 2017, Vol. 37
2017, Vol. 37文章信息
- 欧阳林男, 吴晓芙, 陈永华, 李芸, 冯冲凌, 陈明利
- OUYANG Linnan, WU Xiaofu, CHEN Yonghua, LI Yun, FENG Chongling, CHEN Mingli.
- 锰矿修复区植物生态系统自由能与化学势分析
- Gibbs free energy and chemical potentials of plant communities in a manganese-ore tailing site undergoing ecological restoration
- 生态学报. 2017, 37(8): 2694-2705
- Acta Ecologica Sinica. 2017, 37(8): 2694-2705
- http://dx.doi.org/10.5846/stxb201512032425
-
文章历史
- 收稿日期: 2015-12-03
- 网络出版日期: 2016-10-29





2. 环境科学与工程研究中心, 长沙 410004
2. Research Center of Environmental Science and Engineering, Changsha 410004, China
生态系统是物质与能量交换的场所, 其状态可以用热力学理论来描述。生物因子是生态系统的核心组成分, 生态系统能态变化过程表现为系统生物量与生物组成的变化, 不同立地环境中生态系统能态的差异反映立地条件的差异, 在给定立地条件下, 系统中某一物种的能态反映该一物种对给定立地环境的适应性。在生态系统热力学领域, 生态能质 (eco-exergy) 理论[1-2]被广泛应用到生态系统生产力[3-5]、物种丰富度[6-7]、生态系统健康状态[8-11]等的分析和评价中。然而涉及生态系统自由能和化学势分析方面的研究尚未见报道。
国内外筛选重金属超富集植物常用的两个指标为植物重金属元素浓度C和转移系数TF[12-13]。采用C和TF指标可以直接比较个体物种耐受和积累污染元素的生理生化特性, 由于C、TF是个体水平上的测试指标, 则其不足之处是不能反应物种在重金属污染环境中的生态学特性。在给定面积的区域里, 自然萌发生长植物种类个体的C和TF值大, 其群体水平上的密度d、单位面积生物量B和污染元素吸收量U却不一定大, 而d、B、U等指标分别反应物种的自然生态位、初级生产力和固定污染元素的能力。另外, 在重金属污染区生态修复中, 复合植物群落模式在生态系统结构与功能稳定性、生物多样性、植被覆盖率、植物群落景观效应以及系统固定和去除重金属能力等各个方面都要优于单一植物模式。显然, 在模式植物筛选中, 仅知道物种个体的浓度C、转移系数TF的数据不够, 还需要知道给定区域单位面积上物种的生物量, 密度, 以及其与系统物种之间的竞争、共生、抑制、互利等关系行为。因此, 在描述和评价生态系统的结构功能和物种的生态学特性时, 需要包含个体和群体尺度指标综合信息的复合变量。
作为生态系统的容量和强度因子, Gibbs自由能G和化学势μ是包含d、B、U和物种数S变量的状态函数。区间和物种间G和μ值的差异分别反映区间土壤环境和物种特性的差异, 因此, G和μ值在评价重金属污染区修复效果和筛选修复植物种类中具有一定的理论和实用价值。本文研究的重点是, 基于生态过程是物质与能量交换过程的特点, 运用热力学原理建立生态系统Gibbs自由能G和化学势μ方程;依据给定温度T和压力P条件下组分标准化学势μ0值为常数, 自由能G和化学势μ的绝对值不可知, 但差异可以确定等原理, 解决自由能比和化学势差的计算方法;应用建立的方程分析、比较、评价锰矿污染区植物修复效果;以物种间化学势差作为评价指标筛选优势修复植物。
1 材料与方法 1.1 试验区概况试验区为湘潭鹤岭锰矿尾矿渣堆积废弃地, 低山丘陵地貌, 112°45′—112°55′E, 27°53′—28°03′N, 亚热带季风气候, 日最高气温42.2 ℃, 最低-8 ℃, 年均气温为17.4 ℃;年降水量最高1912.2 mm, 最低1185.5 mm, 年均降水量为1431.4 mm, 降水主要集中在4—7月;年蒸发量最大1580.9 mm, 最小992.2 mm, 年均蒸发量为1321.7 mm。试验修复区为示范工程区, 面积4.53 hm2, 工程地污染物组成复杂, 主要是矿石废弃物、矿渣和选矿后的尾矿泥, 土壤约占混合污染物的40%—50%。试验地养分与污染状况见表 1, 分析数据表明试验工程区养分贫瘠, 复合重金属污染严重。锰元素含量平均值高达24213 mg/kg。试验对照区与修复区相邻, 土壤污染物组成和水热条件与修复区基本一致。
| 测试项目 Analyzed items |
pH | 总氮/(g/kg) Total nitrogen |
总磷/(g/kg) Total phosphorus |
总钾/(g/kg) Total potassium |
重金属总量/(mg/kg) Total heavy metal content |
|||
| Mn | Pb | Cd | Cu | |||||
| 样本Samples | 6.96 | 0.77 | 0.11 | 23.36 | 24213 | 261.6 | 3.30 | 143.60 |
| 中国耕地Farmland in China | 4.50—9.10 | 1.00—2.09 | 0.44—0.85 | 5.00—30.00 | - | - | - | - |
| 中国背景值 Background ofChina | - | - | - | - | 583.0 | 26.0 | 0.1 | 22.6 |
项目组于2009年秋季整地, 2010年春季以泡桐 (Paulownia fortunei) 和栾树 (Koelreuteria bipinnata) 作为建群树种, 在试验点上建立了4 hm2生态修复示范工程区, 污染区不覆土, 为了改良植物根际环境, 苗木移植前每株根际定量施用了含有优势抗性菌株的专用有机肥。泡桐和栾树种植方法为挖穴 (0.5 m×0.5 m×0.5 m), 穴内施入专用有机菌肥作为底肥, 底肥与废弃地基质均匀混合, 比例为1:4。有机菌肥主要成分见表 2。有机菌肥中含有自工程区锰矿渣分离纯化后筛选得到的土著优势抗性菌种, 杆菌, 革兰氏阴性, 具有很强的耐受和去除Mn的能力[14]。示范工程建设前采用矿渣作为基质开展了两年的盆栽试验, 在此基础上筛选了树种, 验证了专用有机菌肥改良效果, 确定了示范工程用量。2010年3月在工程区混合间种1年生泡桐和栾树实生苗, 行距为2.0 m×2.5 m, 试验设置的对照区本底条件与修复区相同, 差别是对照区泡桐和栾树的根际施用了等量化肥。
