 2017, Vol. 37
2017, Vol. 37文章信息
- 黄兴召, 许崇华, 徐俊, 陶晓, 徐小牛
- HUANG Xingzhao, XU Chonghua, XU Jun, TAO Xiao, XU Xiaoniu.
- 利用结构方程解析杉木林生产力与环境因子及林分因子的关系
- Structural equation model analysis of the relationship between environmental and stand factors and net primary productivity in Cunninghamia lanceolata forests
- 生态学报. 2017, 37(7): 2274-2281
- Acta Ecologica Sinica. 2017, 37(7): 2274-2281
- http://dx.doi.org/10.5846/stxb201512132482
-
文章历史
- 收稿日期: 2015-12-13
- 网络出版日期: 2016-08-30





2. 大别山区农林特色产业协同创新中心, 合肥 230036
2. Collaborative Innovation Center of Agri-forestry Industry in Dabieshan Area, Hefei 230036, China
森林生产力是森林生态系统中植物群落在自然环境条件下的生产能力, 是森林生态系统研究的核心内容之一[1-2]。森林生产力数据作为森林生态系统养分循环和能量流动研究的基础, 它不仅显示自身群落结构的基本特征, 也是森林与环境关联的具体体现[3-4]。探讨森林生产力与环境因子及林分结构之间的关系, 分析立地环境和林分因子对森林生产力的影响, 对了解森林生产力对环境因子和林分因子变化的响应具有重要意义, 为森林科学管理和利用提供依据[5-6]。
森林生产力的高低反映了环境因子与林分因子的综合信息[7]。环境因子与森林所在的经纬度位置、海拔和立地条件等因素关联[8]。在这些环境因子中, 温度和降水作为森林生产力形成和发展的必要环境因子, 是森林生产力时空形态变化的主要影响因子[9]。许多文献探讨了不同地区森林生产力与气温、降水之间的关联性, 并且建立了不同森林类型的预估模型[8, 10-11]。同时在估算森林生产力方法中, 有效利用GIS (地理信息系统) 技术, 与环境因子、森林生理生态过程和立地条件等有效结合, 来阐述森林生产力的影响机制[12-15]。林分特征因子中密度和林龄是影响生产力的主要因素, 生产力随林分密度和林龄的变化而变化, 且林分密度和林龄紧密相关[16]。目前对于森林生产力的研究中, 大多数阐述环境因子与生产力的关系, 也有文献分析了林分因子与生产力的关系, 但尚未报道环境因子和林分因子与生产力的关系。
杉木 (Cunninghamia lanceolata) 是我国南方重要用材树种。根据第八次全国森林资源调查, 杉木天然林面积居第九位, 人工林面积居首位[17]。20世纪70年代至今, 很多学者对杉木林生物量和生产力进行了大量的研究和报道, 由于样本的测定费事费力, 因此, 利用这些研究结果, 分析杉木人工林生产力与环境因子或者林分因子之间的关系成为当前研究的热点之一。Luo等[18]收集了511篇 (其中杉木林83篇) 不同森林类型根冠比数据, 分析不同森林类型根冠比与温度和降雨量的变化;侯振宏等[19]收集了78篇杉木人工林生物量和生产力的数据, 分析了区域尺度杉木人工林生物量和生产力与林龄的关系;Reich等[20]通过收集全球6200种森林类型生物量, 研究了生物量与温度的关系;左舒翟等[21]收集94篇杉木林生物量估算参数数据, 分析了生物量估算参数与温度和降雨的关系;Poorter等[22]收集了1200个树种的生物量数据, 分析生物量在树种之间的差异。
以上的研究主要使用线性模型分析杉木林生物量与温度和降雨、或者和林龄的关系, 分析各因子对生产力直接影响作用, 忽略因子之间的间接响应, 特别是杉木林生产力与环境因子和林分特征因子的综合关联特征的研究未见有报道。结构方程模型是一种建立、估计和检验因果关系模型的方法。模型中既包含有可观测的显在变量, 也可能包含无法直接观测的潜在变量。结构方程模型可以替代多重回归、通径分析、因子分析、协方差分析等方法, 清晰分析多个因子对总体的作用和因子之间的相互关系[23]。本研究通过收集杉木林生产力数据, 利用结构方程建立杉木林生产力与温度、降雨、林龄和林分密度的关系, 阐述环境因子和林分因子对生产力的影响, 以期为杉木林可持续经营和长期生产力维持提供理论依据。
