 2017, Vol. 37
2017, Vol. 37文章信息
- 袁兴伟, 刘尊雷, 程家骅, 田永军
- YUAN Xingwei, LIU Zunlei, CHENG Jiahua, TIAN Yongjun.
- 气候变化对冬季东海外海中下层游泳动物群落结构及重要经济种类的影响
- Impact of climate change on nekton community structure and some commercial species in the Offshore Area of the northern East China Sea in winter
- 生态学报. 2017, 37(8): 2796-2808
- Acta Ecologica Sinica. 2017, 37(8): 2796-2808
- http://dx.doi.org/10.5846/stxb201512222549
-
文章历史
- 收稿日期: 2015-12-22
- 网络出版日期: 2016-10-29





2. 日本水产综合研究中心, 日本海区水产研究所, 新泻 951-8121
2. Japan Sea National Fisheries Research Institute, Fisheries Research Agency (FRA), Suidou-cho, Chuo-ku, Niigata 951-8121, Japan
全球气候变化尤其是气候变暖对海洋生物资源分布和数量变动的影响已日渐显著, 联合国粮农组织每两年一次的世界渔业与养殖报告中多次将全球气候变化对渔业资源的影响作为重要主题之一进行讨论。气候变化通过黑潮和东亚季风的变化影响中国近海和海岸带环境[1-2]。近30年来, 我国沿海海洋表面温度上升了0.9℃, 东海至台湾海峡附近海域升温尤为显著[2-4]。就气候变化对渔业资源影响的研究已引起国内外学者的广泛关注, 海表温度长周期大尺度的冷暖切换不仅对鱼类生理学过程产生了重要影响[5], 而且在群落结构、季节洄游、捕食与被捕食关系等也发挥了重要作用, 导致不同生态属性渔业种类渔获量产生变化。Suh等[6]研究发现1910—2008年东、黄海水域中上层鱼类群落出现的6次优势类群更替均与气候年际变化有关;厄尔尼诺现象导致闽南-台湾浅滩上升流渔场中上层和中下层鱼类群落结构发生改变[7]等。气候变化会导致海洋变暖, 改变了鱼类的栖息环境, 严重影响鱼类的分布和丰度。2000年后在台湾海峡渔业资源调查中发现了13种暖水性鱼类新纪录、在北部湾也有热带暖水性鱼类新记录种出现[8-10], 同时, 水温升高致使黄海主要冷水种数和种群密度下降[11], 日本海西南部和东海北部的暖水种单位捕捞努力量渔获量增加[12]等。另外, 气候变化导致的温度变化还可以影响生态系统中初级生产力和次级生产力, 其也将进一步影响渔业资源量[13-15]。同时, 气候变化还导致了生物多样性的变化等[4, 8, 16-18]。因此, 气候变化对渔业的影响是确凿的, 识别气候变化的影响及掌握渔业对其响应机制对渔业管理部门是有重要价值的。
东海北部外海受黑潮分支-对马暖流控制, 与沿岸流及黄海冷水团相交汇形成良好的渔场, 具有独特的生物地理区系特征和多样性模式。其区系组成以暖水性种类占优势, 暖温性次之, 冷温性很少[19], 该研究水域具有诊断气候变化与渔业资源响应的优良特征。1971—2011年间, 东海捕捞努力量呈持续增长状态, 伴随着捕捞强度的不断增强和环境的持续变化, 东海外海水域鱼类群落的种类组成和多样性状况发生了显著的变化[20-22], 移除捕捞因素所产生的变化趋势后, 其资源状况年间变动可能还与气候变化有关。本研究拟利用中国14家国营渔业公司底拖网渔业生产和东海渔业资源底拖网大面定点调查资料, 利用Fox模型拟合捕捞效应所产生的趋势变化, 移除捕捞效应后, 探讨气候变化对中国东海北部外海中、下层鱼类群落的影响及重要经济种类影响模式, 以期根据气候变化趋势掌握重要经济种类渔获量的变动特征, 为东海外海渔业资源的合理开发与管理提供科学依据。
1 材料与方法 1.1 研究水域本研究选择区域为中国东海北部外海水域, 经纬度范围为29°30′—32°00′N、124°00′—127°30′E, 是黑潮及其西分支——台湾暖流、长江冲淡水、陆架边缘流及黄海冷水团交汇的水域[20, 22-23]。
1.2 数据来源 1.2.1 渔业数据渔业数据由两部分组成, 一部分源自中国14家国营海洋渔业公司机轮拖网渔业捕捞统计资料, 作业范围涵盖了禁渔线以外的东、黄海水域, 时间跨度为1971—1982年, 记录了30余种重要经济鱼类的捕捞日期、渔场位置、捕捞产量和投网网次。1999—2011年渔业数据通过双船底拖网获得的科学调查数据进行补充 (2010年数据缺失), 调查范围为机轮拖网禁渔线至127°00′E, 26°30′—35°00′N, 记录了东海区四季节底拖网渔业种类、拖网站点、拖网时间和捕捞产量。为保持两组数据的一致性和连贯性, 选择两时间段冬季 (第一时间段的当年11、12月及次年1月和2月, 第二时间段为当年的12月) 研究水域中共同记录的17种中、下层重要经济种类进行分析 (表 1)。1971—2011年研究水域渔业捕捞努力量状况以浙江省机动渔船的捕捞努力量 (kW) 为示例, 该省份机动渔船功率年际数据取自《中国渔业统计年鉴》。
| 分组Group | 种类Species | 学名Scientific name | 生态属性Preferred temperature |
| 中层类Midwater species | 黄鲫Hairfin anchovy | Setipinna taty | 暖水性 |
| 银鲳Silver pomfret | Pampus argenteus | 暖水性 | |
| 鳓鱼Chinese herring | Ilisha elongata | 暖水性 | |
| 底层类 (含近底层类) Demersal species (included near-bottom species) | 带鱼Largehead hairtail | Trichiurus japonicus | 暖水性 |
| 小黄鱼Small yellow croaker | Larimichthys polyactis | 暖温性 | |
| 大黄鱼Large yellow croaker | Larimichthys crocea | 暖水性 | |
| 黄姑鱼Yellow drum | Nibea albiflora | 暖水性 | |