| 土著优势抗性菌株 Native dominant resistence becteria |
吸附介质 Adsorption media |
总氮 TN |
总磷 TP |
有机质 Organic matter |
| 5 | 42 | 1.80 | 3.10 | 47.90 |
试验区整地前按梅花布点法采集锰尾矿渣样本, 其重金属浓度和营养元素含量见表 1。整地种植后, 在修复区和对照区分别设置固定3个20 m×30 m面积的乔木样方, 每个乔木样方内固定3个5 m×5 m面积的灌木样方, 12个1 m×1 m面积的草本样方。在每块样方中按物种分别选取乔木5株、灌木5株、草本植物7株做为标准样株, 以代表整体植物群落的平均水平。统计所有地表植物种类, 测量植物株高、胸径、地径、株数、盖度。乔灌木分别取根、茎、叶样本, 草本植物连根拔起, 按物种将根、茎、叶同部位混合得到植物分析样本。植物与基质样本采集时间为每年5—6月和8—9月。
1.3.2 样品测定植物样品用超纯水反复洗净, 风干、杀青后, 置80 ℃烘干至恒重, 获得植物干物质生物量, 样品粉碎过100目筛, 装袋密封干燥保存;土壤样品在室内风干, 剔除动植物残体, 磨碎过100目筛, 装袋密封干燥保存。植物与土壤样品均采用湿法消解, 用原子吸收分光光度法 (AA-7000型) 测定Mn、Pb、Cd、Cu、Zn、Cr、K的浓度。基质pH值、总氮、总磷、有机质测定分别采用电位法、半微量凯氏法、钼锑抗比色法和重铬酸钾水浴法。分析取3个平行样, 用平行样平均值作为基础数据, 平行误差范围在5%—15%内。
1.3.3 系统植物种类数和生物量测定在现场调查, 逐一记录修复区和对照区中所有物种。
通过测定标准样株的叶、茎和根的生物量, 得到标准样株单株的平均生物量, 将样地中植物的株数按面积换算成每公顷的株数, 得到密度d, 以密度乘以平均单株生物量计算出每公顷物种的生物量, 系统 (总) 生物量为所有植物种类生物量之和。
1.4 自由能与化学势计算方法 1.4.1 生态系统自由能依据热力学原理[15-16], 在给定温度 (T) 和压力 (P) 条件下, 单位面积生态系统给定状态点上某一物种的自由能 (Gi) 和生态系统的自由能 (G) 分别为
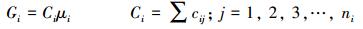
|
(1) |
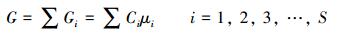
|
(2) |
式中, 取决于研究对象, cij可为物种i第j株的生物量或体内所含某种特殊物质 (或元素例如锰) 量, ni为物种i的密度 (单位面积上物种i的株数), S为单位面积系统中的物种数, Ci为物种i总生物量或所含特殊物质总量, μi为物种i(与Ci相关) 的化学势。物理化学中将物质的化学势定义为Gi对Ci的偏导数:
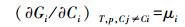
|
式中, 下注T、p、Cj表示给定温度T、压力p和物种Cj。化学势μi与活度 (ai)、活度系数 (γi) 和摩尔分数 (xi) 的关系为:
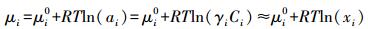
|
(3) |
式中, R为气体常数, T为绝对温度, μi0代表标准化学势, 为给定温度和标准压力条件下摩尔分数xi=1时的μi值。系统组分摩尔分数xi的定义为
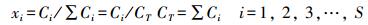
|
(4) |
式中, CT为系统生物总量或所含物质总量。用xi代替ai代入式1、2, 有:
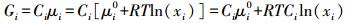
|
(5) |
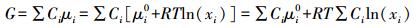
|
(6) |
参考μi的物理化学意义, 当Ci代表物种生物量时, 强度因子μi可视为生态系统中物种i的生长势, 而当Ci为物种所含某种物质量时, μi则可视为生态系统中物种i积累或合成该物质的势能。基于上述关系, 若标准化学势μi0已知, 则可通过测定Ci来计算物种和生态系统的μ和G值。
1.4.2 系统自由能比值对于给定生态系统, 当不区别物种差异, 仅以系统积累或合成某物质的能力作为研究对象时, 则系统唯一的组分因子为系统该物质 (例如锰元素) 的摩尔量。依据定义, 给定温度T与压力p, 物质的摩尔自由能为定值, 则所有物种所含该物质的标准化学势相等,
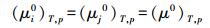
|
即给定T、p, 物质的标准化学势μ0为常数, 不随生态系统和物种变化。而当以系统积累某物质的能力作为研究对象时, 系统只有一个物质组分, 组分的摩尔量既为系统的摩尔总量CT, 组分的摩尔分数xi等于1,
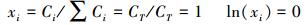
|
综合上述, 由式6可给出系统该物质的Gibbs自由能
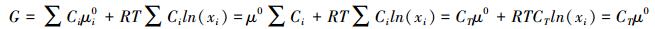
|
(7) |
另一方面, 当不区别物种差异, 仅以系统生物量作为研究对象时, 对于任一生态系统, 有理由假设系统中所有物种的干物质摩尔生物量的能值相等, 从属于这一假设前提, 给定T, p时, 所有物种生物干物质的标准化学势也相等, μ0值为常数, 不随生态系统和物种变化而变化。将生物量作为系统组分时, 系统也只有单一组分, 组分摩尔量等于系统总摩尔量, 摩尔分数x也等于1。因此, 式7也可用于计算系统生物量对Gibbs自由能的贡献。
基于式7, 可给出生态系统Ⅰ与Ⅱ的G值差异和比值
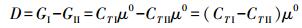
|
(8) |
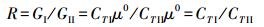
|
(9) |
上式说明, 系统间生物量差不等于Gibbs自由能差, 但系统生物量比等于系统Gibbs自由能比。系统G的比值R无量纲, R大于1, 说明系统Ⅰ的Gibbs自由能态高于系统Ⅱ。式9不能用以计算物种间G的比值, 原因是xi≠xj≠1, ln (xi)≠0。
