultural Universit, Zhengzhou 450002, China;
3. Administration Bureau of Xiaoqinling National Nature Reserve of Henan, Lingbao 472500, China
苔藓是森林生态系统的重要组分, 在森林动态变化过程中扮演着重要的作用[1]。在林业上, 苔藓是划分森林类型的辅助指标, 并且被作为监测环境的指示植物[2]。然而苔藓植物个体相对较小, 缺乏广泛的经济价值, 因此很少成为人们关注的焦点, 其相关生态学研究也相当薄弱[3-4]。
植被群落分类和排序是植被生态学研究中最基本的问题[5]。目前, 常用的植被群落数量分类方法有聚类分析、双向指示种分析和多元回归树[6-7]。其中多元回归树是一种较新的群落数量分类方法, 较聚类分析和双向指示种分析更加客观[8]。目前围绕维管植物群落分类进行了较多的研究[9-11]。但对于苔藓植物分类, 通过查阅文献仅发现, 郭水良等首次在国内采用双向指示种分析, 对长白山苔藓植物进行了分类[2]。随后张元明等也采用双向指示种分析, 对新疆博格达山地地面生苔藓进行了分类[12]。苔藓植物分类研究较少, 并且没有发现采用多元回归树对苔藓植物进行划分的相关研究。
海拔作为地理梯度, 包含了温度、湿度和光照等各种环境因子, 被认为是植物物种、植物区系、植物群落和生态系统的决定因素[13]。山地植被物种多样性随海拔梯度的变化规律也一直是生态学家感兴趣的问题[14-15]。维管植物多样性沿海拔梯度的分布格局已有了大量的研究, 分布格局主要有5种形式, 分别是随海拔升高先降低后升高、先升高后降低、单调升高、单调下降和没有明显格局[14, 16]。然而苔藓物种多样性沿海拔梯度呈什么样的格局?在国内没有发现相关报道。
本研究基于53个20 m×20 m样地, 采集并鉴定了样地内所有生境中的苔藓物种, 采用MTR、CCA和GLM等分析方法, 探讨了以下问题:(1) 小秦岭地区苔藓植物组成, 以及优势科和优势种;(2) 综合分析苔藓植物之间及苔藓植物与地形因子之间的关系;(3) 苔藓物种多样性沿海拔梯度呈什么样的分布格局?与乔木、灌木和草本的分布格局是否一致?
1 材料与方法 1.1 研究区域概况小秦岭国家级自然保护区位于河南省灵宝市西部 (34°23′—34°31′N, 110°23′—110°44′E), 保护区东西长31 km, 南北宽12 km, 全区面积15160 hm2。森林覆盖率81.2%, 其主峰老鸦岔垴海拔2413.8 m, 为河南省最高峰。本区位于暖温带大陆性气候区, 属季风型半干旱气候, 年均气温11.2—14.2℃, 极端最低温度-17℃, 极端最高温度42.7℃, 无霜期170—215 d, 年降水量620 mm左右。
1.2 样地设置2012年, 在保护区主峰老鸦岔垴, 从海拔1020 m到2413 m设置样地。海拔每上升50 m, 选择代表性的群落分别设置样地, 样地大小为20 m×20 m, 共计53个样地。在样地内, 从地面到地上2 m范围内, 对样地内所有岩石、枯倒木、地面及木本植物个体上等所有苔藓进行采样, 样地中同一棵树或同一块石头的不同高度和方位的苔藓都需要采样, 而同一点位置上同种苔藓物种只采标本一份, 同一点位置不同种苔藓都要采集, 标本存放于信封中并记录样方号和生境类型。采集过程中仔细检查样地每个角落, 尽最大努力做到不遗漏, 共收集到苔藓标本1378份。所有标本带回实验室通过显微镜鉴定到种, 凭证标本存放于河南农业大学标本馆苔藓标本室。同时通过GPS测量每个样地的海拔, 地质罗盘测量坡度和坡向。
1.3 苔藓植物物种多度的计算物种多度一般指的是群落中某物种个体数量。传统的经典取样方法通常是在10 cm×10 cm、20 cm×20 cm或50 cm×50 cm等小尺度上进行采样[17-20]。然后通过不同的方法计算物种个体在样方内的物种个体数, 从而得到多度。这种方式对于研究小尺度上苔藓物种多样性、苔藓结皮及树木与附生苔藓之间的关系等方面效果较好。然而这种方法限制了在大尺度上苔藓生态学的发展。
为了有利于从宏观的角度来研究苔藓植物, 研究参考了贾宏汝对苔藓物种多度的处理方式[21], 即某一苔藓物种而言, 只关注该物种在样地中哪一棵树、哪一块石头或者哪一斑块地面出现了, 不关注它在该棵树、该块石头或者该斑块地面上同种个体的数量信息, 也就说该苔藓物种在某个点上有或没有, 从而得到苔藓的多度信息。
1.4 物种多样性指数计算计算每个样地内苔藓植物的生物多样性。
(1) Patrick丰富度指数
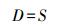
|
(1) |
(2)β多样性指数
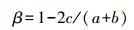
|
(2) |
式中, S为样地中的物种数目, a和b是两个样地各自所单独拥有的物种数, c是两样地间共有的物种数。
1.5 多元回归树本研究采用53个样地海拔、坡向和坡度为自变量, 240种苔藓植物在53个样地内的多度为因变量进行群丛分类。MRT采用R语言“mvpart”程序包进行运算[22]。
1.6 排序分析CCA分析中的物种矩阵选择苔藓物种在样地中的多度组成 (多度大于3的物种)。环境矩阵由海拔、坡向和坡度组成, 分析过程中对数据进行了标准化处理。通过R软件中envfit函数检验每个环境因子与苔藓物种分布的显著性, 模拟次数选择999次[23]。同时对CCA的最终结果进行蒙特卡罗置换检验, 分析模型是否达到显著水平 (P < 0.05)。CCA分析及显著性检验通过R语言软件中“Vegan”程序包实现[24]。
1.7 广义线性回归苔藓物种多样性作为响应变量, 地形因子作为解释变量。通过模型线性拟合来描述苔藓物种多样性随地形因子的变化格局。线性模型和作图通过R语言软件中“car”程序包实现[25]。
2 结果 2.1 苔藓物种多样性采集的1378份苔藓植物标本经过室内鉴定, 共有33科89属240种。其中藓类24科79属226种, 占总物种数的94.2%;苔类9科10属14种, 仅占总物种数的5.8%, 丰富度较低。53个样地苔藓物种多样性概况如表 1所示。小秦岭自然保护区苔藓植物的优势科和多度排前10位的物种概况如表 2所示。不同生境中的苔藓物种多样性差异如图 1所示。附表为小秦岭自然保护区苔藓植物物种名录和区系分布 (附表)。
| 样地编号 No. |
苔藓物种多样性 Bryophytes diversity |
苔类多样性 Liverworts diversity |