| 白姑鱼White croaker | Argyrosomus argentatus | 暖水性 | |
| 鮸Miiuy croaker | Miichthys miiuy | 暖温性 | |
| 绿鳍马面鲀Drab filefish | Navodon septentrionalis | 暖温性 | |
| 短尾大眼鲷Truncatetail bigeye | Priacanthus macracanthus | 暖水性 | |
| 海鳗Pike eels | Muraenesox cinereus | 暖水性 | |
| 大型捕食类Large predatory species | 鲨鱼Shark | ||
| 鳐类Ray | |||
| 无脊椎类invertebrates | 头足类Squid | 暖水性 | |
| 对虾Shrimp | 暖温性 | ||
| 蟹类Crab | 暖水性 |
本文使用的海表温度 (SST) 源自荷兰气候研究中心KNMI Climate Explorer (http://climexp.knmi.nl/select.cgi), 其时间分辨率为月, 空间分辨率为1°×1°。本研究所需SST冬季资料提取范围为122—128°E, 26—34°N, 研究水域冬季海表温度取去年12月及当年1—2月海表温度距平的平均值, 时间跨度为1960—2011年。
引发全球性气候异常的因素繁杂, 表征其变化模态的指标较多, 本研究选择气象数据保存较为完善且能反映大尺度年际变化的太平洋年代际振荡指数 (Pacific Decadal Oscillation index) 作为全球气候变化的重要指标。同时中国近海气候最明显特征是冬、夏季风交替, 季风气候显著, 因此选择亚洲季风指数 (Asian Monsoon index) 作为表征研究水域气候变化的重要指标[24]。两指数数据源自美国国家海洋和大气管理局 (NOAA) 下属的气候预测中心 (http://www.cpc.ncep.noaa.gov/) 提供的月度均值, 为保持与研究时间的同步性, 提取两指数的冬季平均值, 即去年12月及当年1—2月相应指数的平均值, 跨度均为1951—2011年。
1.3 研究方法 1.3.1 渔业数据预处理1971—1982年渔业捕捞统计资料按照经、纬度0.5°×0.5°(1个研究单元) 的空间分辨率进行统计, 并按月份记录了重要经济鱼类的捕捞产量和投网网次。目标种类单位捕捞努力量渔获量 (CPUE) 是以各渔区CPUE为标志变量、相应渔区投网次数为权变量的加权平均值, 单位为箱/网。1999—2011年渔业统计数据以1971—1982年机轮拖网为标准, 按照每网4小时, 每箱20kg的标准, 用效能比进行捕捞努力量标准化。
考虑到海表温度、冬季太平洋年代际振荡指数和亚洲季风指数的时间序列数据单位不一致, 本研究对上述三指数进行标准正态转换处理 (Z-变换), 利用SPSS软件对上述数据进行Kolmogorov-Smirnov正态性检验, 其显著性水平均大于0.05, 表明上述时间序列数据均符合正态分布, 可直接与CPUE的残差数据进行相关性检验和回归分析。
1.3.2 数据分析1971—2011年东海重要经济鱼类冬季渔获量存在年际变化趋势。渔获量的变化同时受到捕捞压力和气候变化的双重影响。根据传统渔业理论, 捕捞压力的单调增长会引起渔获量的变动呈曲线型变化趋势, 这种曲线型变化趋势可以用Fox模型进行拟合。本研究为移除捕捞因素对CPUE的影响而进行了去倾化处理, 即采用Garrod (1969) 对CPUE与捕捞努力量 (f) 间的负指数函数关系进行拟合, 即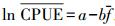
利用Rodionov[27]提出的基于t检验的循序算法STARS (Sequential T-test Analysis of Regime Shift) 检验海表温度和气候指数的跃变指数RSI (regime shift index)。设定冬季海表水温、太平洋年代际振荡指数和亚洲季风指数的显著性检验置信水平P=0.1, 跃变的切断长度L=20, Huber权重参数H=2。
1.3.3 Shannon多样性指数物种多样性对于维护生态系统稳定和生物资源持续利用具有重要作用。本研究选择Shannon多样性指数 (H′) 作为反映丰富度和均匀度的综合指标, 其表达式[28]为
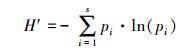
|
式中, pi为种类i的生物量占总生物量的比例, s为生物种类数, ln为以e为底的自然对数。
1.3.4 群落结构与海表温度相关分析本研究采用线性分析法研究游泳动物群落结构中目标种类的CPUE残差与海表温度距平的相关关系, 探讨中、下层游泳动物群落的波动与周期变化规律及其对气候变化的响应。数据采用SPSS 18.0软件进行相关分析, 显著性水平取0.05。
2 结果 2.1 研究水域海表温度及气候变化特征1960—2011年间, 研究水域冬季海表温度发生了显著的年际变化, 大体呈先降温再逐步升温过程, 具体表现为海表温度由1960—1982年的 (18.14±0.35)℃上升到1983—2011年的 (19.40±0.66)℃(均值±标准差), 同期海表温度距平亦由-0.87℃上升到0.64℃, 分别对应“冷相位”周期和“暖相位”周期 (图 1)。从趋势变化上看, 研究水域发生了两次显著的跃变, 具体可分三个阶段, 1960—1982年为降温期, 尤其在1973/1974年和1975/1976年发生的两次较强的拉尼娜事件, 促使海表温度加速下降, 使得海表温度在1976年达到了最低的17.57℃, 之后在低位振荡, 本研究定义该阶段为寒冷期;伴随着1982—1983发生的一次较强的厄尔尼诺现象, 海表温度迅速上升, 并发生了一次跃变, 之后的1987—1988年又发生一次强厄尼尔诺现象, 海表温度进一步攀升, 直至1997—1998年发生的有记录以来最强的一次厄尔尼诺现象, 水温达到了最高的20.75℃, 致使研究水域发生了再一次的跃变, 该周期内海表温度为 (18.98±0.57)℃, 定义该期间为增温期;紧随强厄尔尼诺现象的发生, 1998—1999年和1999—2000年全球陆续发生了两次中等尺度和较大尺度的拉尼娜事件, 致使海表温度在高位波动并呈小幅下降趋势, 但海表温度平均值仍显著高于寒冷期, 本研究定义1999—2011年为温暖期。

|
| 图 1 1971—2011年研究水域冬季海表温度距平及跃变指数 Fig. 1 Anomalies values and regime shift index estimated by STARS for the sea surface temperature in winter from 1971—2011 |