1.4.3 化学势差异物种对系统自由能的贡献与物种的化学势呈正相关。当同一物种在不同立地环境中的Ci存在差异, 则其化学势μi也必然存在差异, 另一方面, 在同一立地环境中生长的不同物种的Ci及μi也会存在差异, 用ΔμⅠ-Ⅱ表示这一差异, 有

|
式中, 下注Ⅰ和Ⅱ可分别代表立地环境Ⅰ、Ⅱ或物种Ⅰ、Ⅱ。依据前面关于标准化学势的讨论, 无论Ci代表物种干物质生物量还是所含某种物质量, 给定T、p条件下, μ0为定值, 上式中μⅠ0-μⅡ0=0,
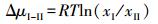
|
(10) |
式10表明, 物种间或物种在不同立地环境中生长或积累和合成某一物质势能的差异可唯一由相应xi比值的自然对数值所决定, 物种xi值大, 说明物种潜在势能大或物种的立地环境好。因此, 式10既可用以分析环境对植物生长的影响, 也可用以比较物种对环境的适应能力。
式10的应用要求给定温度条件, 而植物生长和生态系统状态变化通常不是等温过程, 为消除温度影响和计算方便, 可将上式除以RT转化为无量纲比值δ:
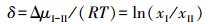
|
(11) |
在同一系统中, 总生物量和总锰吸收量CT为定值, 物种i和物种j的x比值为:
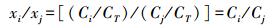
|
当用来比较同一系统中物种i和物种j的化学势差异时, 式11可转化为:
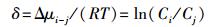
|
采用系统平均值作为对比参照值, 可令:
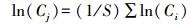
|
则:
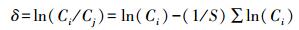
|
(12) |
应用式12可计算出同一区中所有物种的δ值, 若δ为正值, 说明该物种的生物量或锰吸收量大于系统的平均值, δ越大, 则物种的潜在势能越高, 对环境的适应能力越强。
1.4.4 TF值、优势度和simpson指数
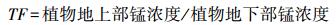
|
(13) |
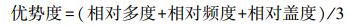
|
(14) |
式中, 相对多度等于第i个物种的个体数除以群落中所有物种的个体数的和;相对频度等于第i个物种在统计样方中出现的次数除以所有种出现的总频数;相对盖度等于第i个物种的盖度除以所有种的总盖度。
Simpson指数Simpson index[18]的计算公式如下:
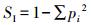
|
(15) |

|
(16) |
式中,ni指样地中第i个植物种的株数, N指样地中所有植物的总株数。
2 结果与分析 2.1 试验区植物系统自由能比值示范工程试验区 (Ⅰ) 造林后第2年泡桐和栾树的成活率在83%以上, 对照区泡桐和栾树种植后第2年成活率很低, 到后期全部枯死。示范工程建成后5 a期间修复区与对照区均陆续自然萌发生长了许多本土植物种类。依据土壤种子库原理[19-21], 给定区域的土壤种子库存信息容量相同[22-23], 因此, 同一区域不同区间自然萌发植物生长差异则唯一取决于区间土壤条件的差异, 而区内物种间生长状态的差异则反映物种对给定区域土壤环境的适应性。
表 3给出了修复区 (Ⅰ) 与对照区 (Ⅱ) 植物种的物种数量S、生物量B、锰吸收量U、密度d、覆盖度、生物量对数值lnB和锰吸收量对数值lnU。表 4给出了Ⅰ和Ⅱ区系统的总生物量∑Bi、总锰吸收量∑U、总密度∑d、总盖度, 以及两区总生物量之差DB、总锰吸收量之差DU、总生物量的比值RB和总锰吸收量的比值RU。
| 区域 Sites |
植物种类 Plant species |
植物种类编号 Serial number of plant species |
物种生物量B Biomass/ (kg/hm2) |
锰吸收量U Manganese uptake/ (g/hm2) |
密度d Density/ hm2 |
盖度 Coverage/% |
lnB | lnU |
| Ⅰ | 泡桐Paulownia fortunei | 1 | 19486.29 | 2676.15 | 833 | 89.54 | 9.88 | 7.89 |
| 栾树Koelreuteria bipinnata | 2 | 1594.37 | 208.91 | 867 | 68.08 | 7.37 | 5.34 | |
| 构树Broussonetia papyrifera | 3 | 365.27 | 203.63 | 1469 | 5.66 | 5.90 | 5.32 | |
| 狗牙根Cynodon dactylon | 4 | 285.66 | 199.23 | 313333 | 29.99 | 5.65 | 5.29 | |
| 芒Miscanthus sinensis | 5 | 266.94 | 79.35 | 39567 | 1.24 | 5.59 | 4.37 | |
| 商陆Phytolacca acinosa | 6 | 117.43 | 194.27 | 3022 | 7.02 | 4.77 | 5.27 | |
| 土荆芥Chenopodium ambrosioides | 7 | 92.50 | 19.14 | 4667 | 3.59 | 4.53 | 2.95 | |
| 灯心草Juncus effusus | 8 | 89.98 | 55.51 | 135556 | 3.73 | 4.50 | 4.02 | |
| 一年蓬Erigeron annuus | 9 | 88.89 | 104.76 | 51111 | 2.44 | 4.49 | 4.65 | |
| 香丝草Conyza bonariensis | 10 | 82.45 | 12.23 | 4533 | 1.03 | 4.41 | 2.50 | |
| 早熟禾Poa annua | 11 | 74.46 | 53.41 | 97778 | 2.19 | 4.31 | 3.98 | |
| 葎草Humulus scandens | 12 | 74.55 | 30.88 | 6117 | 0.13 | 4.31 | 3.43 | |
| 小蓬草Conyza canadensis | 13 | 61.74 | 27.88 | 131111 | 0.03 | 4.12 | 3.33 | |
| 裂叶悬钩子Rubus howii | 14 | 61.87 | 49.01 | 40494 | 4.28 | 4.12 | 3.89 | |
