 2017, Vol. 37
2017, Vol. 37文章信息
- 牛莉平, 许嘉巍, 靳英华, 尹海霞, 王嫒林, 刘羽霞, 张英洁
- NIU Liping, XU Jiawei, JIN Yinghua, YIN Haixia, WANG Ailin, LIU Yuxia, ZHANG Yingjie.
- 温带森林不同演替阶段下的土壤CO2排放通量昼间变化
- Diurnal variation in soil CO2 emission flux at different stages of temperate forest succession
- 生态学报. 2017, 37(8): 2673-2683
- Acta Ecologica Sinica. 2017, 37(8): 2673-2683
- http://dx.doi.org/10.5846/stxb201512182529
-
文章历史
- 收稿日期: 2015-12-18
- 网络出版日期: 2016-10-29

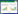




大气温室气体的变化是直接或间接引起气候和环境变化的原因[1], CO2作为主要的温室气体, 其源、汇和排放量的研究成为了焦点。土壤是全球陆地系统中最大的碳库载体, 土壤碳库碳贮存量占全球碳总储存量的67%, 其贮存量约为1300—2 000 PgC[2], 大约是陆地植被碳库的2.5—3倍, 大气碳库的2倍[3], 土壤有机碳库在全球碳循环中具有重要作用[4], 土壤碳库中碳储存量的动态变化对全球气候变化至关重要。
土壤排放CO2的过程被称为土壤呼吸[5], 包括根际呼吸和异养呼吸, 即除了根本身和根际微生物的呼吸, 还有土壤微生物和土壤动物的呼吸[6]。全球每年因土壤呼吸作用向大气中释放的碳的估算值约为75—120 Pg[7], 其量值仅次于全球陆地总初级生产力 (GPP) 的估算值[8]。土壤呼吸是陆地生态系统碳循环的重要环节[9], 也是土壤向大气释放CO2的主要途径[10]。土壤呼吸已成为全球变化和陆地生态系统碳循环的重要研究内容之一[11]。
森林是陆地生态系统的主体, 研究森林土壤碳库的排放规律, 对全球碳平衡有着重要意义。温带湿润区的地带性植被为红松阔叶林。但近年来, 在人为强干扰下, 在长白山地区除自然保护区外, 红松阔叶林地基本消失, 代之以处于演替不同阶段的灌木林 (或幼龄林)、白桦林 (或杨木林、蒙古栎林)、阔叶杂木林, 干扰更严重地区, 已退化为草地。不同演替阶段植被群落组成、结构及群落内小气候差别较大, 向土壤补充的有机碳数量也显著不同, 土壤中根际呼吸和异养呼吸差别明显, 土壤CO2排放通量存在较大差异。鉴于对红松阔叶林下的土壤排放通量的研究较多, 结论明确, 本研究重点分析草地、灌木林地、白桦林地、阔叶杂木林地四种测量样地与红松阔叶林地的昼间土壤CO2排放通量特点的异同, 实现草地、灌木林地、白桦林地、阔叶杂木林地土壤呼吸过程中CO2排放量的估算。并采用时空替代法, 选取了红松针阔叶混交林演替序列的5个不同阶段, 来完成温带森林不同演替阶段下的土壤CO2排放通量的规律分析, 为科学的评估我国温带森林演替过程中固碳潜力的变化提供一定的理论依据。
1 研究地区与方法 1.1 研究区自然概况研究区位于长白山自然保护区北部的实验区 (图 1)。长白山自然保护区位于吉林省东南部, 地理位置为41°41′—42°51′ N, 127°42′—128°16′ E。该地区属于典型的温带季风气候区, 春季干燥而多风;夏季短暂, 温暖而多雨;秋季凉爽而多雾;冬季漫长, 晴朗而寒冷。年均气温0.3—3. 9℃, 年均降水量632.8—782.4 mm[12-13], 年降水集中在4到9月份, 无霜期约120 d。研究区土壤类型为白浆土 (土壤理化性质见表 1),基底为玄武岩台地,母质粘重。土壤的机械组成以粗粉粒 (0.05—0.01 mm)、粘粒 ( < 0.001 mm) 为主[14]。该地区地带性植被 (顶级群落) 为红松阔叶林[15], 主要树种有红松 (Pinus koraiensis)、紫椴 (Tilia amurensis)、黑桦 (Betula dahurica)、蒙古栎 (Quercus mongolica)、水曲柳 (Fraxinus mandschurica) 胡桃楸 (Juglans mandshurica) 和白桦 (Betula platyphylla)、黄菠罗 (Phellodendron amurense) 等。

|
| 图 1 研究区位置图 Fig. 1 Location of the study area |
| 样地代码Plot No. | 样地类型Plot type | pH | 有机质SOM/% | 全氮TN/(g/kg) | 碳氮比C:N | 土壤类型Soil type |
