 2017, Vol. 37
2017, Vol. 37文章信息
- 陈吉龙, 李国胜, 寥华军, 王炳亮, 崔林林
- CHEN Jilong, LI Guosheng, LIAO Huajun, WANG Bingliang, CUI Linlin.
- 辽河三角洲河口湿地典型芦苇群落最大光能转化率模拟
- Simulation of maximum light coversion efficiency for a Phragmites salt marsh in the Liaohe River estuarine wetland
- 生态学报. 2017, 37(7): 2263-2273
- Acta Ecologica Sinica. 2017, 37(7): 2263-2273
- http://dx.doi.org/10.5846/stxb201512162511
-
文章历史
- 收稿日期: 2015-12-16
- 网络出版日期: 2016-08-30





2. 中国科学院地理科学与资源研究所, 北京 100101;
3. 中国地质调查局滨海湿地生物地质重点实验室, 青岛 266071
2. Institute of Geographic Sciences and Natural Resources Research, Beijing 100101, China;
3. Key Laboratory of Coastal Wetland Biogeosciences, China Geologic Survey, Qingdao 266071, China
河口湿地是一类特别的湿地生态系统, 位于陆地、海洋和河流生态系统的交界处, 是生产力最高的生态系统之一, 可以与农田生态系统相媲美, 具有很高的固碳能力[1]。单位面积盐沼、红树林湿地甚至比成熟的热带雨林能封存高得多的碳, 在全球碳循环中扮演着重要角色。传统的样地观测是湿地生产力研究的基本方法, 站点实测数据精度高, 是模型校正和验证不可缺少的基础资料;但由于湿地特殊自然环境, 加大了野外测量的难度;另一方面, 样地观测在区域净初级生产力研究中存在明显局限性。在区域或全球尺度上, 模型估算成为净初级生产力一项重要手段[2]。其中, 以基于光能转化率的Carnegie Ames Stanford Approach (CASA) 模型应用较为广泛[3-4]。CASA模型能够与RS和GIS技术相结合, 为区域生产力的研究提供高效、精确的研究方法。
最大光能转化率是CASA模型最重要的参数之一, 它的大小直接影响到净初级生产力的总量。其含义为在理想情况下, 植被通过光合作用吸收单位光合有效辐射所固定的最大干物质总量。Raymond等认为在没有气候和其它因素限制情况下, 森林的最大光能转化率可达3.5 g C/MJ[5], 而在Harvard森林研究样地, 最大光能转化率约为0.55 g C/MJ[6]。农作物的最大光能转化率变化范围较窄, 介于1.1—1.4 g C/MJ之间, 而自然生态系统变化较大[7], 因此, Heimann等采用平均值1.25 g C/MJ估算了全球植被的年净初级生产力[8]。不同植被类型之间的最大光能转化率存在较大差异, 目前还无法直接测量, 只能通过模拟来求取。Potter提出CASA模型时, 利用Raich等[9]和McGuire等[10]观测的17组净初级生产力数据模拟出最大光能利率为0.389 g C/MJ[3-4]。虽然这个值被广泛用来估算区域植被净初级生产力[11], 但最大光能利率取值偏低导致净生产力被低估的现象也被大量报道[12-13], 这些结果表明对最大光能转化率的本地校正或者模拟对准确估算净初级生产力具有十分重要的作用。由于受实测资料的限制, 相关研究仅有少量的报道。Running等根据生态生理过程模型BIOME-BGC, 模拟出不同植被的最大光能转化率为0.389—1.259 g C/MJ[13]。朱文泉利用中国林业部的林业普查的实测数据和空间分辨率为8 km探路者数据, 采用CASA模型模拟出中国主要森林植被的最大光能转化率为0.542—0.985 g C/MJ[14]。由于陆地生态系统的复杂性, 对于不同植被类型, 最大光能转化率取值应该不同, 森林生态系统的模拟值不能简单应用到湿地生态系统;另外, 使用的数据及方法的差异也可能对结果有一定的影响。即便不考虑这些因素, 迄今为止, 仍未发现有关河口湿地最大光能转化率模拟研究的报道。
辽河三角洲河口湿地是我国重要的河口湿地之一, 是亚洲最大的暖温带滨海湿地, 位于辽宁省西南部辽河平原南端, 渤海辽东湾的顶部 (图 1)。受海洋和陆地交互、淡咸水交汇作用, 复杂动力机制造就了复杂多样的湿地类型和生态环境, 发育了以芦苇、翅碱蓬、獐毛、香蒲等作为建群种的湿地植物群落, 以及水稻田、虾蟹养殖池等人工湿地类型, 其中芦苇面积达到756 km2, 是亚洲第一大芦苇湿地, 已建成世界第二大苇场, 修建了完善的水利排灌体系, 改变了自然状态下河口湿地的水文规律。石油开发的各种地面工程造成了湿地景观破碎化, 严重破坏了湿地的原有生境。在这些人为干扰下, 辽河三角洲河口湿地的植被及净生产力格局发生明显的变化[15]。但由于种种原因, 我国对湿地的研究、监测开展较晚, 湿地生态系统缺少长期、系统的观测数据。尽管目前已经在辽河三角洲开展了一些净初级生产力的研究工作[16], 但绝大部分仍局限于有限样点的野外调查工作。RS和GIS技术为湿地净生产力的研究提供了高效手段, 但目前在本区域的应用也还停留在景观层面的研究上[17]。因此, 辽河三角洲河口湿地净初级生产力的遥感监测被列为中国地质调查项目及国土资源公益性行业科研专项的重要内容, 而准确确定芦苇群落的最大光能转化率是净初级生产力研究的前提工作, 对全面了解河口湿地碳汇功能、研究湿地碳循环具有十分重要的作用, 而且对准确评估我国芦苇湿地的固碳功能、发挥和提高湿地生态系统的固碳潜力具有重要的意义。