| 苍耳Xanthium sibiricum | 15 | 54.68 | 17.68 | 19266 | 0.65 | 4.00 | 2.87 | |
| 盐肤木Rhus chinensis | 16 | 54.35 | 23.31 | 3704 | 2.59 | 4.00 | 3.15 | |
| 钩吻Gelsemium elegans | 17 | 46.15 | 13.53 | 37777 | 1.78 | 3.83 | 2.60 | |
| 小巢菜Vicia hirsuta | 18 | 36.68 | 7.09 | 98556 | 15.33 | 3.60 | 1.96 | |
| 剪股颖Agrostis matsumurae | 19 | 30.17 | 12.60 | 95556 | 1.39 | 3.41 | 2.53 | |
| 野胡萝卜Daucus carota | 20 | 29.96 | 5.49 | 6667 | 0.19 | 3.40 | 1.70 | |
| 酢浆草Oxalis corniculata | 21 | 27.46 | 17.46 | 108889 | 2.45 | 3.31 | 2.86 | |
| 马兰Kalimeris indica | 22 | 26.84 | 8.82 | 4333 | 0.12 | 3.29 | 2.18 | |
| 苔草Carex tristachya | 23 | 25.62 | 12.89 | 66667 | 1.39 | 3.24 | 2.56 | |
| 狗尾草Setaria viridis | 24 | 25.21 | 18.14 | 46667 | 12.63 | 3.23 | 2.90 | |
| 艾Artemisia argyi | 25 | 23.87 | 5.78 | 27355 | 1.51 | 3.17 | 1.75 | |
| 野蔷薇Rosa multiflora | 26 | 22.60 | 17.67 | 7531 | 0.94 | 3.12 | 2.87 | |
| 蒌蒿Artemisia selengensis | 27 | 22.18 | 17.84 | 37778 | 2.28 | 3.10 | 2.88 | |
| 野豌豆Vicia sepium | 28 | 22.00 | 3.45 | 12000 | 17.06 | 3.09 | 1.24 | |
| 蓼Polygonum flaccidum | 29 | 15.78 | 2.63 | 1667 | 0.11 | 2.76 | 0.97 | |
| 大青Clerodendrum cyrtophyllum | 30 | 15.80 | 12.41 | 3333 | 0.43 | 2.76 | 2.52 | |
| 荩草Arthraxon hispidus | 31 | 14.92 | 5.65 | 55556 | 0.78 | 2.70 | 1.73 | |
| 水芹Oenanthe javanica | 32 | 14.68 | 1.10 | 4444 | 0.06 | 2.69 | 0.09 | |
| 鳢肠Eclipta prostrata | 33 | 12.84 | 2.73 | 4444 | 0.28 | 2.55 | 1.00 | |
| 台湾翅果菊Pterocypsela formosana | 34 | 11.13 | 1.90 | 722 | 0.53 | 2.41 | 0.64 | |
| 风轮菜Clinopodium chinense | 35 | 9.67 | 2.82 | 57778 | 0.78 | 2.27 | 1.04 | |
| 龙葵Solanum nigrum | 36 | 9.45 | 7.95 | 2546 | 1.13 | 2.25 | 2.07 | |
| 野燕麦Avena fatua | 37 | 9.41 | 1.61 | 7000 | 0.11 | 2.24 | 0.48 | |
| 飞蓬Erigeron acer | 38 | 3.19 | 2.56 | 13333 | 0.29 | 1.16 | 0.94 | |
| 鸡眼草Kummerowia striata | 39 | 3.19 | 1.93 | 2222 | 0.01 | 1.16 | 0.66 | |
| 青蒿Artemisia carvifolia | 40 | 2.30 | 0.52 | 2222 | 0.03 | 0.83 | -0.66 | |
| 蛇莓Duchesnea indica | 41 | 2.01 | 0.98 | 6667 | 0.14 | 0.70 | -0.02 | |
| 画眉草Eragrostis pilosa | 42 | 1.92 | 1.25 | 12000 | 0.89 | 0.65 | 0.22 | |
| 苦竹Pleioblastus amarus | 43 | 1.70 | 1.03 | 19136 | 0.19 | 0.53 | 0.03 | |
| 平车前Plantago depressa | 44 | 1.67 | 1.18 | 4444 | 0.07 | 0.52 | 0.16 | |
| 通泉草Mazus japonicus | 45 | 1.48 | 1.32 | 2222 | 0.22 | 0.39 | 0.28 | |
| 白茅Imperata cylindrica | 46 | 1.17 | 0.59 | 2222 | 0.01 | 0.15 | -0.53 | |
| 鸭跖草Commelina communis | 47 | 1.02 | 0.47 | 4444 | 0.03 | 0.02 | -0.76 | |
| 牛膝Achyranthes bidentata | 48 | 0.51 | 0.27 | 561 | 0.26 | -0.67 | -1.31 | |
| Ⅱ | 商陆Phytolacca acinosa | 1 | 512.40 | 1076.00 | 21632 | 24.59 | 6.24 | 6.98 |
| 小蓬草Conyza canadensis | 2 | 143.24 | 169.99 | 293333 | 3.69 | 4.96 | 5.14 | |
| 野胡萝卜Daucus carota | 3 | 126.44 | 83.72 | 174333 | 3.74 | 4.84 | 4.43 | |