藓类多样性 Moss diversity |
科多样性 Families diversity |
属多样性 Generas diversity |
海拔 Elevation/m |
| 1 | 12 | 1 | 11 | 8 | 10 | 1020 |
| 2 | 8 | 0 | 8 | 6 | 6 | 1073 |
| 3 | 14 | 1 | 13 | 7 | 9 | 1143 |
| 4 | 14 | 1 | 13 | 8 | 9 | 1170 |
| 5 | 9 | 0 | 9 | 6 | 7 | 1176 |
| 6 | 5 | 0 | 5 | 4 | 4 | 1220 |
| 7 | 7 | 0 | 7 | 3 | 3 | 1230 |
| 8 | 13 | 0 | 13 | 5 | 7 | 1270 |
| 9 | 10 | 0 | 10 | 5 | 5 | 1282 |
| 10 | 16 | 0 | 16 | 7 | 12 | 1320 |
| 11 | 12 | 1 | 11 | 8 | 9 | 1320 |
| 12 | 16 | 2 | 14 | 9 | 11 | 1370 |
| 13 | 9 | 1 | 8 | 6 | 8 | 1370 |
| 14 | 20 | 0 | 20 | 8 | 12 | 1420 |
| 15 | 12 | 0 | 12 | 7 | 11 | 1430 |
| 16 | 20 | 0 | 20 | 7 | 11 | 1470 |
| 17 | 14 | 0 | 14 | 6 | 9 | 1470 |
| 18 | 12 | 0 | 12 | 5 | 8 | 1520 |
| 19 | 22 | 0 | 22 | 8 | 11 | 1530 |
| 20 | 14 | 0 | 14 | 9 | 11 | 1570 |
| 21 | 17 | 1 | 16 | 9 | 13 | 1585 |
| 22 | 13 | 1 | 12 | 10 | 11 | 1620 |
| 23 | 17 | 1 | 16 | 11 | 13 | 1635 |
| 24 | 7 | 0 | 7 | 3 | 3 | 1660 |
| 25 | 14 | 2 | 12 | 8 | 10 | 1670 |
| 26 | 2 | 0 | 2 | 2 | 2 | 1720 |
| 27 | 24 | 0 | 24 | 9 | 18 | 1730 |
| 28 | 16 | 0 | 16 | 7 | 11 | 1770 |
| 29 | 5 | 0 | 5 | 5 | 5 | 1779 |
| 30 | 21 | 1 | 20 | 9 | 10 | 1820 |
| 31 | 19 | 1 | 18 | 7 | 11 | 1830 |
| 32 | 14 | 1 | 13 | 10 | 12 | 1870 |
| 33 | 7 | 0 | 7 | 3 | 4 | 1870 |
| 34 | 29 | 2 | 27 | 14 | 23 | 1920 |
| 35 | 15 | 2 | 13 | 9 | 13 | 1923 |
| 36 | 19 | 1 | 18 | 9 | 13 | 1970 |
| 37 | 26 | 2 | 24 | 11 | 17 | 1970 |
| 38 | 7 | 0 | 7 | 5 | 5 | 2020 |
| 39 | 20 | 1 | 19 | 12 | 14 | 2020 |
| 40 | 10 | 0 | 10 | 7 | 10 | 2075 |
| 41 | 25 | 2 | 23 | 13 | 18 | 2080 |
| 42 | 23 | 1 | 22 | 13 | 18 | 2120 |
| 43 | 21 | 0 | 21 | 8 | 16 | 2186 |
| 44 | 9 | 1 | 8 | 7 | 9 | 2220 |
| 45 | 13 | 2 | 11 | 7 | 9 | 2284 |
| 46 | 26 | 4 | 22 | 13 | 17 | 2303 |
| 47 | 26 | 2 | 24 | 13 | 19 | 2315 |
| 48 | 11 | 0 | 11 | 6 | 7 | 2324 |
| 49 | 26 | 1 | 25 | 8 | 12 | 2339 |
| 50 | 17 | 2 | 15 | 8 | 12 | 2350 |
| 51 | 18 | 0 | 18 | 9 | 15 | 2378 |
| 52 | 21 | 1 | 20 | 12 | 15 | 2400 |
| 53 | 30 | 4 | 26 | 14 | 19 | 2413 |
| 编号 No. |
科名 Families |
属数 Generas |
种数 Species |
物种名 Species name |
多度 Abundance |
| 1 | 青藓科Bryaceae | 6 | 41 | 长肋青藓Brachythecium populeum | 96 |
| 2 | 提灯藓科Mniaceae | 4 | 24 | 灰白青藓Brachythecium albicans | 95 |
| 3 | 灰藓科Hypnaceae | 12 | 29 | 圆叶匐灯藓Plagiomnium vesicatum | 78 |
| 4 | 羽藓科Thuidiaceae | 5 | 19 | 宽叶青藓Brachythecium curtum | 74 |
| 5 | 丛藓科Pottiaceae | 14 | 23 | 大叶匐灯藓Plagiomnium succulentum | 61 |
| 6 | 绢藓科Entodontacae | 2 | 13 | 尖叶匐灯藓Plagiomnium acutum | 48 |
| 7 | 牛舌藓科Anomodontaceae | 4 | 10 | 平肋提灯藓Mnium laevinerve | 48 |
| 8 | 叶苔科Jungermanniaceae | 1 | 2 | 短肋羽藓Thuidium kanedae | 47 |
| 9 | 薄罗藓科Leskeaceae | 3 | 6 | 鳞叶藓Taxiphyllum taxirameum | 46 |
| 10 | 锦藓科Sematophyllaceae | 2 | 3 | 全缘匐灯藓Plagiomnium integrum | 41 |

|