1 材料和方法 1.1 数据收集与整理通过中国知网数据库、维普中文期刊数据库、Science direct等数据库, 搜索杉木林生产力相关的文献, 建立杉木林生产力实测数据集。数据集包括试验样地经纬度、年均降水量 (MAP)、年均温度 (ALT)、林龄 (Age)、平均胸径 (DBH)、平均树高 (H)、林分密度、净初级生产力 (NPP) 等信息。在初步收集的基础上进行筛选。首先, 删除林龄小于3a的杉木天然林或者人工林数据;其次, 杉木林生产力的测定均为标准木解析计算得到, 如果数据的研究样地在同一个地点, 且林龄和平均密度相同的文献数据, 通过求生产力均值作为一行数据;最后, 对于杉木林生产力的异常值通过咨询专家进行专业判断进行取舍。本文最终搜集筛选出155篇644条杉木林生产力数据。数据集样地的分布如图 1所示。包含了安徽、江苏、浙江、江西、福建、湖北、湖南、广西、广东、贵州和四川等11个省份, 其中福建、湖南、江西和广西为杉木样地较多的省份。

|
| 图 1 数据集的样地空间分布图 Fig. 1 Spatial distribution of sampling sites used in data collection |
本文杉木林生产力为杉木林净初级生产力, 即单位时间和单位面积上, 杉木通过光合作用产生的有机物中扣除自养呼吸后的剩余部分, 其计量单位为t hm-2 a-1或者g m-2 a-1。收集数据中少量缺失年均温度和年均降水量的数据, 通过中国气象科学数据共享网 (http://cdc.cma.gov.cn/home.do), 利用缺失数据的经纬度坐标, 获取该样地的年均降水量和年均温度。
1.2 数据分析利用结构方程模型 (Structural Equation Model, SEM), 解析杉木林净初级生产力与年均降水量、年均温度、林龄和林分密度之间的关系。结构方程模型是基于变量的相关系数或者协方差矩阵来分析变量之间关系的一种统计方法。该方法假定一组隐变量之间存在因果关系, 隐变量通常是某几个显变量中的线性组合[24-25]。通过验证显变量之间的相关系数或者协方差, 可以估计出结构模型中的通径系数, 从而在统计上检验所假设的模型对所研究的过程是否合适, 如果证实所假设的模型合适, 就可以说假设隐变量之间的关系是合理的。
结构方程模型的一般表达式为:

|
(1) |
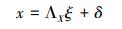
|
(2) |
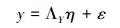
|
(3) |
公式 (1) 为结构方程模型中的结构模型, 表示隐变量与显变量之间的关系。其中, η是隐变量向量, β是隐变量η的系数矩阵, 也是隐变量间的通径系数矩阵;ξ是显变量向量;Г是显变量ξ的系数矩阵, 也是显变量对相应内生潜变量的通径系数矩阵;ζ为残差向量。
公式 (2) 和 (3) 为结构方程模型中的测量模型。测量模型一般由两个方程组成, 分别表示显变量向量ξ和观测变量x之间, 以及隐变量向量η和内生的观测变量y之间的联系。式中, ΛX为观测变量x在显变量向量ξ上的因子载荷矩阵, δ为公式 (2) 的误差向量;ΛY为观测变量y在隐变量向量η上的因子载荷矩阵, ε为公式 (3) 的误差向量。
数据收集的杉木林净生产力的计量单位为t m-2 a-1或g m-2 a-1, 本文统一使用g m-2 a-1为单位。利用结构方程模型时, 对年均温度、年均降水量、林龄、林分密度和净生产力统一进行自然对数转化后进行分析和建模[26]。在分析杉木林净初级生产力与年均降水量、年均温度、林龄和林分密度之间相关系数的基础上, 应用R中的lavaan包建立结构方程模型, 阐述杉木林净初级生产力与年均温度、年均降水量、林龄和林分密度之间的关系。数据统计分析和绘图使用R和Excel软件[27]。
2 结果与分析 2.1 杉木林净生产力与环境因子的分布特征根据收集整理的数据集, 绘制杉木林净初级生产力与之相匹配的环境因子的散点分布图和频度分布图 (图 2), 环境因子包含年均降水量和年均温度两个因子。从散点分布图中 (图 2) 可以看出净初级生产力与年均降雨量无明显的变化趋势。在年均温度小于23 ℃时, 净初级生产力随年均温的升高有上升的趋势。频度分布图用以检验杉木林净初级生产力样地的年均降雨量和年均温度的分布特征, 其分布如图 2所示, 杉木林净初级生产力样地在年均降雨量和年均温度下呈现明显的正态分布。杉木林净初级生产力样地分别集中在年均降雨量1400—1900 mm之间、年均温度16—20 ℃之间。