| 1 | 草地 | 5.3 | 6.8 | 7.9 | 12.6 | 白浆土 |
| 2 | 灌木林 | 5.4 | 8.32 | 8.2 | 13.7 | 白浆土 |
| 3 | 白桦林地 | 5.4 | 12.5 | 6.6 | 15.5 | 白浆土 |
| 4 | 阔叶杂木林地 | 5.2 | 10.2 | 8.1 | 11.1 | 白浆土 |
| 5 | 红松阔叶林地 | 4.9 | 11.17 | 10.6 | 12.9 | 白浆土 |
| SOM:有机质Soil organic matter;TN:全氮Total nitrogen;C:N:碳氮比C:N proportion | ||||||
由于长白山原始森林受到破坏程度和时间长短不同, 在长白山红松阔叶林常常形成不同的演替阶段。本次观测选取了温带森林演替序列的5个不同阶段:草地 (次生杂草地)、灌木林 (幼龄林) 地、白桦林地、阔叶杂木林地和红松阔叶林地, 作为测量样地 (各样地基本地理信息见表 2)。由于演替阶段的不同, 各样地的群落结构、植被组成均存在较大差异 (各样地群落组成及生长状况见表 3)。
| 样地代码 Plot No. |
样地类型 Plot type |
坐标Geographical coordinates | 海拔 Altitude/m |
坡度 Slope/(°) |
坡向 Aspect |
|
| N | E | |||||
| 1 | 草地 | 42°21′ | 128°09′ | 781 | 3.25 | NE |
| 2 | 灌木林 | 42°08′ | 128°12′ | 690 | 2.83 | NE |
| 3 | 白桦林地 | 42°11′ | 128°10′ | 809 | 4.32 | NE |
| 4 | 阔叶杂木林地 | 42°23′ | 128°01′ | 793 | 3.40 | NE |
| 5 | 红松阔叶林地 | 42°24′ | 128°05′ | 748 | 4.12 | NE |
| 主要因子 Important factor |
草地 Grassland |
灌木林 Shrub forest |
白桦林 Secondary birch forest |
阔叶杂木林 Broad-leaved mixed forest |
红松阔叶林 Broad-leaved Korean pine mixed forest |
|
| 林龄Age /a | * | 12 | 28 | 59 | 120 | |
| 郁闭度 | * | * | 0.82 | 0.65 | 0.81 | |
| 物种数 | * | * | 5 | 9 | 18 | |
| 平均株高/m | * | * | 15.3 | 22.5 | 26 | |
| 乔木层 | 平均胸径/cm | * | * | 10.6 | 16.5 | 19.4 |
| Arbor layer | 枝下高/m | * | * | 7.5 | 5.9 | 5.89 |
| 主要乔木 | * | 白桦、山杨、蒙古栎的少量幼苗 | 白桦、山杨、蒙古栎 | 紫椴、水曲柳、春榆、少量红松幼苗 | 红松、糠椴、水曲柳、紫椴、色木槭、春榆 | |
| 盖度 | * | 0.4 | 0.5 | 0.5 | 0.6 | |
| 灌木层 | 平均冠幅/cm | * | 21 | 25 | 41 | 58 |
| Shrub layer | 物种数 | * | 12 | 10 | 9 | 13 |
| 主要灌木 | * | 大蔷薇、细叶杜香、越桔等 | 毛榛子、绢毛绣线菊、珍珠梅、忍冬等 | 毛榛、刺五加、卫矛、暴马丁香、色木槭等 | 毛榛、刺五加、卫矛、东北山梅花等 | |
| 盖度 | 0.8 | 0.5 | 0.3 | 05 | 0.6 | |
| 草本层 | 物种数 | 8 | 10 | 14 | 17 | 32 |
| Herb layer | 主要草本及地被物 | 小叶草、苔草, 狼尾草, 珠芽蓼等 | 草本类有小叶草、苔草等;地衣植被是苔藓 | 小叶芹、毛缘苔草、木贼、蕨类等 | 东北羊角芹、小叶草、苔草、山茄子等 | 山茄子、白花延龄草、透骨草、东北羊角芹等 |
| *表示没有该类型 | ||||||
测定时间选在天气状况较好的生长季 (2013年8、9月), 使用美国LGR公司的便携式温室气体分析仪 (PGGA)(配有3个呼吸室), 对5样地同时野外原位连续测量CO2气体的浓度。监测时间为8:00—18:00, 每个样地选择3次重复。每个呼吸室的测量时间是120 s, 排空时间为60 s, 10 min为一个测量周期;测量前除去地表面覆盖物, 提前1d将3个PVC土壤环插入深度约为5 cm土层中, 尽量减少人为干扰并保留凋落物的自然状态。将仪器和3个全自动土壤呼吸室连接, 从而获得3个通道的监测相关数据。利用仪器所测的相关数据, 算出土壤呼吸通量, 求其平均值。