| 飞蓬Erigeron acer | 4 | 84.26 | 57.75 | 613333 | 2.33 | 4.43 | 4.06 | |
| 早熟禾Poa annua | 5 | 79.52 | 58.87 | 116667 | 4.17 | 4.38 | 4.08 | |
| 水芹Oenanthe javanica | 6 | 48.27 | 21.00 | 13337 | 0.33 | 3.88 | 3.04 | |
| 狗尾草Setaria viridis | 7 | 42.44 | 73.50 | 68889 | 17.99 | 3.75 | 4.30 | |
| 大青Clerodendrum cyrtophyllum | 8 | 27.86 | 45.26 | 13333 | 0.33 | 3.33 | 3.81 | |
| 土荆芥Chenopodium ambrosioides | 9 | 24.13 | 14.89 | 1367 | 0.77 | 3.18 | 2.70 | |
| 艾Artemisia argyi | 10 | 17.87 | 14.04 | 53333 | 0.93 | 2.88 | 2.64 | |
| 龙葵 Solanum nigrum | 11 | 12.80 | 11.86 | 4977 | 1.06 | 2.55 | 2.47 | |
| 小巢菜Vicia hirsuta | 12 | 8.72 | 0.97 | 10112 | 3.21 | 2.17 | -0.03 | |
| 蛇莓Duchesnea indica | 13 | 4.62 | 1.54 | 13333 | 0.13 | 1.53 | 0.43 | |
| 物种生物量 B的标准差范围为:8%—16%, 锰吸收量U的标准差范围为:5%—18% | ||||||||
| 区域 Sites |
总植物种类数 Number of plant species |
总生物量∑Bi Total biomass/ (kg/hm2) |
总锰吸收量∑Ui Total manganese uptake/ (g/hm2) |
总密度∑d Total density/hm2 |
总盖度 Total coverage/% |
DB | DU | RB | 0 RU |
| Ⅰ | 48 | 23323.97 | 4280.20 | 1610166 | 285.61 | ||||
| Ⅱ | 13 | 1132.56 | 1599.37 | 1397981 | 63.27 | 22191 | 2518 | 20.6 | 2.55 |
| DB=∑BⅠ-∑BⅡ, DU=∑UⅠ-∑UⅡ, RB=∑BⅠ/∑BⅡ, RU=∑UⅠ/∑UⅡ | |||||||||
整体上, Ⅰ区的各项数据均显著高于Ⅱ区。Ⅰ区的生物总量、物种数、锰吸收总量、总密度、总盖度分别为Ⅱ区的20.6、3.7、2.6、1.2、4.5倍, Ⅰ与Ⅱ区的RB和RU比值也大于1, 分别为20.6和2.55。基于式8和式9, RB和RU比值为两区系统自由能的比值, 说明Ⅰ区生态系统的生物量和锰吸收量两项指标的Gibbs自由能均高于Ⅱ区, 差异非常显著 (P < 0.001)。表 3、4的数据说明使用有机菌肥不仅可以有效地促进试验区植物生长和锰的吸收, 还可以显著提高物种数量和植被覆盖率, 通过提高系统生产力和丰富生物多样性来改善试验区生态系统结构功能, 从而从整体上提高生态系统的自由能。
2.2 植物种类化学势差异基于植物干物质生物量B数据, 采用式12计算了Ⅰ和Ⅱ区物种的δB值。图 1展示了Ⅰ和Ⅱ区内植物种类间的化学势差异, 图中物种按δB值从大到小排列, 其植物种类编号与表 3编号相同。自图 1可清晰的看出, 整体上, Ⅰ区物种间化学势差异范围显著大于Ⅱ区 (P < 0.001)。Ⅰ和Ⅱ区δB值波动范围分别为-3.79—6.76和-2.17—2.54, 其中δB为负值为小于系统平均δB值的物种。Ⅰ区泡桐的δB值最高, 其次为栾树和构树, 说明泡桐、栾树和构树是改良Ⅰ区的优势植物种类。Ⅱ区δB值最高的物种是商陆, 其次为小蓬草和野胡萝卜, 说明这3种植物具有很强的适应锰污染土壤环境的能力。依据δB值大小排序可筛选出生物量大的耐性植物种类。

|
| 图 1 修复区 (Ⅰ) 和对照区 (Ⅱ) 植物种类间生物量化学势差异 Fig. 1 Difference in chemical potential (biomass) among plant species in restoration (Ⅰ) and control (Ⅱ) sites δB= ln (Bi)-(1/S)∑ln (Bi), Bi:生物量, S:物种数 |
图 2展示了Ⅰ和Ⅱ区内物种锰吸收量的化学势差异。与图 1相比, 物种锰吸收量化学势与生物量化学势差异小, δU与δB值变化趋势基本一致, Ⅰ区δU与δB的相关系数值R=0.994, Ⅱ区R=0.979, 说明植物锰吸收量与生物量呈显著正相关。整体上, Ⅰ和Ⅱ区内各自物种间的化学势差异显著, Ⅰ与Ⅱ区的δU值波动范围分别为-3.52—5.69和-3.42—3.59, Ⅰ区物种间化学势差异显著大于Ⅱ区 (P < 0.001)。与图 1物种排序相似, Ⅰ区δU值最高的物种也为泡桐, 其次为栾树和构树, Ⅱ区最高δU值的物种为商陆, 其次为小蓬草和野胡萝卜。基于δU值分析可筛选出锰吸收量大的植物种类。

|
| 图 2 修复区 (Ⅰ) 和对照区 (Ⅱ) 植物种类间锰吸收量化学势差异 Fig. 2 Difference in chemical potential (manganese) uptake among plant species in restoration (Ⅰ) and control (Ⅱ) sites δU= ln (Ui)-(1/S)∑ln (Ui), Ui:锰吸收量, S:物种数 |
表 5列出了Ⅰ和Ⅱ区基于δB、δU、锰平均浓度、TF值和优势度排序前10的植物种类, 前10物种δB值和 (∑δB)、δU值和 (∑δU)、以及按优势度排序前10物种的Simpson指数值。自表 5可看出, 由δB和δU值大小排序列出的前10植物种类基本一致。采用δB和δU值作为筛选修复植物的依据, 可满足快速恢复植被、增强系统去锰能力和提高污染地生产力的综合目标。Ⅰ区中泡桐、栾树和构树分别为用材林和能源树种, 具有较高的经济价值, 可作为锰污染区植被修复的建群树种。采用锰平均浓度、TF值、优势度排序列出的前10植物种类差异很大, 更为明显的是排前的主要是生物量小和锰吸收总量小的植物种类, 如Ⅰ区中的商陆、一年蓬、通泉草、狗牙根和Ⅱ区的小蓬草、狗尾草均属于一年生草本植物, 可利用的经济价值也很低。