1951—2011年期间, 冬季太平洋年代际振荡指数发生了两次跃变:一次在1977年, 标准正态转换后的太平洋年代际振荡指数距平由负值转换为正值, 表明太平洋水域表层海水温度由冷变暖, 即由“冷相位”周期转变至“暖相位”周期;另一次在2008年, 结果与前一周期恰相反。同时, 冬季亚洲季风指数亦发生了两次跃变:一次在1988年, 另一次在2006年, 转换后的冬季亚洲季风指数距平由正值转换为负值, 再由负值转换为正值, 表明低纬度太平洋水域表层海水温度经历由冷到暖再到冷的过程 (图 2)。

|
| 图 2 1951—2011年研究水域冬季太平洋年代际振荡指数和冬季亚洲季风指数距平、距平累计和及跃变指数 Fig. 2 Anomalies values, their cumulative sums and regime shift index estimatd by STARS for winter Pacific Decadal Oscillation index and winter Asian Monsoon index in the study area from 1951—2011 |
1971—2011年间, 研究水域中、下层游泳动物群落结构发生了不同程度的年际变化。首先, 从游泳动物群落结构组成角度看 (图 3), 寒冷期和温暖期游泳动物所占比例发生显著变化, 具体表现为:寒冷期间, 底层类为群落结构的绝对优势地位, 平均占比为95.29%;中层鱼类是次优势种类, 平均占比为2.65%;无脊椎类平均占比1.48%, 其余为大型捕食类。温暖期间, 底层鱼类仍旧是首要优势种类, 但伴随着大黄鱼资源的严重衰退以及带鱼、小黄鱼资源CPUE的下降, 其占比有所下降, 降至78.96%, 降幅达16.08%;而中层鱼类占比较寒冷期显著增加3.27倍, 占比达14.61%;无脊椎类占比亦有大幅增加, 增至6.41%;大型捕食类占比急剧下降, 不足0.01%。其次, 从游泳动物适温性角度看, 两阶段游泳动物的占比亦有所变化。就海面气候而言, 研究水域属于亚热带气候, 主要以暖水种和暖温种为主, 本研究并没有发现冷温性和冷水性种类。伴随1982—1997年的两次大幅升温过程, 以带鱼、中层鱼类及无脊椎类为代表的暖水性种类占比有所增加 (表 1), 平均占比由寒冷期的62.44%增至72.11%;相应地, 以小黄鱼为代表的暖温性种类占比有一定程度的降低, 平均占比由37.56%降至温暖期的27.89%(图 3)。

|
| 图 3 1971—2011年研究水域游泳动物不同类群及适温性种类CPUE占比的年际变化图 Fig. 3 Annual changes in the proportion of different species group about CPUE in the study area during 1971—2011 |
(1) 游泳动物不同类群资源变化特征
对研究水域中的不同类群游泳动物的CPUE利用Fox模型分别进行回归分析, 结果显示各类群CPUE回归分析后提取的残差及残差和均在1976—1977年出现了极值, 说明该期间游泳动物的资源状况发生了年际变化 (图 4)。具体地, 寒冷期间, 中层鱼类、无脊椎类的资源指数在1976—1977年发生由降低到缓慢增高的转变, 而底层类的资源指数由1971年开始逐渐增高, 并在1976年达到最高值, 接着振荡走低;同时, 大型捕食类在1970年代前期的资源变化趋势与底层类相反, 呈振荡下降趋势, 在1977年突然增至历史最高点, 然后振荡走低;温暖期间, 中层鱼类在1999年达到最高值, 随后呈逐步走低的趋势, 资源指数平均值高于寒冷期, 底层鱼类逐步走低并在2003年达到最低值, 随后呈逐步走高趋势, 但资源指数的平均值低于寒冷期, 无脊椎类呈波动增加趋势, 资源指数平均值高于寒冷期, 而随着大型捕食类资源的逐渐衰退, 其资源指数微幅波动, 其平均值显著低于寒冷期。对不同类群游泳动物CPUE残差与海表温度距平距平进行相关分析后显示 (表 2), 中层类与海表温度相关系数最高, 且Pearson相关系数的双侧显著性检验结果显示, 两者相关性是显著的;其次是无脊椎类和底层类, 而大型捕食类却与海表温度呈负相关关系。

|
| 图 4 1971—2011年研究水域游泳动物不同类群CPUE残差及残差和的年际变化图 Fig. 4 Annual changes in the residuals of CPUE and their cumulative sums of different species group in the study area during 1971—2011 |
| 中层类Midwater species | 底层类Demersal species | 无脊椎类Invertebrates | 大型捕食类Large predatory species | 暖水种Warm water species | 暖温种Warm temperature species | 黄鲫Setipinna taty | 银鲳Pampus argenteus | 小黄鱼Larimichthys polyactis | 带鱼Trichiurus japonicus | 头足类Squid | 对虾Shrimp | |
| SST | 0.44* | 0.20 | 0.33 | -0.29 | 0.41 | -0.18 | 0.61* | 0.42* | -0.30 | 0.27 | 0.38 | -0.34 |
| *表示P < 0.05下相关显著 | ||||||||||||
从游泳动物适温性角度看, 1971—1977年间, 伴随着研究水域海表水温的振荡走低, 以带鱼和中层鱼类为代表的暖水种资源指数在呈先增后降的变化趋势, 并在1977年达到最小值;随着海表水温振荡走高, 暖水种的资源指数呈逐步增加趋势, 并在1999年达到最大值, 随后宽幅振荡呈逐步下降趋势, 总体上, 温暖期间暖水种的资源指数平均值高于寒冷期 (图 5)。而以小黄鱼为代表的暖温种的变化趋势与暖水种大体相反, 寒冷期前期, 资源指数振荡走高, 中后期, 伴随海表水温的逐步升高, 资源指数逐步走低;温暖期间, 资源指数振荡走低, 在2004年后达到最低值, 随后振荡走高, 总体上, 温暖期间暖温种的资源指数平均值低于寒冷期。对不同生态属性游泳动物CPUE残差与海表温度进行相关分析后显示 (表 2), 暖水种和暖温种分别与海表温度呈正、负相关关系, 且前者的相关系数大于后者的绝对值。