利用土壤三参数分析仪和袖珍气象追踪仪, 对土壤 (5 cm) 的温度、湿度以及林间的温度、湿度进行同步测量。
1.3 数据计算和统计分析土壤呼吸气体的通量是单位时间单位面积观测气室内该气体浓度的变化, 正值表示土壤排放某种气体, 负值表示土壤吸收某种气体。将仪器所测的数据代入公式进行计算, 从而求得气体的排放量。土壤CO2排放量的计算公式如下:
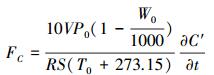
|
式中, F为单位时间单位面积土壤呼吸的气体排放量 (μmol m-2s-1), R为理想气体方程里的常量8.314, S为土环截面积 (cm2), V为气室内气体的体积 (cm3), P0为初始的气压值 (KPa), T0为初始的空气温度 (℃), W0为初始水汽的摩尔分数 (m mol mol-1), ∂C′/∂t为气室内被测气体随时间变化的体积混合比摩尔分数 (mol/s)。
本次实验实地所测的数据是在校正预处理中, 计算每个时间间隔 (30 min) 内所得数据的平均值, 文中均采用绝对通量 (即吸收速率) 进行图标绘制。数据采用Excel 2007和Spss 17.0等进行处理和计算, Origin 8.6做为绘图软件。
2 结果分析 2.1 不同样地土壤CO2排放通量的昼间动态变化不同样地的土壤CO2排放通量的昼间变化总体上表现为单峰曲线变化 (图 2), 整体来看, 其中最大值出现在温度较高时段, 如13:00—15:00, 最小值出现在温度较低的时段, 如早晨。草地和灌木林地的峰值大概在13:00左右, 明显提前于白桦林地、阔叶杂木林地和红松阔叶林地 (14:00—15:00左右)。其中红松阔叶林地其土壤CO2排放通量的规律跟其它样地的有一些不同, 最高值在15:00左右, 比其他几个样地明显推迟。这表明红松阔叶林地的土壤呼吸, 比较其他样地有较明显的滞后性特征。不同样地土壤呼吸CO2排放通量的昼间变化的最高值、最低值、平均值和变化幅度等存在明显差异 (图 3)。土壤CO2排放通量的最高值由低到高排列依次为草地 (3.215 μmol m-2 s-1)、灌木林地 (3.543 μmol m-2 s-1)、白桦林地 (3.787 μmol m-2 s-1)、阔叶杂木林地 (4. 767 μmol m-2 s-1)、红松阔叶林地 (5.152 μmol m-2 s-1)。土壤CO2排放通量的最低值由低到高排列依次为草地 (2.075 μmol m-2 s-1)、灌木林地 (2.110 μmol m-2 s-1)、白桦林地 (2.283 μmol m-2 s-1)、阔叶杂木林地 (2.487 μmol m-2 s-1)、红松阔叶林地 (4.130 μmol m-2 s-1);土壤CO2排放通量平均值由低到高排列依次为草地 (2.760 μmol m-2 s-1)、灌木林地 (2.854 μmol m-2 s-1)、白桦林地 (3.048 μmol m-2 s-1)、阔叶杂木林地 (3.696 μmol m-2 s-1)、红松阔叶林地 (4.61 μmol m-2 s-1)。对各样地的土壤CO2昼间排放通量对比表明, 平均排放通量、最大排放量、最小排放量的变化次序是一样的, 依次为草地<灌木林地<白桦林地<阔叶杂木林地<红松阔叶林地, 与群落演替次序大致相同。

|
| 图 2 5个实验样地土壤CO2排放通量的昼动态变化 Fig. 2 The five experimental plots diurnal variation of soil CO2fluxes |

|
| 图 3 不同样地土壤CO2排放通量日变化幅度 Fig. 3 Diurnal variation ranges of soil CO2 fluxes under different vegetation types 样地1代表草地, 样地2代表灌木林地, 样地3代表白桦林地, 样地4阔叶杂木林地, 样地5代表红松阔叶林地 |
5个样地的CO2排放通量均为正值 (表 4), 说明5个样地均为大气CO2的排放源。其中处在演替阶段后期的红松阔叶林地的CO2排放通量最大, 其次为白桦林地和阔叶杂木林地, 大概为其2/3, 草地和灌木林地CO2排放通量最小, 大概为其1/3。因此可知在自然状态下的土壤有无凋落物或者说是植被的覆盖率对土壤CO2排放通量影响很大。
| 样地类型 Plot type |
时段 Time |
平均排放通量 Average emissions flux/ (μmol m-2s-1) |
该时段排放总量 Total emissions/ (μmol/m2) |
| 草地Grassland | 8.00—18.00 | 2.760 | 99360 |