| 区域 Sites |
名次 Sequence |
δB | δU | 锰浓度 Manganese content |
TF值 Transfer factor value |
优势度 Dominance |
∑δB | ∑δU | Simpson指数值break/>Simpson index value |
| Ⅰ | 1 | 泡桐 | 泡桐 | 商陆 | 大青 | 泡桐 | |||
| 2 | 栾树 | 栾树 | 一年蓬 | 商陆 | 狗牙根 | ||||
| 3 | 构树 | 构树 | 通泉草 | 悬钩子 | 栾树 | ||||
| 4 | 狗牙根 | 狗牙根 | 龙葵 | 蛇莓 | 灯心草 | ||||
| 5 | 芒 | 商陆 | 蒌蒿 | 小蓬草 | 小巢菜 | 25.88 | 27.93 | 0.81 | |
| 6 | 商陆 | 一年蓬 | 飞蓬 | 鸡眼草 | 酢酱草 | ||||
| 7 | 土荆芥 | 芒 | 悬钩子 | 芒 | 早熟禾 | ||||
| 8 | 灯心草 | 灯心草 | 大青 | 飞蓬 | 狗尾草 | ||||
| 9 | 一年蓬 | 早熟禾 | 野蔷薇 | 牛膝 | 小蓬草 | ||||
| 10 | 香丝草 | 悬钩子 | 早熟禾 | 蒌蒿 | 构树 | ||||
| Ⅱ | 1 | 商陆 | 商陆 | 商陆 | 商陆 | 商陆 | |||
| 2 | 小蓬草 | 小蓬草 | 狗尾草 | 土荆芥 | 飞蓬 | ||||
| 3 | 野胡萝卜 | 狗尾草 | 大青 | 小蓬草 | 狗尾草 | ||||
| 4 | 飞蓬 | 早熟禾 | 小蓬草 | 水芹 | 野胡萝卜 | ||||
| 5 | 早熟禾 | 飞蓬 | 龙葵 | 大青 | 小蓬草 | 4.87 | 7.27 | 0.73 | |
| 6 | 水芹 | 野胡萝卜 | 艾 | 狗尾草 | 早熟禾 | ||||
| 7 | 狗尾草 | 大青 | 早熟禾 | 小巢菜 | 艾 | ||||
| 8 | 大青 | 水芹 | 飞蓬 | 蛇莓 | 小巢菜 | ||||
| 9 | 土荆芥 | 土荆芥 | 野胡萝卜 | 飞蓬 | 水芹 | ||||
| 10 | 艾 | 艾 | 土荆芥 | 早熟禾 | 大青 |
自表 5可看出, Ⅰ区的∑δB、∑δU和Simpson指数值均大于Ⅱ区的值。Simpson指数是反映系统物种多样性指标, 说明∑δB和∑δU与Simpson指数值具有相似功能, 可作为系统生物多样性指标。总体上, Ⅱ区前10植物种类的生物量和吸收量虽然小于Ⅰ区, 但其能在未改良的高锰浓度污染环境中自然生长, 耐性很强, 因此可以作为锰污染土壤修复的先锋植物, 在建立污染区复合植物群落模式、提高植被覆盖率中可发挥重要的作用。
3 讨论试验区锰矿渣基质重金属含量高, 养分贫瘠, 只有少数重金属耐性强的植物能在这类恶劣土壤环境存活。而有机菌肥改良区存活的植物种类达48种, 为对照区的3.7倍, 其中有44种自然生草本植物和4种在高浓度重金属环境下难以存活的泡桐、栾树等木本植物, 说明专用有机菌肥的改良效果显著。专用肥含有抗性菌株、吸附介质、有机络合物和氧化还原酶等物质, 其除了能为植物提供必要的养分外, 最重要的功能是通过固定、吸附、络合、氧化和微生物吸收富集等作用来降低植物根际微环境中重金属的毒性, 缓解重金属对植物的直接危害。有关专用有机菌肥缓解重金属毒性的作用机理将在后续结合盆栽试验数据进行深入分析与讨论。
系统自由能G是植物生物量B、锰浓度、锰吸收量U、密度d、物种数S的函数, 包含了植物生长状况、重金属富集能力和生物多样性及其与立地环境的关系等综合信息, 因此可作为反映生态系统生产力、物种结构、丰富度和功能的综合指标。
甄别超富集植物的传统指标中, 植体重金属浓度主要体现植物的耐受性, TF值反映植物向地上部转移重金属元素的能力[12-13]。如García M A等用Pb浓度大小比较植物铅耐性[24], Juárez-Santillán L F等用TF值筛选锰耐性植物[25]。优势度是评价植物生长势的指标, 体现物种在植物群落中的扩繁能力[17]。商陆是噬锰植物, 耐受性和转运锰的能力都很强[26-27], 因此在对照区中, 商陆的5个指标排名都位居前列, 但在改良区中, 商陆排名靠前的指标仅为锰浓度和TF值, 主要原因是其生物量远远小于泡桐和栾树等其他树种, 锰吸收总量也相对小, 因此δB和δU值小。总体上, 在Ⅱ区5项指标排名靠前植物的δB和δU都远远低于Ⅰ区的值。
菌肥改良后Ⅰ区的植物种类数、总密度、总盖度显著增大。在生态系统中, 植物种类互相争夺土壤营养元素和生长空间, 盖度和密度大的物种则优势度相对高, 如盖度大的木本植物泡桐和栾树, 以及易存活密度大的狗牙根和灯心草。因此, 在修复植物筛选中, 仅知道物种个体的浓度C、转移系数TF的数据不够, 还需要知道给定区域单位面积上物种的生物量 (物种生产力), 密度 (物种扩展能力), 以及生态系统中物种间竞争、共生、抑制、互利等关系行为。采用植物重金属浓度、TF指标可以直接比较个体物种耐受和积累污染元素的生理生化特点, 但不能反应物种在重金属污染环境中的生态学特性。例如, 在给定面积的区域里, 自然萌发生长植物种类的锰浓度和TF值大, 而其密度、生物量和污染元素吸收量等值却小。与传统指标相比, 化学势差异指标δB和δU除了能体现植物生长和富集锰元素能力差异外, 还具有反映植物生态学特性的优势。
物种多样性是植被恢复过程中群落变化的重要指标[28], 多样性越高, 生态服务功能越高[29], 抗干扰能力越强[18], 反映物种多样性的指标很多, 常用的是Simpson指数[5, 30]。Simpson指数的变量因子是pi=ni/∑ni, 而化学势的变量因子是xi= Ci/∑Ci, 两者的差别是, Simpson指数以物种个体数ni与系统总个体数∑ni比值为基础来描述系统生物多样性, 而化学势是以物种生物量Ci与系统总生物量∑Ci的比值为基础来反映物种潜在的势能, 其中Ci为个体生物量ci与个体数ni的乘积 (Ci=cini), 因此, 化学势包含了Simpson指数变量因子, 系统∑δB和∑δU的绝对值大则说明系统生物多样性丰富。