|
| 图 1 辽河三角洲河口湿地的位置及样点的分布 Fig. 1 Location of the sampling sites in Liaohe river estuarine wetland *背景为TM432波段合成 |
利用光能转化率原理, Monteith提出利用吸收光合有效辐射和光能转化率来估算陆地净初级生产力的概念, 并考虑了水分、温度和养分胁迫。在此基础上, Potter和Field建立了CASA模型, 实现了基于光能转化率原理的净初级生产力估算, 其理论构架可表示为下式[3-4]。
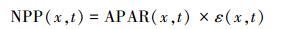
|
(1) |
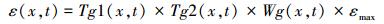
|
(2) |
式中, NPP为净初级生产力, APAR为吸收光合有效辐射, FPAR为光合有效辐射分量, t表示时间, x表示空间位置, εmax为最大光能转化率, Tg1和Tg2为温度胁迫系数, Wg为水分胁迫系数。
1.1.2 CASA模型参数化植被吸收的光合有效辐射取决于太阳总辐射和植被对光合有效辐射的吸收分量, 用下列公式表示:
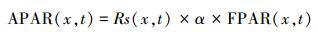
|
(3) |
式中, Rs(x, t) 是像元x处的太阳总辐射, α表示植被所能利用的太阳有效辐射占太阳总辐射的比例, 主要是波长范围在0.38—0.76 μm的可见光, α一般约占太阳总辐射能量的50%左右。由于太阳辐射站点数量少, 常常需要利用采用Ångström-Prescott (A-P) 模型和日照时数来计算[18]。
光合有效辐射分量是植被对光合有效辐射的吸收比例, 取决于植被类型和植被覆盖状况。研究表明光合有效辐射吸收分量与归一化植被指数 (NDVI) 存在线性关系[19]。这一关系可以根据某一植被类型NDVI的最大值和最小值以及所对应的光合有效辐射吸收分量最大值和最小值来确定:
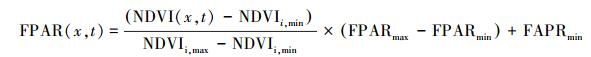
|
(4) |
式中, NDVIi, max和NDVIi, min分别对应第i种植被类型的NDVI的最大值和最小值。进一步的研究表明, 光合有效辐射吸收分量与比值植被指数 (RVI) 也存在较好的线性关系[20], 可由公式表示:
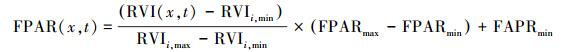
|
(5) |
式中, FPARmin和FPARmax的取值与植被类型无关, 分别为0. 001和0. 95。研究表明由NDVI所估算的光合有效辐射吸收分量比实测值高, 而由RVI所估算的光合有效辐射吸收分量则低于实测值, 但其误差小于由NDVI所估算的结果[21], 考虑到这种情况, Los将这两种方法结合起来, 取其平均值作为最终结果[22], 此时, 估算值与实测值之间的误差达到最小。
光能转化率是CASA模型中最关键的环节。Potter等认为在理想条件下植被具有最大光能转化率, 而在现实条件下的光能转化率受到环境因子如气温、水分状况以及大气水汽压差等因素的影响, 因此利用温度、大气及水分的可利用程度来调节。其公式可以表达为[3-4]:

|
(6) |
式中, Tg1(x, t) 和Tg2(x, t) 为温度胁迫系数, 分别用以下公式计算:
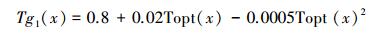
|
(7) |
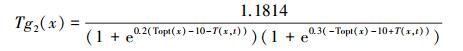
|
(8) |
式中, Topt为一年内NDVI达到最高时月份的平均气温, 认为此温度为植被生长的最适温度。当某一月平均温度小于或等于-10C时, Topt取0, 认为此时光合生产为零。若某月气温比最适宜温度高10℃或低13℃时, 该月的Tg2值等于月平均气温为最适宜温度Topt时的Tg2值的一半。
Wg (x, t) 为水分胁迫系数, 反映了水分条件对光能转化率的影响, 其取值范围为0.5至1, 由下式计算:
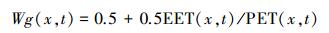
|
(9) |
式中, EET为实际蒸散, 根据FAO推荐的作物潜在蒸散的Penman-Monteith方程计算[23]。PET为潜在蒸散量, 根据Boucher提出的互补关系计算[24]。
根据公式 (1) 和 (2), 则最大光能转化率表示为:

|
(10) |
公式中可以看出, 当NPP、APAR、Tg1、Tg2和Wg为已知时, 便可以计算某一样点上植被的最大光能转化率。由于模型估算往往不可避免的要对某些过程进行简化;另一方面, 不同的数据误差也有可能被带入最后的结果中, 不同样点所模拟的最大光能转化率可能不同。因此, 本文将利用54个样点数据中的27个 (图 1), 根据误差最小原则, 拟合出芦苇的最大光能转化率。
1.1.3 净初级生产力分配部分净初级生产力在生长期分配到根部, 经过累积形成地下生物量。而利用CASA模型所估算的净初级生产力包括地上和地下部分。由于湿地的特殊困境, 加上芦苇发达的深根系, 对地下净初级生产力的测定比较困难。本文野外样地调查仅仅测定了芦苇地上部分。为了使CASA模型模拟的结果与实测生物量具有可比性, 采用根、茎、叶动态配置模式 (Pierre′s model), 根据可利用资源光 (L)、水分 (W) 和假营养 (N) 对不同器官进行碳的动态配置[25], 将地上部分的净初级生产力从CASA模型中所模拟的结果分离出来。
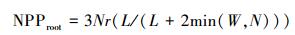
|