| 灌木林地Shrub forest | 8.00—18.00 | 2.854 | 102744 |
| 白桦林地Secondary birch forest | 8.00—18.00 | 3.048 | 109728 |
| 阔叶杂木林Broad-leaved mixed forest | 8.00—18.00 | 3.696 | 133056 |
| 红松阔叶林Broad-leaved Korean pine mixed forest | 8.00—18.00 | 4.610 | 165960 |
| *2013年8月采样及测定 | |||
5个样地气温和土壤温度 (5 cm) 均呈现单峰的昼间变化, 气温比土壤温度高且变化幅度更大, 土壤温度的最大值相比气温最大值, 有一定的滞后性。与土壤CO2排放通量的变化曲线对比, 发现土壤CO2排放通量和温度有一定的相关性, 其中土壤温度的变化曲线和CO2排放通量的变化趋势更为一致。说明与气温相比, 土壤温度更影响土壤的呼吸过程 (图 4)。

|
| 图 4 土壤CO2排放通量与气温、土壤温度的日变化动态 Fig. 4 Diurnal variation patterns of soil CO2 flux with air temperature and soil temperature (0—5cm) |
温度与土壤CO2排放量的关系采用指数回归方程进行分析, 方程如下:
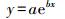
|
式中, y表示的是土壤CO2排放量, x为温度, a是0℃时的土壤呼吸, b是温度敏感系数。
应用Origin 8.6软件分别对5个样地土壤CO2排放通量与气温 (表 5) 和土壤温度进行回归分析 (图 5)。
| 样地类型 Plot type |
回归方程 Regression equation |
相关系数 R2 |
| 样地1(a) Plot 1(a) | y=1.28e0.037 x | 0.784* |
| 样地2(b) Plot 2(b) | y=1.42e0.041 x | 0.533* |
| 样地3(c) Plot 3(c) | y=1.729e0.046x | 0.407* |
| 样地4(d) Plot 4(d) | y=1. 259e0.046x | 0.355* |
| 样地5(e) Plot 5(e) | y=3.106e0.028x | 0.322* |
| * P<0.05 | ||

|
| 图 5 昼间土壤CO2排放通量与0—5cm土壤温度的响应曲线 Fig. 5 The response curve of soil CO2 flux with air temperature and soil temperature (0—5cm) on the diurnal scale |
5个样地的地表空气温度与土壤CO2排放通量回归方程是显著 (表 5), 但相关系数不高, 说明样地内土壤CO2排放通量与地表空气温度相关, 但相关性不高。
从图 5可以看出, 5个样地的土壤呼吸CO2排放通量都与土壤温度拟合较好且呈现明显的正相关, 随着土壤温度的升高, 土壤CO2排放量也增大。对土壤CO2排放量与土壤温度进行回归分析 (P<0.05), 结果是草地 (y=1.197e0.0429x, R2=0.861)、灌木林 (y=1.365e0.0464x R2=0.836)、白桦林 (y=0.769e0.0718x, R2=0.713)、杂木林 (y=1.318e0.0729x, R2=0.695)、红松阔叶林 (y=2.647e0.0412x, R20.648), 草地和灌木林的相关系数较高, 白桦林、杂木林和红松林的相关系数依次降低, 这表明从整体来看, 处在演替前期的土壤排放量与土壤温度的相关性要高于演替后期。可见, 植被群落的生长差异和所处的生长阶段对土壤CO2排放量有一定影响。
土壤呼吸过程的敏感性用Q10[16]表示, Q10是衡量土壤呼吸和土壤温度之间的重要参数, 表示温度每增加10℃, 土壤呼吸速率增加的倍数。计算公式如下:

|
式中, b是温度敏感系数。
经过计算, 草地土壤呼吸的Q10值为1.54、灌木林地土壤呼吸的Q10值为1.59、白桦林地土壤呼吸的Q10值为2.05、阔叶杂木林地土壤呼吸的Q10值为2.07, 红松阔叶林地土壤呼吸的Q10值为1.50, 可以看出各样地土壤呼吸过程的敏感性由大到小排序依次为红松阔叶林地 < 草地 < 灌木林地 < 白桦林地 < 阔叶杂木林。
2.4 湿度与土壤CO2排放通量的关系土壤湿度的日变化幅度较小, 其中在12:00左右, 土壤湿度最小 (图 6)。由于5个样地的植被生长不同, 林内的郁闭度不同, 因而5个样地的土壤湿度有一定的差异。样地1和样地2的土壤湿度最小, 样地5和样地4次之, 样地3的土壤湿度最大。