|
| 图 5 1971—2011年研究水域暖水种和暖温种CPUE残差及残差和的年际变化图 Fig. 5 Annual changes in the residuals of CPUE and their cumulative sums of warm water species and warm temperature species in the study area during 1971—2011 |
(2) 几种重要经济种类资源变化特征
图 6列举了5种不同栖息水层和生态属性的经济种类资源指数变化曲线。总体上, 中层类和底层类资源变化大致呈现相反的趋势, 即中层类在寒冷期的资源指数低于温暖期, 而底层类在寒冷期的资源指数高于温暖期。其中1977年是一个重要时间节点, 不同种类均在该节点前后发生了显著变化。具体地, 寒冷期:以黄鲫、银鲳、带鱼及头足类 (以太平洋褶柔鱼 (Todarodes pacificus) 和剑尖枪乌贼 (Uroteuthis edulis) 为主) 为代表的暖水种随着海表温度的进一步降低, 其资源指数大体呈现下降趋势, 并在1977年达到最低值, 伴随海表温度的升高, 其资源指数逐步回升;以小黄鱼和对虾为主的底层类暖温种, 其资源指数伴随海表温度的降低而振荡走高, 大体在1977年前后达到最高值, 随后逐步下降。温暖期内, 由于海表温度在1998年达到最高值后呈振荡走低趋势, 暖水性种类的资源指数在1999年达到最高值, 随后与海表水温保持大致同步态势, 即振荡下降, 但仍旧在相对较高范围内;暖温种的资源指数变动趋势大体呈振荡下降趋势, 资源指数平均值低于寒冷期。对不同经济种类的CPUE残差与海表温度进行相关分析后显示 (表 2), 黄鲫和银鲳与海表温度呈显著性正相关关系, 带鱼和头足类与海表温度亦呈正相关关系, 而小黄鱼和对虾则与海表温度负相关。

|
| 图 6 1971—2011年研究水域重要经济种类的CPUE残差及残差和的年际变化图 Fig. 6 Annual changes in the residuals of CPUE and their cumulative sums of different species during 1971—2011 |
总体上, Shannon多样性指数与海表温度的变化特征保持较高的同步性, 寒冷期游泳动物的多样性指数低于温暖期 (图 7)。具体地, 多样性指数在寒冷期间宽幅波动并在1976年达到最低点, 1978年达到次低点, 随后振荡走高, 但仍处在较低水平;2000年多样性指数跃升至最大值后开始呈振荡走低趋势, 但仍处在高位。对比海表温度和太平洋年代际振荡指数的变化特征, 初步断定研究水域游泳动物多样性指数的变化可能与气候变化有关。分别将寒冷期和温暖期中各年份冬季游泳动物Shannon多样性指数与当年冬季的海表温度进行相关性检验分析后发现, 研究水域游泳动物多样性指数均与海表温度呈正相关关系 (R寒=0.41, P寒=0.24; R温=0.47, P温=0.18)。

|
| 图 7 1971—2011年Shannon指数的年际变化图 Fig. 7 Annual changes in Shannon diversity index during 1971—2011 |
大量研究已证实, 在中纬度北太平洋海域发生了大气-海洋环境的年代际振荡或“体系转变”, 尤以1976/1977年的太平洋年代际振荡指标为代表[29-31]。所谓体系转换, 即意味着突然的而非渐进的变化, 在1年左右的时间内从一种特征行为转变成另一种特征行为。PDO是一种类似ENSO变化模式的长周期气候波动现象, 在PDO转换阶段, 海洋中涌升流和平流发生变化, 同时由于海洋环流具有大范围输送浮游动物的能力, 因而可使食物链中较低营养级的生物得到大量营养物质, 为鱼类提供丰富的饵料[32-33]。考虑到时间序列 (1971—2010年) 较短, 且传统的Mann-Kendall (M-K) 检验和滑动t检验均无法有效检测时间序列后期的突变点, 本研究利用Rodionov提出的一种基于t检验的循序算法STARS进行检验分析。该方法较前两种检验更为严谨, 能够检测到位于时间序列后期的跃变, 但也有一定弊端, 即它无法完全去除边界效应的影响, 因而无法保证跃变时间点的绝对准确性。图 2表明, 利用该方法检验的结果与King[30]及Mantua等[32]保持一致;同时STARS检测出东亚季风发生大尺度跃变的时间节点亦与Tian等[12]吻合。该方法已被证明是一种便捷、有效的检验手段, 并得到了广泛的应用[12, 27, 34]。STARS检验结果显示 (图 1), 研究水域在经历两次明显的升温过程, 且两次发生跃变的时间节点恰与ENSO事件的出现相一致, 而ENSO事件是致使海表水温已发生显著变化的重要诱因。考虑到我国位于东亚气候区, STARS检测出东亚季风指数在1988年发生了由强到弱的显著变化, 说明海表温度在此时间节点发生了由冷到暖的转变。综上所述, 可以确认研究水域在1982—1997年间发生了显著的气候变化。
3.2 气候变化对鱼类群落结构的影响东海位于东亚气候区, 海洋物理环境较为复杂。气候变化引起海表温度的波动进而影响海洋鱼类数量结构变动[35]。水温的变化会直接影响鱼类的生长、摄食、产卵及洄游等, 影响鱼类种群的变化, 并最终影响到渔业资源的数量、质量及其开发利用[36]。气候变化导致了研究水域海表温度急剧升高, 致使研究水域中不同栖息水层游泳动物CPUE的变化模式以及其分别与海表温度的相关性分析结果和东海周边临近水域鱼类群落结构变化大体一致[6-7, 12]。如对马海峡水域在20世纪70年代末出现了结构性转变, 引起鱼类生物量、渔获量和生态系统的结构变化以及优势种的转变[37], 这种海洋鱼类资源结构的长周期变化与北极涛动 (Arctic Oscillation, 简写AO) 和太平洋年代际振荡 (Pacific Decadal Oscillation, 简写PDO) 有关[38];气候变化促使鱼类向更高纬度水域迁移, 造成不同生态属性种类资源状况发生改变。Cheung等[39]指出, 在一个中等的气候变暖事件中, 全球的鱼类分布可能向两极移动45—49km/年代。利用Fish Bio-energetic Model 2分析显示, 水温升高造成小黄鱼、带鱼和大黄鱼等底层鱼类性成熟年龄提前, 怀卵量减少, 幼鱼成活率降低, 进而导致成鱼年龄缩短最终导致其成鱼数量减少, 渔获量下降, 其中对大黄鱼的影响尤为显著[40]。