| 图 1 不同生境中苔藓物种多样性差异 Fig. 1 Bryophytes diversity between different habitats 白色代表只在该生境中出现的物种数, 灰色代表在多个生境中出现的物种数 |
| 物种Species | 区系类型Floristic type |
| 薄囊藓Leptobryum pyriforme | 世界广布 |
| 兜叶细鳞苔Lejeunea cavifolia | 北温带分布 |
| 反纽藓Timmiella anomala | 旧世界温带分布 |
| 亮叶绢藓Entodon aeruginosus | 中国特有 |
| 拟三列真藓Bryum pseudotriquetrum | 温带地区广布 |
| 圆枝青藓Brachythecium garovaglioides | 东亚分布 (中-日) |
| 白氏藓brotherus leana | 中国广布 |
| 全缘匐灯藓Plagiomnium integrum | 热带亚洲分布 |
| 仙鹤藓多蒴变种Atrichum var. gracilisetum besch | 北半球广布 |
| 侧枝匐灯藓Plagiomnium maximovicgii | 北温带分布 |
| 大凤尾藓Fissidens nobilis | 温带亚洲分布 |
| 毛尖青藓Brachythecium piligerum | 东亚分布 (中-日) |
| 青藓Brachythecium pulchellum | 东亚分布 (中-日) |
| 网孔凤尾藓Fissidens areolatus | 温带亚洲分布 |
| 褶叶小墻藓Weisiopsis anomala | 东亚分布 (中-日) |
| 东亚小石藓Weisia exserta | 东亚分布 |
| 具丝毛灯藓Rhizomnium tuomikoskii | 东亚分布 (中-日) |
| 绿枝青藓Brachythecium | 中国特有 |
| 平叶原鳞苔Archilegeunea planiuscula | 热带亚洲至热带大洋洲分布 |
| 小胞仙鹤藓Atrichum rhystophyllum | 东亚分布 (中-日) |
| 八齿碎米藓Fabronia ciliaris | 中国特有 |
| 匐枝青藓Brachythecium procumbens | 东亚分布 |
| 大叶匐灯藓Plagiomnium succulentum | 温带亚洲分布 |
| 尖叶青藓Brachythecium coreanum | 东亚分布 |
| 阔叶丛本藓Anoectangium clarum | 温带亚洲分布 |
| 齿边同叶藓Isopterygium serrulatum | 温带亚洲分布 |
| 羽枝青藓Brabhythecium piumosum | 北温带分布 |
| 东亚仙鹤藓Atrichum yakushimense | 东亚分布 |
| 绿叶绢藓Entodon viridulus | 东亚分布 (中-日) |
| 柔叶青藓Brachythecium moriense | 东亚分布 (中-日) |
| 曲枝青藓Brachythecium dicranoides | 中国特有 |
| 小酸土藓Oxystegus cuspidatus | 温带亚洲分布 |
| 异形凤尾藓Fissidens anomalus | 北温带亚洲 |
| 银藓Anomobryum filiforme Solms | 世界广布 |
| 粗肋薄罗藓Leskea scabrinervis | 中国特有 |
| 毛尖羽藓Thuidium philibertii | 亚热带山区 |
| 狭叶湿地藓Hyophila stenophylla | 东亚分布 (中-日) |
| 毛尖燕尾藓Bryhnia trichomitria | 东亚分布 (中-日) |
| 球根真藓Bryum radiculosum | 世界广布 (除澳洲) |
| 羽枝美喙藓Eurbynchuium longirameum | 中国特有 |
| 皱叶小石藓Weisia controversa | 世界广布 |
| 南亚同叶藓Isopterygium bancanum | 温带亚洲分布 |
| 凸尖鳞叶藓Taxiphyllum cuspidifolium | 东亚-北美间断分布 |
| 原叶细鳞苔Lejeunea sordida | 世界广布 |
| 匐灯藓Plagiomnium cuspidatum | 北温带分布 |
| 柳叶藓原变种Amblystegium serpens | 世界广布 |
| 细枝薄罗藓Leskea gracilescens | 东亚-北美间断分布 |
| 中华附干藓Schwetschkea sinica | 中国特有 |
| 华东反齿藓Anacamptodon fortunei mitt | 东亚分布 (中-日) |
| 泛生丝瓜藓Pohlia cruda | 世界广布 |
| 薄边毛灯藓Rhizomnium horikawae | 温带亚洲分布 |
| 毛齿匐灯藓Plagiomnium tezukae | 东亚分布 (中-日) |
| 小牛舌藓全缘亚种Andmodon minor var. integerrimus | 温带亚洲分布 |
| 粗枝藓Gollania neckerella | 东亚分布 (中-日) |
| 小树平藓Homaliodendron exiguum | 北温带亚洲 |
| 拟附干藓Schwetschkeopsis fabronia | 东亚-北美间断分布 |
| 小凤尾藓Fissidens bryoides | 北半球广布 |
| 小叶美喙藓Eurbynchuium filiforme | 中国特有 |
| 短枝燕尾藓Bryhnia brachycladula | 东亚分布 (中-日) |
| 皱叶牛舌藓Anomodon dentatus | 北温带分布 |
| 燕尾藓Bryhnia nocae-angliae | 北温带分布 |
| 光萼叶苔Jungermannia leiantha | 北温带分布 |
| 卷叶凤尾藓Fissidens cristatus | 旧世界温带分布 |
| 柔软明叶藓Vesicularia flaccida | 东亚分布 (中-日) |
| 耳叶丛藓Pottia truncata | 中国特有 |
| 异叶小羽藓Haplocladium larminatii | 东亚分布 |
| 多褶耳叶苔Frullania polyptera | 热带亚洲分布 |
| 长柄绢藓Entodon macropodus | 世界广布 |
| 多毛细羽藓Cyto-hypnum vestitissimum | 北温带分布 |