|
| 图 6 土壤CO2排放通量与土壤湿度的日变化动态 Fig. 6 Diurnal variation patterns of soil CO2flux and soil moisture |
本研究对5个样地的土壤湿度和土壤CO2排放通量的关系进行分析 (图 7), 发现5个不同立地条件下的土壤湿度与土壤CO2排放通量没有明显的相关关系。这说明湿度不是影响土壤呼吸的主要因素。这可能是由于本次实验的时间是在8月中旬到9月初, 在此期间长白山地区一直降水较多, 而长白山地区又属于温带湿润地区, 地下水也较丰富, 水分不是限制因素, 因此温度对于土壤CO2排放通量的影响大大超过了土壤水分的影响。

|
| 图 7 土壤CO2排放通量与土壤湿度的关系 Fig. 7 The relationship between soil CO2 flux and soil moisture |
许多研究已证明, 土地利用方式的改变会导致土壤碳通量的改变[17-19]。森林受到破坏后, 改变了地上生物量、凋落物量、光照条件, 从而影响林内小气候, 改变土壤的温度和湿度, 影响土壤的理化性质和土壤微生物种类; 进而影响土壤的呼吸, 导致森林的不同演替阶段下CO2排放通量差异较大。
(1) 温带森林不同演替阶段土壤CO2排放通量的昼动态变化
5个样地的CO2排放通量的昼间排放都呈现出明显的单峰型, 最大值都出现在温度较高时段, 最小值都出现在温度较低时段, 但是红松阔叶林地的土壤呼吸呈现出明显的滞后性, 这可能是因为红松针阔叶混交林冠层茂密、林相结构复杂, 土壤微生物和以及土壤表层的凋落物较多的缘故。大多数学者研究结果也表明, 土壤呼吸作用的日变化多呈单峰型曲线[20];土壤呼吸的最大值出现在12:00—14:00, 最小值出现在清晨[21]。一些学者研究发现, 红松针阔叶混交林比其他生态系统土壤呼吸的峰值有明显的延后, 大概在18:00左右, 森林生态系统复杂的层片结构和丰富的凋落物会对土壤呼吸速率的日动态产生“时滞效应”[13], 这是其他生态系统所没有的。
(2) 温带森林不同演替阶段的土壤CO2排放量的估算
本研究发现, 随着温带森林演替的正向进行, 土壤CO2排放通量依次增大, 次序为草地<灌木林地<白桦林地<阔叶杂木林地<红松阔叶林地。温带森林不同演替阶段下的土壤CO2排放通量具有统一性, 均表现为CO2的源。温带森林不同演替阶段下的土壤CO2排放通量的统一性, 确保了小的时段 (如昼间) 观测能通过换算, 为实现CO2排放量的估算提供依据。虽然有相关研究认为树种和林型对森林土壤CO2的排放通量无显著的影响[22], 在年尺度范围内, 不同林龄的林地土壤CO2排放通量之间的平均值无明显差异[23], 也有一些研究发现,植被稀疏或者经过灭活处理的土壤,虽然在白天释放CO2,但是夜间却从大气吸收一部分CO2,总体从全天看可表现为净吸收CO2[24-26]。但从本研究来看, 不同群落下的土壤CO2的排放通量还是有较明显的差异:处于演替初期的生态系统, 如草地, 处于固碳阶段, 植被稀疏、土壤微生物比较少、根呼吸和微生物呼吸都较弱, 所以对CO2释放的贡献率较小;而处于演替后期的生态系统, 如红松阔叶林, 处于碳收支平衡阶段, 森林群落地表凋落物较多、物种丰富、土壤中微生物也比较多、呼吸活动较强, 因此对CO2释放的贡献率最大。
据相关统计, 世界温带森林土壤的CO2年平均释放量为2511 g m-2a-1 [8], 以这个数据为基础, 参照研究区各样方昼间平均排放量, 可以大概计算出草地的土壤CO2年排放量为1503 g m-2a-1, 灌木林地的土壤CO2排放量为1554 g m-2a-1, 白桦林地的土壤CO2年排放量为1660 g m-2a-1, 阔叶杂木林地的土壤CO2的年排放量为2013 g m-2a-1。估算的白桦林地的土壤CO2年排放量与刘绍辉等[27]测得的温带白桦林年排放量为1132 g m-2a-1比较接近。
(3) 温带森林不同演替阶段土壤呼吸的影响因素
大部分的学者研究都表明, 温度是影响长白山红松阔叶林土壤呼吸最重要的因素[12, 13, 28-31]。环境因子中, 温度和湿度都是土壤呼吸的影响因子, 但是土壤呼吸对温度的敏感性大于对湿度的敏感性。通过分析Q10, 发现各样地土壤呼吸过程的敏感性由大到小排序依次为红松阔叶林地 < 草地 < 灌木林地 < 白桦林地 < 阔叶杂木林, 这表明在不同演替阶段的土壤呼吸中, 红松阔叶林对温度的变化敏感度不高, 次生林对温度的变化较为敏感。温度监测对土壤CO2排放量估算作用明显。