气候变化改变洋流变化, 通过控制海洋生态系统的营养盐供给来影响海洋初级生产力, 初级生产力的变动最终影响鱼类的数量变动。随着人类活动的增加, 输入到东海的营养盐有显著增加趋势[41], 而本研究中的中层鱼类和无脊椎类的摄食对象主要是浮游动物[20], 气候变化提升初级生产力和人类活动引起的营养盐增多趋势都有利于中层类的资源量的增加, 从而促使其资源状况显著好转。张家坤等[42]指出, 长江口以北 (32°—36°N) 水域中上层鱼类渔获量和平均网产均与海水温度呈显著负相关关系, 说明温度对研究区域鱼类资源有重要影响。1997年FAO研究报告认为, 鳀鱼、太平洋沙丁鱼 (Sardinops sagsx) 等中上层鱼类资源的变动是由长期的气候变化所决定的[43]。另外, 寒冷期和温暖期的暖水种和冷水种资源状况大体呈现相反趋势, 且相关性分析显示暖水种与海表温度距平呈正相关关系 (表 2), 这主要是因为暖水种以中层类和无脊椎类为主, 且中层类占比超过60%, 加之中层类与水温的显著正相关关系不难得出两者的正相关性。考虑到小黄鱼在底层类占比较高且它和对虾均与海表温度的负相关性, 易知底层类与海表温度呈负相关关系。该相关性之所以较弱, 一方面由带鱼在底层类的占比仅次于小黄鱼且与温度呈正相关造成的, 另一方面, 种类适应性变化和群落结构演替也是主因。Tian等[44]利用主成分析 (PCA) 对对马海峡水域底层不同生态属性种类资源变动进行研究后表明, 上世纪80年代末的气候变化造成对马海峡水域海表温度的急剧升高, 致使暖水性鱼类CPUE由低至高, 而冷水性鱼类CPUE由高至低。
鱼类生物量变化受到捕捞死亡率、栖息地环境丢失、环境污染、生态环境扰动以及物种间的相互影响共同制约, 过度捕捞和气候变化共同影响了鱼类补充量变化和优势种的转变, 使得海洋生态系统变得越来越不稳定[45]。大型捕食类中鳐类资源的衰退更多受到底拖网的破坏性捕捞所致, 而鲨类则更多是受饵料鱼类的过度捕捞影响, 致使1999年以来资源一直处于严重衰减状态, 其资源指数仅做微幅波动, 资源指数显著低于寒冷期, 气候变化或许只是其资源变动的一个因素。海表温度的升高对不同经济种类的影响模式不尽相同, 大多数鱼类的补充量水平和鱼卵孵化与当年的温度密切相关[46-47], 如黄鲫[48-49]、头足类[50-51]的繁殖、补充和洄游在很大程度上受到海表温度的影响, 本研究发现其资源指数与海表温度距平呈显著正相关关系 (表 2)。带鱼有昼夜移动的特性, 其资源变动很大程度上受到水温变化的影响, 表 2显示, 带鱼CPUE残差与海表温度距平呈正相关关系。陈永利等[52]研究发现, 东海带鱼渔获量与黑潮陆坡区的海表水温具有显著的正相关;王跃中等[15]研究显示, 南海北部海表温度与带鱼渔获量变动显著正偏相关, 说明水温变化显著影响到带鱼的资源变动。小黄鱼资源指数与海表水温距平呈弱负相关关系, 温暖期的资源指数低于寒冷期。李忠炉[53]研究发现, 黄海34.5°N以北海区小黄鱼相对资源密度的升高和气候变化引起的小黄鱼分布变化有关。因为SST的升高可促使小黄鱼种群向深水或高纬度海区移动, 致使小黄鱼群体向北移动, 34°N以南水域资源密度有所下降。刘允芬[40]利用模型预测后发现, 气候变化导致东海东北部小黄鱼产量减少8%左右。另外, 研究发现, 很多经济种类CPUE残差变化的时间节点与温度跃变的节点不一致, Tian等[12]认为年龄组成的差异是造成鱼类资源状况跃变和气候变化不同步的主要原因。由此推断小黄鱼年龄结构特征的改变 (由3龄及以上降至当龄或1龄)[54]可能是导致其资源指数与温度距平相关系数较小的重要因素。
气候变化引起的海表温度、CO2浓度和海平面的上升、降水量变化和海洋水文结构变化以及紫外线辐射增强等对海洋生物多样性会产生较大影响[55]。研究水域既受到黑潮暖流、台湾暖流等影响, 同时又受到黄海冷水团的影响, 以亚热带成分占优势, 属于印度-西太平洋暖水区系, 生物多样性较高[20]。温暖期的生物多样性指数略高于寒冷期, 一方面, 可能是由于大尺度生物地理模式和气候变化的交互作用导致了中高纬度鱼类群落丰富度增加[39]。Niquen[56]研究发现, ENSO事件促使智利近岸水域温度骤升, 致使海洋生物多样性指数由0.87升至1997/1998的1.23—1.70。东海周边的韩国、日本水域在20世纪80年代左右处于低温期, 海洋鱼类平均营养级和多样性指数较低, 伴随80年代末温度的急剧上升, 该水域生态结构和优势种发生显著变化, 多样性显著增加, 这种显著性变化是由气候变化有关[6, 12, 38, 57]。另一方面, 由于海表温度的上升导致海洋生物物种分布向北迁移, 暖水性种类增多, 导致多样性指数增加。2000年后台湾海峡发现了13中暖水性鱼类新记录种, 北部湾也有热带暖水性记录种苏门答腊金线鱼 (Nemipterus mesoprion), 研究表明气候变化是导致这些新记录种生存空间北移的主要原因[8-10]。
3.3 研究展望东海地处东亚强季风区并东濒西北太平洋, 海洋生态系统受东亚季风、西太平洋强边界流黑潮、沿岸流和长江冲淡水等气候因子的影响显著, 必然会受到气候变化的影响[4]。同时, 影响渔业资源变动的因素有很多, 除捕捞因素和水温变化外, 还可能受到降水、季风、海洋环流和热带气旋等因子变化的影响[14]。政府间气候变化专业委员会 (IPCC) 认为, 在未来50—100年间, 气候变化对世界渔业的影响可能超过过度捕捞。中国针对气候变化对海洋生物影响的研究尚处于初级阶段, 对气候变化事实的认知和对其自然规律的认识等问题亟待加深, 针对气候变化对海洋生态系统结构和功能影响的研究仍需加强, 渔业资源对气候变化的响应机制和机理仍未查明, 模拟或预测气候变化对海洋生物群落结构影响的研究有待深化, 同时也缺乏长时间序列的海洋生物调查资料等。游泳动物群落结构与气候变化的复杂关系印证了在气候变化下渔业管理的必要性和针对性。在健全和完善捕捞渔业准入制度、实施投入控制管理与产出控制管理并举、以及执行产卵场保护和伏季休渔政策的基础上, 结合不同生态属性种类对气候变化的响应模式, 预测和分析优势种波动变化规律, 合理评估捕捞压力, 以便更好地提出适应性和预防性渔业管理策略[21]。全球气候变化中许多问题的暴露有一定的滞后性, 人类应采取各种预防措施对渔业资源进行养护和管理, 尽可能减少对自然生态的人为破坏, 实现渔业资源的可持续利用。
| [1] | 《第二次气候变化国家评估报告》编写委员会. 第二次气候变化国家评估报告. 北京: 科学出版社, 2011. |
| [2] | 杜建国, CheungW W L, 陈彬, 周秋麟, 杨圣云, YeG Q. 气候变化与海洋生物多样性关系研究进展. 生物多样性, 2012, 20(6): 745–754. |