| 拟同叶藓Isopterygiopsis muelleriana | 世界广布 |
| 温带灰藓强弯亚种Hypnum subimponens | 东亚分布 (中-日) |
| 斜蒴藓Camptothecium | 世界广布 |
| 大麻羽藓Claopodium assurgens | 亚热带低山区 |
| 细麻羽藓Claopodium gracillimum | 东亚分布 |
| 狭叶仙鹤藓Atrichum angustatum | 中国、欧洲、北美洲 |
| 链齿藓Desmatodon latifolius | 中国特有 |
| 钝叶牛舌藓Anomodon rotundatus | 中国特有 |
| 深绿绢藓Entodon luridus | 东亚分布 |
| 陈氏耳叶苔Frullania chenii | 中国特有 |
| 赤根青藓Brachythecium dicranoides | 旧世界温带分布 |
| 多疣细羽藓Cyrto-hypnum pygmaeum | 东亚-北美间断分布 |
| 毛灯藓Rhizomnium punctatum | 北温带分布 |
| 尖毛细羽藓Cyto-hypnum fuscatum | 温带亚洲分布 |
| 细枝藓Lindbergia brachyptera | 北温带分布 |
| 东亚拟鳞叶藓Pseudotaxiphyllum pohliaecarpum | 东亚分布 (中-日) |
| 卷叶毛口藓TrichostomumBruch brachydontiumBruch | 中国特有 |
| 狭叶长喙藓Rhynchostegium fauriei | 东亚分布 (中-日) |
| 延叶凤尾藓Fissidens perdecurrens | 东亚分布 (中-日) |
| 美丽长喙藓Rhynchostegium subspeciosum | 中国特有 |
| 尖叶美喙藓Eurhynchium eustegium | 东亚分布 (中-日) |
| 提灯藓Mnium hornum | 世界广布 |
| 中华裂萼苔Chiloscyphus sinensis | 中国特有 |
| 薄罗藓Leskea polycarpa | 北温带分布 |
| 台湾棉藓原变种Plagiothecium formosicum | 中国特有 |
| 短枝羽藓Thuidium submicropteris | 东亚分布 (中-日) |
| 瘤根真藓Bryum bornholmense | 中国、欧洲 |
| 狭叶扭口藓Barbula subcontorta | 北温带亚洲 |
| 金灰藓Pylaisiellapolyantha | 北温带分布 |
| 瓦叶藓Miyabea frutcella | 东亚分布 (中-日) |
| 细湿藓原变种Campylium hispidulum | 世界广布 |
| 具喙匐灯藓Plagiomnium rhynchophorum | 泛热带分布 |
| 水生长喙藓Rhynchostegium riparioides | 北半球广布 |
| 橙色锦藓Sematophyllum phoeniceum | 热带亚洲-热带非洲分布 |
| 刺叶提灯藓Mnium spinosum | 世界广布 |
| 密毛细羽藓Cyrto-hypnum gratum | 旧世界热带分布 |
| 脆枝曲柄藓原变种campylopus fragilis | 北温带分布 |
| 蛇苔Conocephalum conicum | 北半球广布 |
| 酸土藓Oxystegus cuspidatus | 世界广布 |
| 淡色同叶藓Isopterygium albescens | 温带亚洲分布 |
| 皱叶麻羽藓Claopodium rugulosifolium | 中国特有 |
| 侧立藓pleuroweisia schliephackei | 欧亚大陆共有 |
| 尖叶提灯藓Plagiomnium acutum | 北温带分布 |
| 异叶提灯藓Mnium heterophyllum | 北温带分布 |
| 东亚金灰藓Pylaisiella brotheri | 东亚分布 |
| 偏蒴藓Ectropothecium buitenzorgii | 热带亚洲分布 |
| 带叶牛舌藓Anomodon perlingulatus | 中国特有 |
| 墙藓Tortula subulata | 北温带分布 |
| 短尖美喙藓Eurhynchium angustirete | 东亚分布 |
| 褶叶小墙藓Hyophila anomala | 东亚分布 (中-日) |
| 疣灯藓Trachycystis microphylla | 东亚分布 |
| 大金灰藓Pylaisiella robusta | 东亚分布 (中-日) |
| 短肋羽藓Thuidium kanedae | 东亚分布 (中-日) |
| 灰白青藓Brachythecium albicans | 北温带分布 |
| 鳞叶藓Taxiphyllum taxirameum | 热带亚洲至热带美洲分布 |
| 圆叶匐灯藓Plagiomnium vesicatum | 旧世界温带分布 |
| 长肋青藓Brachythecium populeum | 北温带分布 |
| 钝叶绢藓Entodon obtusatus | 东亚分布 |
| 细叶小羽藓Haplocladium microphyllum | 北温带分布 |
| 北方金灰藓Pylaisiella sewynii | 北温带分布 |
| 脆枝青藓Brachythecium thraustum | 中国特有 |
| 短叶毛锦藓Pylaisiadelpha yokohamae | 东亚分布 (中-日) |
| 密枝青藓Brachythecium amnicola | 中国特有 |
| 疏网美喙藓Eurbynchuium laxirete | 东亚分布 (中-日) |
| 圆叶裂萼苔chiloscyphus horikawana | 东亚分布 (中-日) |
| 大羽藓Thuidium cymbifolium | 东亚分布 |
| 娇美绢藓Entodon pulchellus | 东亚分布 |
| 宽叶青藓Brachythecium curtum | 世界广布 |
| 扭尖美喙藓Eurhynchium kirishimense | 东亚分布 (中-日) |
| 陕西鳞叶藓Taxiphyllum giraldii | 东亚分布 (中-日) |
| 长帽绢藓Entodon dolichocucullatus | 中国特有 |