(11) |
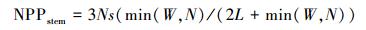
|
(12) |
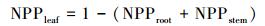
|
(13) |
式中, Nr和Ns代表没有资源限制时, 净初级生产力分配到根和茎中的比率, 默认取值0.3。资源可利用性因子L、W、N取值介于0.1(资源限制) 与1(资源充分并容易获取) 之间。L利用叶面积指数来估算, L=e-K×LAI, K为消光系数, 默认取值为0.5。水分因子W取决于土壤湿度和土壤质地。假营养因子N利用湿度和气温来计算, N=Fp×Ft, 其中Fp=PPT/PET, Ft=2[(T-30)/10], PPT为降水量, T为平均气温, PET潜在蒸散。
1.2 数据收集与处理基于CASA模型估算植被净初级生产力涉及大量的气象数据、遥感数据以及其它基础资料, 数据质量的好坏直接关系到结果的可靠性, 因此, 在数据的收集与处理过程中, 要尽可能的降低数据本身所包含的误差, 并使数据具有统一的标准。
1.2.1 气象数据气象数据包括2011年每日的总辐射、日照时数、平均气温、最高气温、最低气温、大气压、相对湿度。所有气象数据都从中国气象科学数据共享网 (http://cdc.cma.gov.cn/) 获取。根据CASA模型估算区域的净初级生产力, 需要基于网格点的气象数据, 这要求将气象站点的气象要素通过空间插值到区域, 但研究区域内仅有营口站的气象数据, 为了减少插值的边缘误差, 下载了整个辽宁省内的27个气象站的数据, 然后通过ANUCLIM插值方法内插出整个辽宁省网格点气象数据, 最后利用研究区域的边界剪切出区域内的数据[26]。
1.2.2 遥感数据遥感数据是进行区域净初级生产力研究不可缺少的数据源。由于MODIS多波段数据可以同时提供有关陆地、植被、大气、温度等特征信息, 其时间分辨率高, 重覆周期短, 是开展区域净初级生产力研究非常重要的资源。因此, 本文也使用MODIS数据产品来模拟芦苇群落的净初级生产力, 具体包括2011年全年16d合成的MODIS09反射率产品、2011年全年16d合成MOD13植被指数产品以及2011年全年16d合成MODIS15叶面积指数产品。所有MODIS数据均从NASA (http://reverb.echo.nasa.gov) 下载获取。这些数据产品都经过了一系列严格的质量控制和检验, 已经在国内外广泛使用, 实践证明这类产品的精确度能够满足此类研究工作的需要。
1.2.3 净初级生产力测量数据净初级生产力测量采用收割法, 选取连续分布的且长势均质的典型芦苇群落, 设置样地54个 (图 1), 其中27个用于模拟最大光能转化率, 另一半用于验证模拟结果。大样地面积设为250 m×250 m, 以匹配MODIS像元大小, 在每一样地的四角和中心设置5个2 m×2 m小样方, 将样方芦苇齐地收割, 按器官部位分别称总鲜重, 并各取样品一份 (300—500 g左右), 用于测定不同器官的干物质比率。并收集样方中所有调落物并测定鲜重, 同样也取样品一份, 测算调落物的干物质比率。将样品带回实验室在80℃下烘干48 h至恒重并称重, 然后根据不同器官的干物质比率及其总鲜重, 推算其样方内总干重, 以5个小样方的平均值计算样地单位面积生物量, 最后乘以碳转换系数 (0.45), 换算成单位面积上植被地上部分碳含量。各样点的调查具体情况见表 1。
| 模拟样点编号Sampling sites No. | 经度/(°) Longitude | 纬度/(°) Latitude | 植被覆盖度Vegetation coverage | 观测NPP/(kg C/m2) Observated NPP | 最大光能转化率/(g C/MJ) Maximum light conversion efficiency |
| 1 | 121.64584 | 40.91238 | 95% | 0.604 | 1.757 |
| 2 | 121.68842 | 40.94095 | 90% | 0.493 | 1.291 |
| 3 | 121.68153 | 40.96292 | 85% | 0.771 | 1.930 |
| 4 | 121.73628 | 40.95840 | 90% | 0.424 | 1.122 |
| 5 | 121.71923 | 41.00459 | 80% | 0.490 | 1.607 |
| 6 | 121.75887 | 41.00171 | 80% | 0.664 | 1.712 |
| 7 | 121.80473 | 41.02351 | 60% | 0.402 | 1.158 |
| 8 | 121.72478 | 41.02634 | 80% | 1.090 | 2.297 |
| 9 | 121.75261 | 41.04469 | 75% | 0.435 | 1.177 |
| 10 | 121.82411 | 41.04255 | 70% | 0.483 | 1.193 |
| 11 | 121.78935 | 41.07868 | 70% | 0.463 | 1.112 |
| 12 | 121.75301 | 41.11547 | 80% | 0.588 | 1.667 |
| 13 | 121.74062 | 41.16428 | 80% | 0.633 | 1.738 |
| 14 | 121.65253 | 41.23127 | 80% | 0.540 | 1.397 |
| 15 | 121.70103 | 41.26432 | 70% | 0.406 | 1.182 |
| 16 | 121.73387 | 41.20943 | 70% | 0.534 | 1.386 |