| [3] | Belkin I M. Rapid warming of large marine ecosystems. Progress in Oceanography, 2009, 81(1/4): 207–213. |
| [4] | 蔡榕硕. 气候变化对中国近海生态系统的影响. 北京: 海洋出版社, 2010. |
| [5] | Reichardt M R. The Effects of Climate Variability on Early Marine Growth And Age at Maturity of Fraser River Sockeye Salmon (Oncorhynchus Nerka)[D]. Colwood: Royal Roads University, 2005. |
| [6] | Gong Y, Suh Y S. Climate change and fluctuations of pelagic fish populations in the Far East region. Journal of Ecology and Field Biology, 2012, 35(1): 15–25. |
| [7] | 何发祥, 洪华生, 陈刚. ENSO现象与台湾海峡西部海区中下层鱼类渔获量关系. 海洋湖沼通报, 2003(1): 27–34. |
| [8] | 陈宝红, 周秋麟, 杨圣云. 气候变化对海洋生物多样性的影响. 台湾海峡, 2009, 28(3): 437–444. |
| [9] | 戴天元. 福建海区渔业资源生态容量和海洋捕捞业管理研究. 北京: 科学出版社, 2004: 35–210. |
| [10] | 黄梓荣, 王跃中. 北部湾出现苏门答腊金线鱼及其形态特征. 台湾海峡, 2009, 28(4): 516–519. |
| [11] | 刘瑞玉. 中国海物种多样性研究进展. 生物多样性, 2011, 19(6): 614–626. |
| [12] | Tian Y J, Kidokoro H, Watanabe T, Iguchi N. The late 1980s regime shift in the ecosystem of Tsushima Warm Current in the Japan/East Sea: evidence from historical data and possible mechanisms. Progress in Oceanography, 2008, 77(2/3): 127–145. |
| [13] | Kennedy V S. Anticipated effects of climate change on estuarine and coastal fisheries. Fisheries, 1990, 15(6): 16–24. DOI:10.1577/1548-8446(1990)015<0016:AEOCCO>2.0.CO;2 |
| [14] | Qiu Y S, Wang Y Z, Chen Z Z. Runoff-and monsoon-driven variability of fish production in East China Seas. Estuarine, Coastal and Shelf Science, 2008, 77(1): 23–34. DOI:10.1016/j.ecss.2007.09.001 |
| [15] | 王跃中, 孙典荣, 陈作志, 贾晓平. 气候环境因子和捕捞压力对南海北部带鱼渔获量变动的影响. 生态学报, 2012, 32(24): 7948–7957. |
| [16] | Barange M, Field J G, Harris R P, Hofmann E E, Perry R I, Werner F E. Marine Ecosystems and Global Change. New York: Oxford University Press, 2011. |
| [17] | Bell J D, Johnson J E, Hobday A J. Vulnerability of tropical Pacific fisheries and aquaculture to climate change. Noumea, New Cledonia: Secretariat of the Pacific Community, 2011. |
| [18] | Hollowed A B, Barange M, Ito S I, Kim S, Loeng H. Climate change effects on fish and fisheries: forecasting impacts, assessing ecosystem responses, and evaluating management strategies. Sendai, Japan: ICES, 2010. |
| [19] | 郑元甲, 陈雪忠, 程家骅, 王云龙, 沈新强, 陈卫忠, 李长松. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 2003: 1–424. |
| [20] | 刘尊雷, 袁兴伟, 杨林林, 严利平, 田永军, 程家骅. 气候变化对东海北部外海越冬场渔业群落格局的影响. 应用生态学报, 2015, 26(3): 901–911. |
| [21] | Lin C, Xu B, Huang S. Long-term variations in the oceanic environment of the East China Sea and their influence on fisheries resources//Beamish R J, ed. Climate change and northern fish populations. Canadian Special Publication of Fisheries and Aquatic Sciences, 1995, 121: 307-315. |
| [22] | 袁兴伟, 刘尊雷, 姜亚洲, 林楠, 张辉, 程家骅. 冬季东海外海鱼类群落特征及其对拉尼娜事件的响应. 中国水产科学, 2014, 21(5): 1039–1047. |