| 东亚缩叶藓Ptychomitrium fauriei | 东亚分布 |
| 美灰藓Eurohypnum leptothallum | 温带亚洲分布 |
| 灰青藓Brachythecium glauculum | 中国特有 |
| 长叶提灯藓Mnium lycopodioides | 世界广布 |
| 平肋提灯藓Mnium laevinerve | 温带亚洲分布 |
| 威氏缩叶藓Ptychomitrium wilsonii | 东亚分布 (中-日) |
| 圆叶提灯藓Plagiomnium vesicatum | 东亚分布 (中-日) |
| 尖叶匐灯藓Plagiomnium acutum | 温带亚洲分布 |
| 绒叶青藓Brachythecium uncinifolium | 北温带分布 |
| 剑叶藓Merceyopsis sikkimensis | 热带亚洲分布 |
| 日本细喙藓Rhynchostegiella japonica | 东亚分布 (中-日) |
| 斜枝青藓Brachythecium campylothallum | 中国广布 |
| 齿边细枝藓Lindbergia serrulatus | 中国特有 (西南成分) |
| 叉肋藓Teachyphyllum inflexum | 旧世界热带分布 |
| 横生绢藓Entodon prorepens | 东亚分布 |
| 拟圆叶剪叶苔Jungermannia (S.) pseudocyclops Inoue | 中国特有 |
| 芽胞湿地藓HyophilaBrid javanica | 东亚分布 (中-日) |
| 扭尖美喙藓Eurhynchium kirishimense Takaki | 东亚分布 (中-日) |
| 小细柳藓Platydictya subtilis | 世界广布 |
| 真藓Bryum argenteum | 世界广布 |
| 密根多毛藓Lescuraea radicosa | 北温带分布 |
| 暖地明叶藓Vesicularia ferriei | 东亚分布 (中-日) |
| 小仙鹤藓Atrichum henryi | 东亚分布 (中-日) |
| 暗绿多枝藓Haplohymenium triste | 北温带分布 |
| 宽叶美喙藓Eurhynchium hians | 旧世界温带分布 |
| 细柳藓Platydictya jungermannioides | 北温带分布 |
| 芽孢湿地藓Hyophila propagulifera | 东亚分布 (中-日) |
| 短月藓Brachymenium nepalense | 热带亚洲至热带非洲分布 |
| 剑叶大帽藓Encalypta apathulata | 世界广布 |
| 陕西灰藓Hypnum shensianum | 中国特有 |
| 匐枝长喙藓Rhynchostegium serpenticaule | 中国特有 |
| 密集匐灯藓Plagiomnium conferteidens | 旧世界温带分布 |
| 羊角藓Herpetineuron toccoae | 热带亚洲至热带美洲分布 |
| 大墙藓Tortula princeps | 世界广布 |
| 多疣麻羽藓Claopodium pellucinerve | 东亚分布 |
| 细湿藓稀齿变种Campylium hispidulum | 世界广布 |
| 小凤尾藓原变种Fissideus bryoides var. bryouides | 北温带分布 |
| 羽叶凤尾藓Fissidens plagiochloides | 东亚分布 |
| 土生真藓Bryum tuberosum | 东亚分布 |
| 树形疣灯藓Trachycystis ussuriensis | 温带亚洲分布 |
| 直叶凤尾藓Fissidens strictulus | 东亚分布 |
| 尖叶绢藓Entodon acutifolius | 中国特有 |
| 牛舌藓Anomodon viticulosus | 旧世界温带分布 |
| 丛生真藓Bryum caespiticium | 世界广布 |
| 皮叶苔Targionia hypophylla | 世界广布 |
| 羽状青藓Brachythecium plumosum | 北半球广布 |
| 狭叶小羽藓Haplocladium angustifolium | 旧世界温带分布 |
| 丛本藓Anoectangium aestivum | 北温带分布 |
| 密叶美喙藓Eurhynchium savatieri Schimp | 东亚分布 (中-日) |
| 扭叶丛本藓Anoectangium stracheyanum | 温带亚洲分布 |
| 狭叶拟绢藓Entodontopsis wightii | 北温带亚洲 |
| 大粗枝藓Gollania robusta | 中国特有 |
| 多变粗枝藓Gollania varians | 东亚分布 |
| 狭叶美喙藓Eurhynchium coarctum | 中国特有 |
| 紫色细湿藓Campylium prophyriticum | 中国特有 |
| 齿边缩叶藓Ptychomitrium dentatum | 东亚分布 |
| 拟小凤尾藓Fissidens tosaensis | 东亚分布 (中-日) |
| 小牛舌藓原变种Andmodon minor var. minor | 东亚-北美间断分布 |
| 长叶绢藓Entodon longifolius | 热带亚洲分布 |
| 虫毛藓Boulaya mittenii | 东亚分布 |
| 高山红叶藓Bryoerythrophyllum alpigenum | 旧世界温带分布 |
| 褶叶青藓Brachythecium salebrosum | 北温带分布 |
| 细枝羽藓Thuidium delicatulum | 世界广布 |
| 具缘提灯藓Mnium marginatum | 世界广布 |
| 短叶小石藓WeisiaHedw controversaHedw | 中国特有 |
| 毛口藓Trichostomum brachydontium | 世界广布 |
| 斜枝长喙藓Rhynchostegium inclinatum | 东亚分布 (中-日) |
| 长尖明叶藓Vesicularia reticulata | 热带亚洲分布 |
| 丝瓜藓Pohlia nutans | 世界广布 |
| 绿叶绢藓Entodon viridulus | 东亚分布 |