|
| 图 2 杉木林净初级生产力与年均降水量和年均温度的分布 Fig. 2 Distribution of NPP responding to MAP and MAT for Cunninghamia lanceolata forests 散点图为杉木林净初级生产力与年均降水量和年均温度的分布情况, 频度图杉木林净初级生产力分别在年均降水量和年均温度的频度分布情况 |
征绘制杉木林净初级生产力与林分因子的散点分布图和频度分布图 (图 3), 林分因子包含林龄和林分密度两个因子。从散点分布图中可以看出净初级生产力随着林龄的增长有下降趋势, 与林分密度有先上升后下降的趋势。同时, 检验杉木林净初级生产力样地的林龄、林分密度的分布特征, 其分布如图 3所示, 杉木林净初级生产力样地分别集中在林龄5—30 a之间、林分密度1500—3000株/hm2之间。杉木林净初级生产力样地在林分密度下呈现明显的正态分布, 但在林龄因子下的正态分布特征不明显。

|
| 图 3 杉木林净初级生产力与林龄和林分密度的分布 Fig. 3 Distribution of NPP responding to Age and Density for Cunninghamia lanceolata forests 散点图为杉木林净初级生产力与林龄和林分密度的散点分布情况, 频度图杉木林净初级生产力分别在林龄和林分密度的频度分布情况 |
对年均温度、年均降水量、林龄、林分密度和净生产力统一进行自然对数转化, 对其进行相关分析。结果如表 1所示, 杉木林净生产力与年均降水量和年均温度呈显著正相关, 相关系数分别是0.378和0.630;与林龄和林分密度呈显著负相关, 相关系数分别是-0.332和-0.406。林分密度和年均温度呈显著正相关, 相关系数是0.489;与林龄呈显著负相关, 相关系数是-0.467。林龄与年均温度、年均降水量相关性均不显著。
| 分析因子Factor | 年均降雨量MAP | 年均温度MAT | 林龄Age | 密度Density | 生产力NPP |
| 年均降雨量MAP | 1.000 | 0.576** | 0.024 | -0.168 | 0.630** |
| 年均温度MAT | 1.000 | 0.288 | 0.498** | 0.378* | |
| 林龄Age | 1.000 | -0.467** | -0.332* | ||
| 密度Density | 1.000 | -0.406** | |||
| 生产力NPP | 1.000 | ||||
| *相关显著 (P < 0.05); **极显著 (P < 0.01); MAP:mean annual precipitation,MAT:mean annual temperature,ALT: altitude,NPP: net primary productivity) | |||||
在对年均温度、年均降水量、林龄、林分密度和净生产力统一进行自然对数转化的基础上, 应用R中的lavaan包建立结构方程模型, 结果如图 4所示 (图 4中直线为结构方程隐变量与显变量的线性关系, 线上数字为通径系数)。年均降水量、年均温度、林龄、林分密度对杉木林净生产力都有直接影响, 他们之间的总通径系数分别为0.398 (P < 0.01)、0.746 (P < 0.01)、-0.321 (P < 0.01) 和-0.738 (P < 0.01)。年均降水量和年均温度的通径系数为0.633 (P < 0.01), 林分密度与林龄的通径系数为-0.514 (P < 0.01)。由此可知, 年均温度和林龄不仅直接影响杉木林净生产力, 还通过影响年均降水量和林分密度间接影响林分净生产力。年均温度和林龄的直接通径系数分别为0.494 (P < 0.01) 和-0.700 (P < 0.01);年均温度和林龄的间接通径系数分别为0.252 (P < 0.05) 和0.379 (P < 0.05) (表 2)。结果方程分析结果显示, 影响杉木林净初级生产力变化因素的62%来自年均温度、年均降水量、林龄和林分密度。

|
| 图 4 杉木林净初级生产力结构方程模型 Fig. 4 Results of structural equation models for NPP of Cunninghamia lanceolata forests 直线为结构方程隐变量与显变量的线性关系, 线上数字为通径系数; **显著相关 (P < 0.01) |