| [23] | Hu A Y, Jiao N Z, Zhang R, Yang Z. Niche partitioning of marine Group I Crenarchaeota in the euphotic and upper mesopelagic zones of the East China Sea. Applied and Environmental Microbiology, 2011, 77(21): 7469–7478. DOI:10.1128/AEM.00294-11 |
| [24] | Gong D Y, Wang S W, Zhu J H. East Asian winter monsoon and Arctic Oscillation. Geophysical Research Letters, 2001, 28(10): 2073–2076. DOI:10.1029/2000GL012311 |
| [25] | 李寇军, 邱永松, 王跃中. 自然环境变动对北部湾渔业资源的影响. 南方水产, 2007, 3(1): 7–13. |
| [26] | 王继隆, 李继龙, 杨文波, 王国伟. 主要气侯因子对东中国海主要经济鱼种生物量的影响研究. 湖南农业科学, 2010(9): 142–147. |
| [27] | Rodionov S N. A sequential algorithm for testing climate regime shifts. Geophysical Research Letters, 2004, 31(9): L09204. |
| [28] | Wilhm J L. Use of biomass units in Shannon's formula. Ecology, 1968, 49(1): 153–156. DOI:10.2307/1933573 |
| [29] | Mantua N J, Hare S R. The Pacific decadal oscillation. Journal of Oceanography, 2002, 58(1): 35–44. DOI:10.1023/A:1015820616384 |
| [30] | King J R. Report of the study group on fisheries and ecosystem responses to recent regime shifts. PICES Scientific Report No. 28, 2005: 162–162. |
| [31] | 商少凌, 张彩云, 洪华生. 气候-海洋变动的生态响应研究进展. 海洋学研究, 2005, 23(3): 14–22. |
| [32] | Mantua N J, Hare S R, Zhang Y, Wallace J M, Francis R C. A Pacific interdecadal climate oscillation with impacts on salmon production. Bulletin of the American Meteorological Society, 1997, 78(6): 1069–1079. DOI:10.1175/1520-0477(1997)078<1069:APICOW>2.0.CO;2 |
| [33] | 樊伟, 程炎宏, 沈新强. 全球环境变化与人类活动对渔业资源的影响. 中国水产科学, 2001, 8(4): 91–94. |
| [34] | 梁苏洁, 丁一汇, 赵南, 孙颖. 近50年中国大陆冬季气温和区域环流的年代际变化研究. 大气科学, 2014, 38(5): 974–992. |
| [35] | Masuda R. Seasonal and interannual variation of subtidal fish assemblages in Wakasa Bay with reference to the warming trend in the Sea of Japan. Environmental Biology of Fishes, 2008, 82(4): 387–399. DOI:10.1007/s10641-007-9300-z |
| [36] | 陈新军. 渔业资源与渔场学. 北京: 海洋出版社, 2004: 116–122. |
| [37] | Zhang C I, Lee J B, Kim S, Oh J H. Climatic regime shifts and their impacts on marine ecosystem and fisheries resources in Korean waters. Progress in Oceanography, 2000, 47(2/4): 171–190. |
| [38] | Tian Y J, Kidokoro H, Watanabe T. Long-term changes in the fish community structure from the Tsushima warm current region of the Japan/East Sea with an emphasis on the impacts of fishing and climate regime shift over the last four decades. Progress in Oceanography, 2006, 68(2/4): 217–237. |
| [39] | Cheung W W L, Lam V W Y, Sarmiento J L, Kearney K, Watson R, Pauly D. Projecting global marine biodiversity impacts under climate change scenarios. Fish and Fisheries, 2009, 10(3): 235–251. DOI:10.1111/faf.2009.10.issue-3 |
| [40] | 刘允芬. 气候变化对我国沿海渔业生产影响的评价. 中国农业气象, 2000, 21(4): 1–5. |
| [41] | Ning X, Lin C, Hao Q, Liu C, Le F, Shi J. Long term changes in the ecosystem in the northern South China Sea during 1976-2004. Biogeosciences Discussions, 2008, 5(5): 3737–3779. DOI:10.5194/bgd-5-3737-2008 |