| 狭边大叶藓Rhodobryum ontariense | 北温带亚洲 |
| 皱叶粗枝藓Gollania ruginosa | 温带亚洲分布 |
| 垂蒴棉藓Plagiothecium nemorale | 热带亚洲分布 |
| 长尖扭口藓Barbula unguiculata | 中国特有 |
| 阔叶桧藓Pyrrhobryum latifolium | 热带亚洲至热带美洲分布 |
| 陈氏唇鳞苔Cheiloeunea chenii | 中国特有 |
| 阔边大叶藓Rhodobryum giganteum | 温带亚洲分布 |
| 木藓Thamnobryum subseriatum | 东亚分布 |
| 细罗藓Leskeella nervosa | 北温带分布 |
| 糙叶美喙藓Eurhynchium squarrifolium | 东亚分布 (中-日) |
| 戟叶梳藓Ctenidium hastile | 东亚分布 (中-日) |
| 中华粗枝藓Gollania sinensis | 中国特有 |
| 东亚小羽藓Haplocladium strictulum | 东亚分布 (中-日) |
| 短萼叶苔Jungermannia breviperianthia | 中国特有 |
| 中华细枝藓Lindbergia sinensis | 中国特有 |
| 锐齿短萼苔Wettsteinia inversa | 东南亚分布 |
| 树状美喙藓Eurbynchuium arbuscula | 东亚分布 (中-日) |
| 皱叶绢藓Entodon plicatus | 东亚分布 |
| 扭叶镰刀藓Drepanocladus revolvens | 北半球寒温带 |
| 弯叶灰藓Hypnum hamulosum | 欧亚大陆共有 |
| 钝叶平蒴藓Plagiobrym giraldii | 中国特有 |
| 弯叶青藓Brachythecium reflexum | 北温带分布 |
| 皱叶匐灯藓Plagiomnium arbusculum | 东亚分布 (中-日) |
| 芽胞裂萼苔Chiloscyphus minor | 北半球广布 |
| 湿柳藓Hygroamblystegium tenax | 世界广布 |
| 红毛细羽藓Cyto-hypnum versicolor | 中国特有 |
| 鞭枝多枝藓Haplohymenium Flagelliforme | 温带亚洲分布 |
| 短尖燕尾藓Bryhnia hultenii | 东亚分布 (中-日) |
| 楔形耳叶苔Frullania inflexa | 东亚分布 |
MRT能兼顾误差, 又能保证树的规模足够小[8], 以二岐式分割法将53个样地分类, 经过交叉验证, 结合宋永昌关于群丛的命名原则[26], 认为可以分为4类 (图 2)。

|
| 图 2 小秦岭自然保护区苔藓植物MRT分类 Fig. 2 Dendrogram of the multivariate regression trees classification of the bryophyte community in Xiaoqinling Nature Reserve |
Ⅰ长肋青藓 (Brachythecium populeum)+大叶匐灯藓 (Plagiomnium succulentum)+圆叶匐灯藓 (Plagiomnium vesicatum) 群落, 包括1—35号样地, 该群落主要分布在海拔1020—1923 m。
Ⅱ灰白青藓 (Brachythecium albicans)+宽叶青藓 (Brachythecium curtum)+短肋羽藓 (Thuidium kanedae)+平肋提灯藓 (Mnium laevinerve) 群落, 包括42—53号样地, 该群落主要分布在海拔1970—2080 m的地带。
Ⅲ短肋羽藓 (Thuidium kanedae)+光萼叶苔 (Jungermannia leiantha)+薄罗藓 (Leskea polycarpa)+叉肋藓 (Trachyphyllum inflexum) 群落, 包括36, 37, 38和40号样地, 该群落主要分布在海拔1970—2075 m。
Ⅳ宽叶青藓 (Brachythecium curtum)+垂蒴棉藓 (Plagiothecium nemorale)+全缘匐灯藓 (Plagiomnium integrum) 群落, 包括39和41号样地, 该群落分布在海拔2020—2080 m。
2.3 排序结果对排序结果进行蒙特卡罗检验, 显著性小于0.05, 说明排序的结果可以接受地形因子对物种分布的解释量, 排序结果如图 3所示。每个环境因子的显著性如表 3所示。海拔、坡度和坡向三者的解释量为7.29%, 其中第一轴的特征值为21.85%, 第二轴的特征值为15.83%。苔藓在排序轴上的分布基本反映了植物空间分布特征随地形因子的变化趋势。

|
| 图 3 苔藓物种分布与地形因子的关系 Fig. 3 Relationship between topography factors and bryophyte species distribution 图中圆点和三角形分别代表物种和样地 |
| 环境因子Environmental factors | CCA1 | CCA2 | R2 | Pr(>r) |
| 海拔Elevation | 0.99704 | -0.07683 | 0.7041 | 0.001*** |
| 坡度Slope | -0.12824 | -0.99174 | 0.5145 | 0.001*** |
| 坡向Aspect | 0.51398 | -0.85780 | 0.2324 | 0.008** |
苔藓物种丰富度与海拔表现出极显著相关 (P<0.001), 而与坡度和坡向未能达到显著水平;苔藓β多样性与海拔、坡度和坡向均未达到显著水平。苔藓物种多样性和β多样性随海拔的变化格局分别如图 4所示。苔藓物种多样性随海拔的变化成明显的上升格局, 而苔藓β多样性随海拔的变化呈水平变化。

|
| 图 4 苔藓物种多样性沿海拔分布格局 Fig. 4 distribution pattern of bryophyte species diversity along elevation gradients |
每种苔藓物种 (个体数>3) 的海拔分布范围如图 5所示。长肋青藓、灰白青藓、圆叶匐灯藓和宽叶青藓等物种的海拔分布极为广泛, 在海拔1020—2378 m都有分布, 而丛本藓 (Anoectangium aestivum)、狭叶小羽藓 (Haplocladium angustifolium) 和灰白青藓 (Brachythecium albicans) 等物种海拔分布范围较窄。