通过把5个样地的土壤CO2排放通量分别与气温和土壤 (5 cm) 温度进行指数拟合, 发现土壤温度比气温更能解释土壤CO2排放通量的变异, 土壤温度与土壤CO2排放通量的指数关系拟合更好, 相关系数更高。这也与大多数学者研究结果一致, 如刘颖, 韩士杰等[28, 30]观测到长白山地区长白松林地的土壤呼吸与土壤温度之间呈显著的指数相关关系;王光军, 田大伦等[32]通过对亚热带杉木和马尾松林地土壤呼吸的研究, 得出土壤温度可以分别解释土壤呼吸变化的91.7%和78%, 土壤呼吸速率与土壤温度呈显著的指数相关性。王德宣等[33]在若盖尔高原的研究中也表明沼泽湿地和草地的CO2排放量的日变化与气温呈现正相关。研究表明土壤表层温度变化对土壤呼吸排放通量的敏感性更高[34], 因为对土壤呼吸起作用的微生物和植物根系主要分布在土壤的表层, 土壤温度升高可以促进微生物的活动和植物根系的呼吸作用[35]。
土壤呼吸是一个复杂的过程, 其中土壤中的微生物呼吸作用和植物的根系呼吸对土壤呼吸的贡献最大, 而这些都与土壤水分密切相关。有相关研究认为, 森林土壤CO2排放量在一定范围内是随着温度和水分的增加而增加的, 如刘硕等[36]在研究中表明北京温带森林3种林型土壤CO2排放量随着土壤含水量的增加而增大。土壤湿度与土壤CO2排放通量的关系较为复杂, 因为土壤湿度的变化还受其它外界因素的影响, 如土壤温度、大气的温度、降水、林内的郁闭度等, 这些因素在不同的地方对土壤湿度的影响程度不同, 这都增加了土壤湿度对土壤呼吸影响的研究难度。但是大部分研究都表明, 土壤湿度只有在最低或最高的情况下, 才会影响土壤呼吸过程, 抑制CO2排放通量[36]。也有研究表明, 温度和湿度对土壤呼吸的作用大部分时候是交互进行的:当温度低于5℃时, 土壤呼吸对湿度的反应不敏感;在温度为10—20℃时反应敏感, 同样当含水量低于体积含水量的7.5%, 土壤呼吸对温度的反应不敏感, 但含水量是体积含水量的10%—25%时反应敏感[38]。
(4) 问题和展望
本研究同时对温带森林不同演替阶段的土壤CO2昼间排放通量进行观测, 实现不同演替阶段下的土壤CO2排放量的估算, 估算值与他人的监测值吻合较好。今后应扩大到整个生长季的观测, 提高估算的精度, 并对比其他生长期与生长旺盛时期的差别, 最终确定温带森林不同演替阶段的土壤CO2年排放量的高效率观测方法。
| [1] | Vitousek P M. Beyond global warming: ecology and global change. Ecology, 1994, 75(7): 1861–1876. DOI:10.2307/1941591 |
| [2] | Jenkinson D S, Adams D E, Wild A. Model estimates of CO2 emissions from soil in response to global warming. Nature, 1991, 351(6324): 304–306. DOI:10.1038/351304a0 |
| [3] | Granier A, Ceschia E, Damesin C, Dufrêne E, Epron D, Gross P, Lebaube S, Le Dantec V, Le Goff N, Lemoine D, Lucot E, Ottorini J M, Pontailler J Y, Saugier B. The carbon balance of a young Beech forest. Functional Ecology, 2000, 14(3): 312–325. DOI:10.1046/j.1365-2435.2000.00434.x |
| [4] | 顾峰雪, 黄玫, 张远东, 李洁, 闫慧敏, 郭瑞, 钟秀丽. 氮输入对中国东北地区土壤碳蓄积的影响. 生态学报, 2016, 36(17). DOI:10.5846/stxb201502260389 |
| [5] | 刘绍辉, 方精云. 土壤呼吸的影响因素及全球尺度下温度的影响. 生态学报, 1997, 17(5): 469–476. |
| [6] | Tang J W, Baldocchi D D, Xu L K. Tree photosynthesis modulates soil respiration on a diurnal time scale. Global Change Ecology, 2005, 11(8): 1298–1304. DOI:10.1111/gcb.2005.11.issue-8 |
| [7] | Hibbard K A, Law B E, Reichstein M, Sulzman J. An analysis of soil respiration across northern hemisphere temperate ecosystems. Biogeochemistry, 2005, 73(1): 29–70. DOI:10.1007/s10533-004-2946-0 |