| [42] | 张甲珅, 苏奋振, 杜云艳. 东海区中上层鱼类资源与海表温度关系. 资源科学, 2004, 26(5): 147–152. |
| [43] | Lluch-Cota D B, Hernández-Vázquez S, Lluch-Cota S E. Empirical investigation on the relationship between climate and small pelagic global regimes and El Nino-Southern Oscillation (ENSO). FAO Fisheries Circular. NO: 934, Rome: FAO, 1997. |
| [44] | Tian Y J, Kidokoro H, Fujino T. Interannual-decadal variability of demersal fish assemblages in the Tsushima Warm Current region of the Japan Sea: impacts of climate regime shifts and trawl fisheries with implications for ecosystem-based management. Fisheries Research, 2011, 112(3): 140–153. DOI:10.1016/j.fishres.2011.01.034 |
| [45] | Brander K. Impacts of climate change on fisheries. Journal of Marine Systems, 2010, 79(3/4): 389–402. |
| [46] | Drinkwater K F, Myers R A. Testing predictions of marine fish and shellfish landings from environmental variables. Canadian Journal of Fisheries and Aquatic Sciences, 1987, 44(9): 1568–1573. DOI:10.1139/f87-189 |
| [47] | Kim S, Jung S, Zhang C I. The effect of seasonal anomalies of seawater temperature and salinity on the fluctuation in yields of small yellow croaker, Pseudosciaena Polyactis, in the Yellow Sea. Fisheries Oceanography, 1997, 6(1): 1–9. DOI:10.1046/j.1365-2419.1997.00025.x |
| [48] | 刘勇, 程家骅, 陈学刚. 东海北部与黄海南部黄鲫群体洄游分布的初步研究. 海洋水产研究, 2006, 27(3): 1–6. |
| [49] | 李增光, 叶振江, 张弛, 庄龙传, 王明琳. 黄海南部春季帆张网黄鲫CPUE分布及其影响因素分析. 中国海洋大学学报, 2013, 43(5): 30–36. |
| [50] | Kang Y S, Kim J Y, Kim H G, Park J H. Long-term changes in zooplankton and its relationship with squid, Todarodes pacificus, catch in Japan/East Sea. Fisheries Oceanography, 2002, 11(6): 337–346. DOI:10.1046/j.1365-2419.2002.00211.x |
| [51] | Kidokoro H, Goto T, Nagasawa T, Nishida H, Akamine T, Sakurai Y. Impact of a climate regime shift on the migration of Japanese common squid (Todarodes pacificus) in the Sea of Japan. ICES Journal of Marine Science, 2010, 67(7): 1314–1322. |
| [52] | 陈永利, 王凡, 白学志, 白虹, 纪风颖. 东海带鱼 (Trichiurus haumela) 渔获量与邻近海域水文环境变化的关系. 海洋与湖沼, 2004, 35(5): 404–412. |
| [53] | 李忠炉. 黄渤海小黄鱼、大头鳕和黄鮟鱇种群生物学特征的年际变化[D]. 北京: 中国科学院研究生院, 2011. |
| [54] | 林龙山, 凌建忠, 程家骅, 俞连福. 小黄鱼资源状况及合理利用意见. 海洋渔业, 2000, 22(3): 120–122. |
| [55] | Intergovernmental Panel on Climate Change (IPCC). Climate Change 2007: Synthesis Report. Contribution of Working Groups I, Ⅱ and Ⅲ to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland: IPCC, 2007. |
| [56] | Ñiquen M, Bouchon M. Impact of El Niño events on pelagic fisheries in Peruvian waters. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2004, 51(6/9): 563-574. |
| [57] | Zhang C I, Lee J B, Seo Y I, Yoon S C, Kim S. Variations in the abundance of fisheries resources and ecosystem structure in the Japan/East Sea. Progress in Oceanography, 2004, 61(2/4): 245–265. |