|
| 图 5 小秦岭自然保护区苔藓物种的海拔分布范围 Fig. 5 The elevation range of bryophyte species distribution in Xiaoqinling nature reserve |
通过对小秦岭地区苔藓植物的调查研究表明, 该保护区的苔藓植物比较丰富。苔类植物总数远小于藓类, 这与汪岱华等的研究结果是一致的[27]。青藓科 (Bryaceae)、提灯藓科 (Mniaceae)、灰藓科 (Hypnaceae) 和羽藓科 (Thuidiaceae), 该4科的物种数占总物种数的47%, 是小秦岭国家自然保护区的主要类群。长肋青藓 (Brachythecium populeum)、灰白青藓 (Brachythecium albicans)、圆叶匐灯藓 (Plagiomnium vesicatum) 和宽叶青藓 (Brachythecium curtum) 为小秦岭自然保护区的常见种。
本次研究结果显示, 不同的生境中苔藓多样性差异较大, 在样地中约有57.5%物种只倾向于分布某一类生境中, 这就表明生境异质性有助于丰富苔藓物种多样性, 这也是在有些森林实施管理中要保持生境类型多样性的主要原因。
基于地形因子和物种组成数据的MRT技术将小秦岭自然护区53块样地的240种苔藓分成了4类, 结果显示4个群落都有各自的物种组成和群落特征。通过CCA分析表明海拔、坡度和坡向显著影响着苔藓物种分布。不同的海拔梯度上温度、降水及空气湿度等存在很大差异[28], 不同坡度和坡向的土壤水分和养分含量也存在一定的差异[29-30]。因此, 海拔、坡度和坡向3个地形因子显著影响着苔藓植物的分布。但三者的解释量只有7.29%, 解释量较低, 并且影响苔藓物种分布的环境因子也较多, 今后的研究应逐步考虑气候条件、树皮的理化性质及光照强度等更多的环境因子。
本研究结果表明小秦岭地区苔藓物种多样性随海拔的上升呈显著的上升格局。Stehn等在阿巴拉契亚山区研究也表明苔藓物种多样性随海拔的上升呈上升格局[31], 本研究结果与其是一致。陈云等在小秦岭地区研究了乔木、灌木和草本随海拔的变化格局, 结果显示乔木层物种丰富度呈单峰曲线, 灌木层的丰富度在中海拔呈下降趋势, 草本层物种丰富度沿海拔梯度呈下降趋势[32]。苔藓物种与乔木、灌木和草本随海拔的变化呈现出不一样的格局, 这就表明苔藓物种生长所需要的环境条件和乔木、灌木、草本是有很大差异。海拔对苔藓β多样性的影响未能达到显著水平, 这可能是因为苔藓个体小, 受局部微生境影响较大。
以上分析已表明海拔显著影响着苔藓植物的分布, 那么每种苔藓物种在海拔上是怎样分布的?结果表明不同苔藓的海拔生长范围存在较大差异, 特别是一些海拔分布范围较窄的物种, 说明这些物种对环境适应能力差, 因此苔藓物种保护过程中应多关注这些物种, 并且要针对不同物种和不同海拔段实施不同的保护措施。
总的来说, 通过本研究了解了小秦岭国家级自然保护区苔藓物种组成、分类以及每种苔藓植物的海拔分布范围;苔藓物种多样性随海拔的上升呈显著的上升格局, 与乔木、灌木和草本的变化格局不同。希望此研究能为苔藓植物生态学研究和物种多样性保护提供参考资料。然而, 维管植物和苔藓共存于一个生态系统中, 本文未能考虑它们对苔藓群落的影响, 当然, 维管植物与苔藓群落之间的关系也是非常复杂的, 维管植物究竟怎样影响着苔藓群落?例如, 乔木、灌木和草本分别是如何影响苔藓物种分布的?不同生境中的苔藓 (树生苔藓、地面生苔藓、石生苔藓等) 与维管植物之间存在怎样的关系?这些问题都有待于深入研究。
致谢: 中国科学院植物研究所贾渝老师帮助苔藓鉴定,特此致谢。| [1] | 雷波, 包维楷, 贾渝. 6种人工针叶幼林下地表苔藓植物层片的物种多样性与结构特征. 植物生态学报, 2004, 28(5): 594–600. DOI:10.17521/cjpe.2004.0079 |
| [2] | 郭水良, 曹同. 长白山地区森林生态系统树附生苔藓植物群落分布格局研究. 植物生态学报, 2000, 24(4): 442–450. |
| [3] | 叶吉, 郝占庆, 于德永, 闫海冰, 封德全. 苔藓植物生态功能的研究进展. 应用生态学报, 2004, 15(10): 1939–1942. DOI:10.3321/j.issn:1001-9332.2004.10.047 |
| [4] | 姜炎彬, 邵小明. 苔藓植物分布及其物种多样性的研究评述. 武汉植物学研究, 2010, 28(3): 385–390. |
| [5] | 吴征镒. 中国植被. 北京: 科学出版社, 1980: 1–1375. |
| [6] | 韩军旺, 王进, 张旭, 贾宏汝, 袁志良. 小秦岭国家级自然保护区植物群落数量分类与排序. 河南科学, 2015, 33(4): 547–552. |
| [7] | 张文静, 张钦弟, 王晶, 冯飞, 毕润成. 多元回归树与双向指示种分析在群落分类中的应用比较. 植物生态学报, 2015, 39(6): 586–592. DOI:10.17521/cjpe.2015.0056 |
| [8] | 赖江山, 米湘成, 任海保, 马克平. 基于多元回归树的常绿阔叶林群丛数量分类-以古田山24公顷森林样地为例. 植物生态学报, 2010, 34(7): 761–769. |
| [9] | De'ath G. Multivariate regression trees: a new technique for modeling species-environment relationships. Ecology, 2002, 83(4): 1105–1117. |
| [10] | 张荣, 刘彤. 古尔班通古特沙漠南部植物多样性及群落分类. 生态学报, 2012, 32(19): 6056–6066. |
| [11] | 黄永涛, 姚兰, 艾训儒, 吕世安, 丁易. 鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析. 植物生态学报, 2015, 39(10): 990–1002. DOI:10.17521/cjpe.2015.0096 |
| [12] | 张元明, 曹同, 潘伯荣. 新疆博格达山地面生苔藓植被的数量分类与排序研究. 植物生态学报, 2002, 26(1): 10–16. |