| 17 | 121.80019 | 41.20012 | 70% | 0.872 | 2.013 |
| 18 | 121.80817 | 41.15675 | 80% | 0.779 | 2.037 |
| 19 | 121.88129 | 40.91645 | 80% | 0.978 | 2.610 |
| 20 | 121.86287 | 40.93732 | 75% | 0.879 | 2.256 |
| 21 | 121.93085 | 40.94338 | 70% | 0.707 | 1.804 |
| 22 | 121.93343 | 40.96785 | 70% | 0.868 | 2.353 |
| 23 | 121.89491 | 40.96758 | 75% | 0.679 | 1.780 |
| 24 | 121.89799 | 40.98792 | 60% | 0.590 | 1.572 |
| 25 | 121.84760 | 41.20659 | 60% | 0.482 | 1.208 |
| 26 | 121.84058 | 41.25302 | 75% | 0.496 | 1.330 |
| 27 | 121.80091 | 41.24509 | 80% | 0.589 | 2.031 |
表 1列出了模拟的芦苇最大光能转化率, 样点模拟值为1.112—2.611 g C/MJ (1.656±0.43) (表 1), 模拟值平均值为1.667 g C/MJ。Potter提出CASA模型时, 植被最大光能转化率为0.389 g C/MJ[3-4], 明显低于本区域芦苇群落的最大光能转化率。在广东鼎湖山和黑石顶的观测也表明常绿阔叶林的实际光能转化率分别要大于0.605 g C/MJ和0.515 g C/MJ[21], 因此, 彭少麟采用最大光利用率为1.25 g C/MJ估算了广东省植被的净初级生产力[27], 但依然要低于芦苇群落的最大光能转化率。采用与本文相同的方法, 朱文泉利用净初级生产力的实测数据, 根据误差最小的原则模拟出中国主要森林植被类型的最大光能转化率为0.542—0.985 g C/MJ[21]。而Running根据生态生理过程模型BIOME-BGC模拟的不同的植被类型的最大光能转化率为0.389—1.259 g C/MJ[13], 其结果也都明显低于本区域芦苇群落。
利用模拟的最大光能转化率, 我们计算了芦苇群落的实际光能转化率, 样点模拟平均值为0.957—1.102 g C/MJ, 要高于我国植被的年平均光能转化率0.196—0.701 g C/MJ[14], 甚至要高于常绿阔叶林、常绿针叶林、热带常绿阔叶林等森林植被[27]。图 2为芦苇群落光能转化率的时间动态, 表现出明显的单峰曲线季节变化模式;从年初到4月上旬, 由于低温天气的限制, 光能转化率趋近于0;当温度升高到适宜植被生命活动时, 植被开始生长发育, 光能转化率开始上升;随着生长季的持续, 温度和降水的增加有利于叶片的光合作用, 光能转化率不断提高, 叶面积不断增加, 覆盖度逐渐增大, 在7月下旬本地区的温度和降水分别达到最大值时, 光能转化率也达到最高值1.524 g C/MJ;在植被长势达到最茂盛后, 随着温度和降水的减少, 加上大部分绿叶开始变黄或调落, 叶绿素含量降低, 光能转化率也开始降低;在生长季结束后, 低温寒冷天气限制了植被的生长, 光能转化率趋近于0。

|
| 图 2 芦苇群落光能转化率的时间动态 Fig. 2 Temporal vartion of light coversion efficiency of Phragmites salt marsh |
河口湿地位于陆地、海洋和河流生态系统的交界处, 是生产力最高的生态系统之一[1]。乔治亚海岸互花米草的净初级生产力达到7620 g C/m2[28], 我国长江口典型芦苇湿地净初级生产力也可达1100—2400 g C/m2[29]。这除了与河口湿地所处的特殊环境有关外, 还可能取决于湿地植被高的光能转化率。本文的结果表明芦苇群落具有很高的最大和实际光能转化率, 甚至要高于Running[13], 朱文泉[14]模拟的阔叶林、针叶林以及针阔混交林等森林植被, 这可能是由于植被的生长都需要充足的水分和营养条件, 而芦苇沼泽湿地恰恰处于陆地与水体的过渡地带, 它同时兼具丰富的陆生和水生动植物资源, 在长期的自然选择和进化过程中形成了其它任何生态系统都无法比拟的基因库;别一方面, 沼泽湿地的泥碳土壤经过长期累积, 为沼泽植被的生长提供了丰富的营养元素;加上河口湿地海洋与河流的水文交互作用有利于植被光合作用对碳的转化, 已有的研究也表明河口湿地高的生产力与水文的季节性波动有着密切的相关性[30]。这些特殊的水文和土壤条件为河口湿地芦苇群落对碳的转化提供了有利的条件, 形成了芦苇群落高的光能转化率。
3.2 模拟结果验证对模拟结果进行验证是进一步分析和应用的基础, 由于在目前还不能通过试验和设备来测量植被的最大光能转化率, 无法通过实测值来检验模拟的效果。最大光能转化率真是CASA模型的重要参数, 因此, 利用54个野外测量数据中的另外27个, 并利用模拟的芦苇最大光能转化率来计算对应样点的地上净初级生产力, 通过验证地上净初级生产力来间接验证模拟的最大光能转化率的可靠性。图 3为CASA模型模拟的芦苇地上净初级生产力与实测值散点图。总体上看, CASA模型模拟结果与观测值具体较好的一致性。模拟值与实测值之间的相关系数为0.63(P < 0.001), 达到极显著性水平。最大相对均方根误差 (RRMSE) 为40%, 平均相对均方根误差为24%。27个验证点的平均模拟值和观测值较为接近, 分别为619.9 g C/m2和627.3 g C/m2, 两者仅相差7.5 g C/m2。这间接的表明本文所模拟的芦苇最大光能转化率具有较高的可靠性。而当最大光能转化率采用0.389 g C/MJ时, 模拟的地上净初级生产力为112.87—184.59 g C/MJ (146.95±13.96) (图 3), 明显低估了本区域芦苇群落的固碳能力。中国芦苇资源丰富, 分布集中而广泛, 全国范围内有l4个主要芦苇产区, 芦苇湿地面积达130万hm2[31]。按照本文的最大光能转化率计算, 则芦苇的年固碳能力可达到10.04—15.14 Tg C (13.55±1.25);而最大光能转化率采用0.389 g C/MJ时, 模拟的芦苇年净初级生产力仅为2.34—3.53 Tg C (3.16±0.29), 则芦苇湿地的固碳能力将会被低估75%。