| 隐变量Endogenous variable | 显变量Exogenous variable | 直接系数Direct Effects | 标准差SD | 间接系数Indirect effects | 标准差SD | 总系数Total effect | 标准差SD |
| 生产力NPP | 年均降雨量MAP | 0.398 | 0.008 | 0.398 | 0.008 | ||
| 年均温度MAT | 0.494 | 0.009 | 0.252 | 0.061 | 0.746 | 0.183 | |
| 林龄Age | -0.700 | 0.009 | 0.379 | 0.120 | -0.321 | 0.203 | |
| 密度Density | -0.738 | 0.008 | -0.738 | 0.008 |
通过收集筛选的155篇644条杉木林生产力数据, 在对年均温度、年均降水量、林龄、林分密度和净生产力统一进行自然对数转化的基础上, 分析杉木林净初级生产力与年均降雨量、年均温度、林分密度和林龄之间的关系。杉木林净生产力与年均降水量和年均温度呈显著正相关, 这与多数学者的研究结论一致[28-31]。环境因子中, 杉木样地研究区年均降水量从西到东逐渐降低, 年均气温从南到北逐渐降低, 这种分布差异直接影响森林净初级生产力的区域格局[5, 32]。
选择结构方程模型分析杉木净初级生产力与年均降雨量、年均温度、林分密度和林龄之间的关系, 而不是单独建立生产力与各因子的模型, 主要有以下两个原因[33-34]:一是杉木林净初级生产力样本在年均降雨量、年均温度、林分密度和林龄下是否符合正态分布。因为样本的分布特征直接影响结果分析。本研究中杉木林净初级生产力样地在年均降雨量、年均温度和林分密度下呈现明显的正态分布, 但在林龄下正态分布不明显。如果直接使用林龄因子分析, 模型的预测就会存在异方差现象[35]。因此, 在建立结构方程模型时, 对年均温度、年均降水量、林龄、林分密度和净生产力统一进行自然对数转化[26]。二是单独建立生产力与各因子的模型, 势必忽略因子之间的间接影响[35-36]。因为净初级生产力与年均降雨量、年均温度、林分密度和林龄之间存在显著相关, 而且年均降雨量、年均温度和林分密度之间也存在显著相关。
结构方程模型很好的解析杉木净初级生产力与环境因子和林分因子之间的关系。它不仅可以分析净初级生产力与年均降雨量、年均温度、林分密度和林龄之间的直接影响系数, 还可以分析年均降雨量、年均温度、林龄和林分密度之间的间接影响系数。结果显示, 影响杉木林净初级生产力变化因素的62%来自年均降水量、年均温度、林龄和林分密度。年均降水量、年均温度、林龄、林分密度对杉木林净生产力都有直接影响, 其总通径系数分别为0.398 (P < 0.01)、0.746 (P < 0.01)、-0.321 (P < 0.01) 和-0.738 (P < 0.01)。同时, 年均温度和林龄不仅直接影响杉木林净初级生产力, 还通过影响年均降水量和林分密度间接影响林分净生产力。年均温度和林龄的直接通径系数分别为0.494 (P < 0.01) 和-0.700 (P < 0.01);年均温度和林龄的间接通径系数分别为0.252 (P < 0.05) 和0.379 (P < 0.05)。
杉木净初级生产力与环境因子和林分因子之间结构方程的分析结果还表明存在其他重要因子影响杉木林净初级生产力。Kang等[26]利用结构方程模型, 研究生物量与年均温度、年均降水量、土壤有机质、全氮和pH值的关系中, 得出生物量变化因素的89%与其有关, 其中土壤有机质、全氮和pH值的影响值为0.31。Lamb等[37]利用结构方程模型, 研究氮沉降与土壤温度、土壤湿度、土壤有机质、全氮和pH值的关系中, 得出氮沉变化因素的39%与土壤有机质、全氮和pH值有关。聂道平[29]、孙长忠和沈国舫[30-31]对杉木人工林生产力评价中, 认为立地条件是影响杉木林净初级生产力的重要因素。因此, 如何考虑土壤理化性质和立地条件对杉木林净初级生产力的影响, 完善影响杉木林净初级生产力的变化因素分析, 成为下一步研究内容之一。
| [1] | Bloom A J, Chapin Ⅲ F S, Mooney H A. Resource limitation in plants-an economic analogy. Annual Review of Ecology and Systematics, 1985, 16(1): 363–392. DOI:10.1146/annurev.es.16.110185.002051 |