| [13] | Pescador D S, de Bello F, Valladares F, Escudero A. Plant trait variation along an altitudinal gradient in Mediterranean high mountain grasslands: controlling the species turnover effect. PLoS One, 2015, 10(3). |
| [14] | 朱源, 康慕谊, 江源, 刘全儒. 贺兰山木本植物群落物种多样性的海拔格局. 植物生态学报, 2008, 32(3): 574–581. |
| [15] | Koenig I, Feldmeyer-Christe E, Mitchell E A D. Comparative ecology of vascular plant, bryophyte and testate amoeba communities in four Sphagnum peatlands along an altitudinal gradient in Switzerland. Ecological Indicators, 2015, 54: 48–59. DOI:10.1016/j.ecolind.2015.01.043 |
| [16] | He J S, Chen W L. A review of gradient changes in species diversity of land plant communities. Acta Ecologica Sinica, 1997, 17(1): 91–99. |
| [17] | 蔺菲, 郝占庆, 李步杭, 叶吉, 戴冠华, 张健, 倪伟东. 长白山暗针叶林苔藓植物群落特征与林木更新的关系. 生态学报, 2007, 27(4): 1308–1314. |
| [18] | 刘蔚秋, 戴小华, 王永繁, 雷纯义. 影响广东黑石顶树附生苔藓分布的环境因子. 生态学报, 2008, 28(3): 1080–1088. |
| [19] | 王中生, 安树青, 方炎明. 苔藓植物生殖生态学研究. 生态学报, 2003, 23(11): 2444–2452. DOI:10.3321/j.issn:1000-0933.2003.11.029 |
| [20] | 福英, 白学良, 张乐, 毕庚辰, 冯超, 寇瑾, 萨如拉. 五大连池火山熔岩地貌苔藓植物对土壤养分积累的作用. 生态学报, 2015, 35(10): 3288–3297. |
| [21] | 贾宏汝. 环境空间异质性对暖温带-北亚热带过渡区植物群落构建的影响[D]. 郑州: 郑州大学, 2015. |
| [22] | De'Ath G. mvpart: multivariate partitioning. R package version 1.2-4. 2006. |
| [23] | Patiño J, González-Mancebo J M. Exploring the effect of host tree identity on epiphyte bryophyte communities in different Canarian subtropical cloud forests. Plant Ecology, 2011, 212(3): 433–449. DOI:10.1007/s11258-010-9835-5 |
| [24] | MHH W H. Vegan: Community Ecology Package. R package version, 2-0. 2011. |
| [25] | Fox J, Weisberg S. Using car functions in other functions. 2013. |
| [26] | 宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001. |
| [27] | 汪岱华, 王幼芳, 左勤, 李敏, 魏倩倩, 李晓芹, 赵明水. 浙江西天目山30年来苔藓物种多样性变化. 生物多样性, 2013, 21(2): 170–176. |
| [28] | Lieberman D, Lieberman M, Peralta R, Hartshorn G S. Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. Journal of Ecology, 1996, 84(2): 137–152. DOI:10.2307/2261350 |
| [29] | Auslander M, Nevo E, Inbar M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. Journal of Arid Environments, 2003, 55(3): 405–416. DOI:10.1016/S0140-1963(02)00281-1 |
| [30] | Ferrer-Castán D, Vetaas O R. Pteridophyte richness, climate and topography in the Iberian Peninsula: comparing spatial and nonspatial models of richness patterns. Global Ecology and Biogeography, 2005, 14(2): 155–165. DOI:10.1111/geb.2005.14.issue-2 |
| [31] | Stehn S E, Webster C R, Glime J M, Jenkins M A. Elevational gradients of bryophyte diversity, life forms, and community assemblage in the southern Appalachian Mountains. Canadian Journal of Forest Research, 2010, 40(11): 2164–2174. DOI:10.1139/X10-156 |
| [32] | 陈云, 王海亮, 韩军旺, 韦博良, 贾宏汝, 叶永忠, 袁志良. 小秦岭森林群落数量分类、排序及多样性垂直格局. 生态学报, 2014, 34(8): 2068–2075. |