|
| 图 3 地上净初级生产力模拟值与观测值 Fig. 3 Scatter plots of aboveground NPP estimation vs. observation *原始结果是指采用最大光能转化率0.389 g C/MJ所计算的NPP, 模拟结果是指采用1.667g C/MJ所计算的NPP |
利用CASA模型来模拟植被的最大光能转化率, 需要先利用遥感数据和气象数据计算APAR, FPAR, Wg和Tg, 因此, 这些数据误差也会被带入最后的模拟结果中。APAR的数据误差主要来源于利用日照时数对太阳总辐射的估算, FPAR的误差则主要来源于NDVI, Wg和Tg则主要取决于气温和降水。将这些主要的影响因素以2011年的实际值为基准, 在±40%的变化范围内, 以5%的间距变化, 采用单因素分析误差对模拟结果的影响。由于植被的NDVI的值介于0—1之间, 当变化超过30%时, 出现NDVI大于1的情况, 因此NDVI的变化范围设为±30%。
图 4结果为不同影响因素值域下模拟的芦苇最大光能转化率和最大光能转化率对其影响因素的一阶导数。从图中可以看出, 不同的因素对模拟结果的影响不同。当总辐射和NDVI在±40%的范围内变化时, 模拟的芦苇最大光能转化率分别为1.239—2.568 g C/MJ (1.752±0.409) 和1.313—2.276 g C/MJ (1.716±0.308), 模拟值随着总辐射和NDVI的增加而呈现下降的趋势 (图 4), 并表现出较强的敏感性, 其一阶导数分别为-1.351—-0.642(-0.917±0.23) 和-1.218—-0.708(-0.924±0.17)。在低值区域, 这种敏感性表现的更加明显, 并虽着总辐射和NDVI的增加而呈现下降的趋势 (图 4)。尽管如此, 模拟值仍然落在样点的变化范围之内 (1.112—2.611 g C/MJ)。本文的结果表明在高纬度及总辐射较低的地区, 控制总辐射的精度显得格外重要, 因为利用CASA模型模拟的植被最大光能转化率可能具有更大不确定性。但由于观测设备和观测方法的限制, 我国太阳辐射观测站稀少, 常需要利用日照时数来估算。在本研究区域也没有太阳总辐射观测站, 只能利用日照时数来计算[26]。即便是整个辽宁省也仅有朝阳、大连和沈阳具有辐射观测数据, 因此, 利用这3个点的数据, 采用交叉验证的方法来检验所估算的太阳总辐射的精度。检验结果显示估算的太阳总辐射具有很高的精度 (图 5), 3个样点的相对均方根误差均小于7%, 总体精度达到95%, 按±5%的误差估计, 则模拟的芦苇最大光能转化率介于1.598—1.743 g C/MJ之间, 绝对变化幅度为-0.069—0.076 g C/MJ, 相对变化幅度仅为-4.14%—4.56%。这表明本文模拟的结果具有一定的稳定性和可靠性。

|
| 图 4 模拟的最大光能转化率对主要因素的敏感性 Fig. 4 Sensibility of the simulated maximum light conversion efficiency to affect factors |

|
| 图 5 太阳总辐射的观测值与预测值 Fig. 5 Scatter plots of global solar radiation estimation vs. observation RRMSE:相对均方根误差Relaitve root mean square error |
模拟的芦苇最大光能转化率对NDVI也具有较强的敏感性, NDVI±30%的变化引起的模拟结果绝对变化幅度为-0.35—0.609 g C/MJ, 相对变化幅度为-21.2%—36.5%, 这也证明空间分辨率和植被覆盖度对模拟的结果有重要影响。随着植被覆盖度的增加, 这种敏感性有降低的趋势, 如图 4所示, 一阶导数随着NDVI的增加而呈上升 (负值) 的趋势。NDVI的不确定性是个十分复杂的问题, 有来自不同的遥感平台、传感器、遥感方式的差异的影响, 即便是不考虑这些因素, 云、大气成分、地面背景、观测角度等因素也都对NDVI有着重要的影响。刘良明等指出MODIS NDVI的不确定性约为12%, 随着NDVI值的增加, 这种不确定性还会减少[32];而Trishchenko等的研究表明MODIS NDVI数据的误差高达30%[33]。以最大误差30%考虑, 则模拟结果为1.313—2.276 g C/MJ, 仍然比较集中在样点的变化范围之内 (1.112—2.611 g C/MJ)。别一方面, 由于MODIS NDVI被广泛作为区域植被及净初级生产力的研究的首选数据源。相同的数据源使得这些因素引起的NDVI的不确定性具有相似性, 使最终结果之间差异将会大大减小, 结果具有可比性。即便是使用不同尺度或者分辨率的MODIS NDVI数据 (图 6), 其结果也十分相近。从图 6中可以看出, 250 m和1000 m分辨率的MODIS NDVI几乎重合, 差异约为1%, 模拟值变化仅仅为0.016 g C/MJ。这是因为MODIS NDVI的合成都是利用分辨率为250 m的第一波段和第二波段反射率为基础, 优先选择近星下点无云像元, 尽可能减小云、暗影、大气气溶胶的影响、应用BRDF模式将角度订正到一致的标准化的视角和太阳大顶角的条件下, 保证合成资料的质量和一致性[34]。因此, 本文基于250 m分辨率的MODIS NDVI数据的结果仍然可以应用到1000 m分辨率尺度下。