| [8] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B, 1992, 44(2): 81–99. DOI:10.3402/tellusb.v44i2.15428 |
| [9] | 王新源, 李玉霖, 赵学勇, 毛伟, 崔夺, 曲浩, 连杰, 罗永清. 干旱半干旱区不同环境因素对土壤呼吸影响研究进展. 生态学报, 2012, 32(15): 4890–4901. |
| [10] | Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7–20. DOI:10.1023/A:1006247623877 |
| [11] | 阳小成, 阿舍小虎, 苗原, 刘银占. 川西北高寒草甸不同放牧模式对土壤呼吸的影响. 生态学报, 2016, 36(17). DOI:10.5846/stxb201502120343 |
| [12] | 王旭, 周广胜, 蒋延玲, 曹铭昌, 曾伟, 宋健. 长白山阔叶红松林皆伐迹地土壤呼吸作用. 植物生态学报, 2007, 31(3): 355–362. |
| [13] | 王旭, 周广胜, 蒋延玲, 李峰. 长白山红松针阔混交林与开垦农田土壤呼吸作用比较. 植物生态学报, 2006, 30(6): 887–893. |
| [14] | 律兆松, 徐琪. 中国白浆土研究Ⅰ.白浆土机械组成特点及元素地球化学分异特征. 土壤学报, 1993, 30(3): 274–288. |
| [15] | 郭利平, 姬兰柱, 王珍, 王智轩. 长白山红松阔叶林不同演替阶段优势种的变化. 应用生态学报, 2011, 22(4): 866–872. |
| [16] | Chen H, Harmon M E, Griffiths R P, Hicks W. Effects of temperature and moisture on carbonrespired from decomposing woody roots. Forest Ecology and Management, 2000, 138(1/3): 51–64. |
| [17] | Upadhyay T P, Solberg B, Sankhayan P L, Shahi C. Land-use changes, forest/soil conditions and carbon sequestration dynamics: a bio-economic model at watershed level in Nepal. Journal of Bioeconomics, 2013, 15(2): 135–170. DOI:10.1007/s10818-011-9123-z |
| [18] | Fernandez C W, Kennedy P G. Moving beyond the black‐box: fungal traits, community structure, and carbon sequestration in forest soils. New Phytologist, 2015, 205(4): 1378–1380. DOI:10.1111/nph.13289 |
| [19] | Hagedorn F, Maurer S, Egli P, Blaser P, Bucher J B, Siegwolf R. Carbon sequestration in forest soils: effects of soil type, atmospheric CO2 enrichment, and N deposition. European Journal of Soil Science, 2001, 52(4): 619–628. DOI:10.1046/j.1365-2389.2001.00412.x |
| [20] | 韩广轩, 周广胜. 土壤呼吸作用时空动态变化及其影响机制研究与展望. 植物生态学报, 2009, 33(1): 197–205. |
| [21] | 于爱忠, 黄高宝, 柴强. 不同耕作措施对西北绿洲灌区冬小麦农田土壤呼吸的影响. 草业学报, 2012, 21(1): 273–278. DOI:10.11686/cyxb20120135 |
| [22] | Fernandez I J, Son Y, Kraske C R, Rustad L E, David M B. 1993. Soil carbon dioxide characteristics under different forest types and after harvest. Soil Science Society of America Journal, 1992, 57(4): 1115-1121. |