试验数据分析结果表明, 采用自由能比和物种化学势差来评价锰矿污染区植被修复效果和筛选修复植物具有特殊的意义。已建立的自由能和化学势方程在描述生态系统能态, 系统及组分间能流, 结构功能, 生产力, 生物多样性和均匀度等方面具有较为广泛的应用价值。依据热力学原理, 系统自由能变ΔG=0是系统平衡的标志[15], 而化学势对温度的偏导数∂μ/∂T为偏摩尔熵的负值[16]。因此, ΔG值可以作为判断生态系统平衡的指标, 而∂μ/∂T方程可用以分析生态系统的熵与物种多样性的关系。这些方面的工作还有待今后进一步结合生态学理论开展深入的研究。
4 结论(1) 修复区植物种类数、生物量、锰吸收量和自由能远大于对照区的值, 说明使用有机菌肥可有效地促进试验区植物生长和锰的吸收, 丰富系统生物多样性, 改善试验区生态系统结构功能。
(2) 建立的生态系统Gibbs自由能方程可用以描述锰矿生态试验区植被系统的结构功能, 自由能比值不仅可用以检验污染区植物修复效果, 还可以作为比较生态系统立地环境差异的指标。
(3) 物种化学势反映物种适应和修复污染环境的能力, 与传统指标相比, 物种化学势能更好地反映植物的生态学特性, 可作为污染区修复植物筛选的重要指标。
| [1] | Jørgensen S E. Ecosystem services, sustainability and thermodynamic indicators. Ecological Complexity, 2010, 7(3): 311–313. DOI:10.1016/j.ecocom.2009.12.003 |
| [2] | Zhang J J, Gurkan Z, Jørgensen S E. Application of eco-exergy for assessment of ecosystem health and development of structurally dynamic models. Ecological Modelling, 2010, 221(4): 693–702. DOI:10.1016/j.ecolmodel.2009.10.017 |
| [3] | Perryman M E, Schramski J R. Evaluating the relationship between natural resource management and agriculture using embodied energy and eco-exergy analyses: A comparative study of nine countries. Ecological Complexity, 2015, 22: 152–161. DOI:10.1016/j.ecocom.2015.04.002 |
| [4] | Molozzi J, Salas F, Callisto M, Marques J C. Thermodynamic oriented ecological indicators: Application of eco-exergy and Specific eco-exergy in capturing environmental changes between disturbed and non-disturbed tropical reservoirs. Ecological Indicators, 2013, 24: 543–551. DOI:10.1016/j.ecolind.2012.08.002 |
| [5] | Lu H F, Wang Z H, Campbell D E, Ren H, Wang J. Emergy and eco-exergy evaluation of four forest restoration modes in southeast China. Ecological Engineering, 2011, 37(2): 277–285. DOI:10.1016/j.ecoleng.2010.11.003 |
| [6] | 唐得昊, 邹欣庆, 刘兴健. 海岸带生态系统健康评价中能质和生物多样性的差异——以江苏海岸带为例. 生态学报, 2013, 33(4): 1240–1250. |
| [7] | Jørgensen S E. Application of exergy and specific exergy as ecological indicators of coastal areas. Aquatic Ecosystem Health and Management, 2000, 3(3): 419–430. DOI:10.1016/S1463-4988(00)00034-8 |
| [8] | Tang D H, Zou X Q, Liu X J, Liu P T, Zhamangulova N, Xu X W H, Zhao Y F. Integrated ecosystem health assessment based on eco-exergy theory: A case study of the Jiangsu coastal area. Ecological Indicators, 2015, 48: 107–119. DOI:10.1016/j.ecolind.2014.07.027 |
| [9] | Draganovic V, Jørgensen S E, Boom R, Jonkers J, Riesen G, Van Der Goot A J. Sustainability assessment of salmonid feed using energy, classical exergy and eco-exergy analysis. Ecological Indicators, 2013, 34: 277–289. DOI:10.1016/j.ecolind.2013.05.017 |
| [10] | 李晴新, 冯剑丰, 朱琳. 生态能质 (eco-exergy) 在水生生态系统建模和评价中的应用. 生态学杂志, 2011, 30(2): 376–383. |
| [11] | Xu F, Yang Z F, Chen B, Zhao Y W. Ecosystem health assessment of the plant-dominated Baiyangdian Lake based on eco-exergy. Ecological Modelling, 2011, 222(1): 201–209. DOI:10.1016/j.ecolmodel.2010.09.027 |