|
| 图 6 两种分辨率的芦苇MODIS NDVI Fig. 6 Temporal vartion of MODIS NDVI with the resolution of 250m and 1000m MODIS NDVI:中分辨率成像光谱仪植被指数Moderate Resolution Imaging Spectroradiometer Normalized Difference Vegetation Index |
模拟的最大光能转化率随着气温的增加呈上升的趋势, 但随着降水量的增加呈下降的趋势, 两者的一阶导数都较小, 并且保持比较恒定的值 (图 4), 表明模拟结果对温度和降水量的敏感性较小而且比较稳定。模拟值分别为1.547—1.801 g C/MJ (1.669±0.08) 和1.586—1.774 g C/MJ (1.672±0.06) 之间, 绝对变化幅度分别为-0.12—0.134 g C/MJ和-0.08—0.107 g C/MJ。而实际上, 温度和降水的观测简单而且精度高, 其误差远远要小于50%。因此, 温度和降水误差对模结果的影响更加微弱。
4 结论准确确定最大光能转化率是利用光能利用率模型研究区域植被净初级生产力的基础。本文利用遥感技术结合地面气象观测和生产力野外监测, 利用CASA模型反演了辽河三角洲河口湿地典型芦苇群落的最大光能转化率。芦苇群落具有很高的碳转化能力, 最大光能转化率达到1.667 g C/MJ, 实际光能转化率达到0.957—1.102 g C/MJ。模拟最大光能转化率对总辐射和NDVI表现出较强的敏感性, 在低值区域, 这种敏感性表现的更加明显;因此, 在总辐射较低的地区, 控制总辐射的精度显得格外重要。本文总辐射模拟精度达到95%, 按±5%的误差估计, 则模拟值相对变化幅度仅为-4.14%—4.56%, 具有一定的稳定性和可靠性。模拟结果对NDVI的敏感性随着NDVI的增加而降低, 即便是以误差30%考虑, 则模拟值仍然比较集中在样点的变化范围之内。而在实际应用中, 由于MODIS NDVI被广泛用于区域净初级生产力的研究, 不同分辨率的MODIS NDVI差异较小, 模拟结果也更加稳定。模拟芦苇最大光能转化率对温度和降水量的敏感性较小而且比较稳定, 因此, 温度和降水误差对模拟结果影响较小。
| [1] | Odum W E, Odum E P, Odum H T. Nature's pulsing paradigm. Estuaries, 1995, 18(4): 547–555. DOI:10.2307/1352375 |
| [2] | Parton W J, Scurlock J M O, Ojima D S, Gilmanov T G, Scholes R J, Schimel D S, Kirchner T, Menaut J C, Seastedt M T, Moya E G, Kamnalrut A, Kinyamario J I. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide. Global Biogeochemical Cycles, 1993, 7(4): 785–890. DOI:10.1029/93GB02042 |
| [3] | Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production:a process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811–841. DOI:10.1029/93GB02725 |
| [4] | Field C B, Randerson J T, Malmstr m C M. Global net primary production:combining ecology and remote sensing. Remote Sensing of Environment, 1995, 51(1): 74–88. DOI:10.1016/0034-4257(94)00066-V |
| [5] | Raymond E, Hunt J R. Relationship between woody biomass and PAR conversion efficiency for estimating net primary production from NDVI. International Journal of Remote Sensing, 1994, 15(8): 1725–1729. DOI:10.1080/01431169408954203 |
| [6] | Wofsy S C, Goulden M L, Munger J W, Fan S M, Bakwin P S, Daube B C, Bassow S L, Bazzaz F A. Net exchange of CO2 in a mid-latitude forest. Science, 1993, 260(5112): 1314–1317. DOI:10.1126/science.260.5112.1314 |