| [23] | 孙向阳, 乔杰, 谭笑. 温带森林土壤中的CO2排放通量. 东北林业大学学报, 2001, 29(1): 34–39. |
| [24] | Persons A N, Barrett J E, Wall D H, Virginia R A. Soil carbon dioxide flux in Antarctic Dry Valley ecosystems. Ecosystems, 2004, 7(3): 286–295. |
| [25] | Xie J W, Li Y, Zhai C X, Li C H, Lan Z D. CO2 absorption by alkaline soils and its implication to the global carbon cycle. Environmental Geology, 2009, 56(5): 953–961. DOI:10.1007/s00254-008-1197-0 |
| [26] | Hamerlynck E P, Scott R L, Sánchez-Cañete E P, Barron-Gafford G A. Nocturnal soil CO2 uptake and its relationship to subsurface soil and ecosystem carbon fluxes in a Chihuahuan Desert shrubland. Journal of Geophysical Research, 2013, 118(4): 1593–1603. |
| [27] | 刘绍辉, 方精云, 清田信. 北京山地温带森林的土壤呼吸. 植物生态学报, 1998, 22(2): 119–126. |
| [28] | 刘颖, 韩士杰. 长白山典型森林生态系统土壤呼吸的季节动态. 东北林业大学学报, 2012, 40(11): 107–110. DOI:10.3969/j.issn.1000-5382.2012.11.026 |
| [29] | 梁宇, 许嘉巍, 胡远满, 常禹, 布仁仓. 长白山阔叶红松林退化生态系统的土壤呼吸作用. 应用生态学报, 2010, 21(5): 1097–1104. |
| [30] | 刘颖, 韩士杰. 长白山四种森林土壤呼吸的影响因素. 生态环境学报, 2009, 18(3): 1061–1065. |
| [31] | 王淼, 刘亚琴, 郝占庆, 王跃思. 长白山阔叶红松林生态系统的呼吸速率. 应用生态学报, 2006, 17(10): 1789–1795. DOI:10.3321/j.issn:1001-9332.2006.10.004 |
| [32] | 王光军, 田大伦, 闫文德, 朱凡, 李树战. 亚热带杉木和马尾松群落土壤系统呼吸及其影响因子. 植物生态学报, 2009, 33(1): 53–62. |
| [33] | 王德宣, 宋长春, 王毅勇, 赵志春. 若尔盖高原沼泽湿地与草地二氧化碳通量的比较. 应用生态学报, 2008, 19(2): 285–289. |
| [34] | 贾淑霞, 王政权, 梅莉, 孙玥, 全先奎, 史建伟, 于水强, 孙海龙, 谷加存. 施肥对落叶松和水曲柳人工林土壤呼吸的影响. 植物生态学报, 2007, 31(3): 372–379. |
| [35] | 杨庆朋, 徐明, 刘洪升, 王劲松, 刘丽香, 迟永刚, 郑云普. 土壤呼吸温度敏感性的影响因素和不确定性. 生态学报, 2011, 31(8): 2301–2311. |
| [36] | 刘硕, 李玉娥, 孙晓涵, 万运帆, 高清竹, 秦晓波, 马欣. 温度和土壤含水量对温带森林土壤温室气体排放的影响. 生态环境学报, 2013, 22(7): 1093–1098. |
| [37] | Luo Y Q, Zhou X H. Soil Respiration and the Environment. San Diego, California: Academic Press/Elsevier, 2006. |
| [38] | Carlyle J C, Ba Than U. Abiotic controls of soil respiration beneath an eighteen-year-old Pinus radiata stand in south-eastern Australia. Journal of Ecology, 1988, 76(3): 654–662. DOI:10.2307/2260565 |