| [12] | Tassi E L, Pedron F, Barbafieri M. Evaluating the absorption of boron by plants-a potential tool to remediate contaminated sediments from Cecina River Basin in Italy. Water, Air, & Soil Pollution, 2011, 216(1/4): 275–287. |
| [13] | Kumar N, Bauddh K, Kumar S, Dwivedi N, Singh D P, Barman S C. Accumulation of metals in weed species grown on the soil contaminated with industrial waste and their phytoremediation potential. Ecological Engineering, 2013, 61: 491–495. DOI:10.1016/j.ecoleng.2013.10.004 |
| [14] | 凌薇薇, 吴晓芙, 陈永华, 刘庆, 郭丹丹. 除锰微生物的筛选及其除锰特性研究. 中南林业科技大学学报, 2011, 31(6): 152–156. |
| [15] | Engel T, Reid P. Physical Chemistry (1st Edition). New York: Pearson Education Inc, 2006: 133. |
| [16] | 傅献彩, 沈文霞, 姚天扬. 物理化学 (第四版). 北京: 高等教育出版社, 1990: 160. |
| [17] | 李侠, 于明坚, 慎佳泓, 胡仁勇, 李铭红. 杭州湾滩涂Na元素含量对植物多样性和优势度的影响. 生态学报, 2007, 27(11): 4603–4611. DOI:10.3321/j.issn:1000-0933.2007.11.028 |
| [18] | 王芸, 欧阳志云, 郑华, 陈法霖, 陈圣宾, 曾静. 南方红壤区3种典型森林恢复方式对植物群落多样性的影响. 生态学报, 2013, 33(4): 1204–1211. |
| [19] | 贺梦璇, 李洪远, 莫训强, 孟伟庆, 杨佳楠. 基于响应面分析法土壤种子库植被恢复的因素优化. 应用生态学报, 2014, 25(8): 2311–2316. |
| [20] | Tang Y, Cao M, Fu X H. Soil seedbank in a dipterocarp rain forest in Xishuangbanna, Southwest China. Biotropica, 2006, 38(3): 328–333. DOI:10.1111/btp.2006.38.issue-3 |
| [21] | 马全林, 卢琦, 魏林源, 靳虎甲. 干旱荒漠白刺灌丛植被演替过程土壤种子库变化特征. 生态学报, 2015, 35(7): 2285–2294. |
| [22] | 程玲, 王亚珍, 许景伟, 翟树强, 曲木子, 李传荣. 灵山湾国家森林公园垂序商陆土壤种子库时空动态及其对防除措施的响应. 生态学报, 2015, 35(15): 5095–5103. |
| [23] | Lin L X, Cao M. Edge effects on soil seed banks and under-story vegetation in subtropical and tropical forests in Yunnan, SW China. Forest Ecology and Management, 2009, 257(4): 1344–1352. DOI:10.1016/j.foreco.2008.12.004 |
| [24] | García M Á, Alonso J, Melgar M J. Lead in edible mushrooms: Levels and bioaccumulation factors. Journal of Hazardous Materials, 2009, 167(1/3): 777–783. |
| [25] | Juárez-Santillán L F, Lucho-Constantino C A, Vázquez-Rodríguez G A, Cerón-Ubilla N M, Beltrán-Hernández R I. Manganese accumulation in plants of the mining zone of Hidalgo, Mexico. Bioresource Technology, 2010, 101(15): 5836–5841. DOI:10.1016/j.biortech.2010.03.020 |
| [26] | 梁文斌, 薛生国, 沈吉红, 王萍, 王钧. 锰胁迫对垂序商陆叶片形态结构及叶绿体超微结构的影响. 生态学报, 2011, 31(13): 3677–3683. |
| [27] | 徐向华, 李仁英, 刘翠英, 施积炎, 林佳. 超积累植物垂序商陆 (Phytolacca americana L.) 吸收锰机制的初步探讨. 环境科学, 2013, 34(11): 4460–4465. |
| [28] | Zhang J T, Dong Y R. Factors affecting species diversity of plant communities and the restoration process in the loess area of China. Ecological Engineering, 2010, 36(3): 345–350. DOI:10.1016/j.ecoleng.2009.04.001 |
| [29] | Rey Benayas J M, Newton A C, Diaz A, Bullock J M. Enhancement of biodiversity and ecosystem services by ecological restoration: a meta-analysis. Science, 2009, 325(5944): 1121-1124. |
| [30] | 沈亚强, 魏宏斌, 程旺大, 张红梅, 王为东, 尹澄清. 嘉兴市石臼漾水源生态湿地运行过程中植物多样性研究. 环境科学, 2011, 32(10): 2883–2890. |