| [7] | Russell G, Jarvis P G, Monteith J L. Absorption of radiation by canopies and stand growth//Russell G, Marshall B, Jarvis P G, eds. Plant Canopies:Their Growth, Form and Function. Cambridge:Cambridge University Press, 1989:21-39. |
| [8] | Heimann M, Keeling C. A three-dimensional model of atmospheric CO2 transport based on observed winds:2. model description and simulated tracer experiments. American Geophysical Union, 1989, 55: 237–275. |
| [9] | Raich J W, Rastetter E B, Melillo J M, Kicklighter D W, Steudler P A, Peterson B J, Grace A L, Moore Ⅲ B, Vörösmarty C J. Potential net primary productivity in South America:application of a global model. Ecological Applications, 1991, 1(4): 399–429. DOI:10.2307/1941899 |
| [10] | McGuire A D, Melillo J M, Joyce L A, Kicklighter D W, Grace A L, Moore Ⅲ B, Vorosmarty C J. Interactions between carbon and nitrogen dynamics in estimating net primary productivity for potential vegetation in North America. Global Biogeochemical Cycles, 1992, 6(2): 101–124. DOI:10.1029/92GB00219 |
| [11] | Lobell D B, Hicke J A, Asner G P, Field C B, Tucker C J, Los S O. Satellite estimates of productivity and light use efficiency in United States agriculture, 1982-98. Global Change Biology, 2002, 8(8): 722–735. DOI:10.1046/j.1365-2486.2002.00503.x |
| [12] | Goetz S J, Prince S D. Remote sensing of net primary production in boreal forest stands. Agricultural and Forest Meteorology, 1996, 78(3/4): 149–179. |
| [13] | Running S W, Thornton P E, Nemani R, Glassy J M. Global terrestrial gross and net primary productivity from the earth observing system//Sala O E, Jackson R B, Mooney H A, Howarth R W, eds. Methods in Ecosystem Science. New York:Springer-Verlag, 2000:44-57. |
| [14] | 朱文泉, 潘耀忠, 何浩, 于德永, 扈海波. 中国典型植被最大光利用率模拟. 科学通报, 2006, 51(6): 700–706. |
| [15] | 王宪礼, 胡远满, 布仁仓. 辽河三角洲湿地的景观变化分析. 地理科学, 1996, 16(3): 260–265. |
| [16] | 汲玉河, 周广胜. 辽河三角洲地区植被的地上净初级生产力格局及其制约因子. 生态环境学报, 2010, 19(10): 2269–2274. DOI:10.3969/j.issn.1674-5906.2010.10.001 |
| [17] | 刘红玉, 吕宪国, 刘振乾, 杨青, 张柏. 辽河三角洲湿地资源与区域持续发展. 地理科学, 2000, 20(6): 545–551. |
| [18] | Chen J L, Li G S, Xiao B B, Wen Z F, Lv M Q, Chen C D, Jiang Y, Wang X X, Wu S J. Assessing the transferability of support vector machine model for estimation of global solar radiation from air temperature. Energy Conversion and Management, 2015, 89: 318–329. DOI:10.1016/j.enconman.2014.10.004 |
| [19] | Roujean J L, Breon F M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote sensing of Environment, 1995, 51(3): 375–384. DOI:10.1016/0034-4257(94)00114-3 |
| [20] | Sellers P J, Berry J A, Collatz G J, Field C B, Hall F G. Canopy reflectance, photosynthesis, and transpiration. Ⅲ. A reanalysis using improved leaf models and a new canopy integration scheme. Remote Sensing of Environment, 1992, 42(3): 187–216. DOI:10.1016/0034-4257(92)90102-P |
| [21] | 彭少麟, 张祝平. 鼎湖山地带性植被生物量、生产力和光能利用效率. 中国科学:B辑, 1994, 24(5): 497–502. |
| [22] | Los S O. Linkages between global vegetation and climate:an analysis based on NOAA advanced very high resolution radiometer data[D]. Washington:National Aeronautics and Space Administration (NASA), 1998. |
| [23] | Allen R G, Pereira L S, Raes D. Crop evapotranspiration:guidelines for computing crop water requirements. FAO irrigation and drainage paper 56. Rome:FAO, 1998. |
| [24] | 张新时. 植被的PE (可能蒸散) 指标与植被-气候分类 (二)——几种主要方法与PEP程序介绍. 植物生态学报, 1989, 13(3): 197–207. |
| [25] | Friedlingstein P, Joel G, Field C B, Fung I Y. Toward an allocation scheme for global terrestrial carbon models. Global Change Biology, 1999, 5(7): 755–770. DOI:10.1046/j.1365-2486.1999.00269.x |
| [26] | Chen J L, Li G S. Parameterization and mapping of solar radiation in data sparse regions. Asia-Pacific Journal of Atmospheric Sciences, 2012, 48(4): 423–431. DOI:10.1007/s13143-012-0038-y |
| [27] | 彭少麟, 郭志华, 王伯荪. 利用GIS和RS估算广东植被光利用率. 生态学报, 2000, 20(6): 903–909. |
| [28] | Schubauer J P, Hopkinson C S. Above-and belowground emergent macrophyte production and turnover in a coastal marsh ecosystem, Georgia. Limnology and Oceanography, 1984, 29(5): 1052–1065. DOI:10.4319/lo.1984.29.5.1052 |
| [29] | 梅雪英, 张修峰. 长江口典型湿地植被储碳、固碳功能研究-以崇明东滩芦苇带为例. 中国生态农业学报, 2008, 16(2): 269–272. |
| [30] | Ahearn D S, Viers J H, Mount J F, Dahlgren R A. Priming the productivity pump:flood pulse driven trends in suspended algal biomass distribution across a restored floodplain. Freshwater Biology, 2006, 51(8): 1417–1433. DOI:10.1111/fwb.2006.51.issue-8 |
| [31] | 闫明, 潘根兴, 李恋卿, 邹建文. 中国芦苇湿地生态系统固碳潜力探讨. 中国农学通报, 2010, 26(18): 320–323. |
| [32] | 刘良明, 梁益同, 马慧云, 黄靖. MODIS和AVHRR植被指数关系的研究. 武汉大学学报:信息科学版, 2004, 29(4): 307–310. |
| [33] | Trishchenko A P, Cihlar J, Li Z Q. Effects of spectral response function on surface reflectance and NDVI measured with moderate resolution satellite sensors. Remote Sensing of Environment, 2002, 81(1): 1–18. DOI:10.1016/S0034-4257(01)00328-5 |
| [34] | Huete A, Justice C, Van Leeuwen W. MODIS vegetation index (MOD 13). Version 3. Algorithm theoretical basis document. ATBD-MOD-13, NASA, 1999. |