| [2] | Cornelissen J H C, Castro-Díez P, Carnelli A L. Variation in relative growth rate among woody species//Lambers H, Poorter H, van Vuuren M M I, eds. Inherent Variation in Plant Growth:Physiological Mechanisms and Ecological Consequences. Leiden, the Netherlands:Backhuys, 1998:363-392. |
| [3] | Brown S. Measuring carbon in forests:current status and future challenges. Environmental Pollution, 2002, 116(3): 363–372. DOI:10.1016/S0269-7491(01)00212-3 |
| [4] | Poorter H, Sack L. Pitfalls and possibilities in the analysis of biomass allocation patterns in plants. Frontiers in Plant Science, 2012, 3: 259. |
| [5] | Enquist B J, Niklas K J. Global allocation rules for patterns of biomass partitioning in seed plants. Science, 2002, 295(5559): 1517–1520. DOI:10.1126/science.1066360 |
| [6] | Enquist B J, Allen A P, Brown J H, Gillooly J F, Kerkhoff A J, Niklas K J, Price C A, West G B. Biological scaling:does the exception prove the rule?. Nature, 2007, 445(7127): E9–E10. DOI:10.1038/nature05548 |
| [7] | Read J, Stokes A. Plant biomechanics in an ecological context. American Journal of Botany, 2006, 93(10): 1546–1565. DOI:10.3732/ajb.93.10.1546 |
| [8] | 刘世荣, 郭泉水, 王兵. 中国森林生产力对气候变化响应的预测研究. 生态学报, 1998, 18(5): 478–483. |
| [9] | 盛炜彤, 范少辉. 人工林长期生产力保持机制研究的背景、现状和趋势. 林业科学研究, 2004, 17(1): 106–115. |
| [10] | 赵敏, 周广胜. 中国北方林生产力变化趋势及其影响因子分析. 西北植物学报, 2005, 25(3): 466–471. |
| [11] | 朴世龙, 方精云, 郭庆华. 1982-1999年我国植被净第一性生产力及其时空变化. 北京大学学报:自然科学版, 2001, 37(4): 563–569. |
| [12] | Cramer W, Kicklighter D W, Bondeau A, Iii B M, Churkina G, Nemry B, Ruimy A, Schloss A L. Comparing global models of terrestrial net primary productivity (NPP):overview and key results. Global Change Biology, 1999, 5(S1): 1–15. DOI:10.1046/j.1365-2486.1999.00009.x |
| [13] | Prince S D, Goward S N. Global primary production:a remote sensing approach. Journal of Biogeography, 1995, 22(4/5): 815–835. DOI:10.2307/2845983 |
| [14] | 朴世龙, 方精云, 郭庆华. 利用CASA模型估算我国植被净第一性生产力. 植物生态学报, 2001, 25(5): 603–608. |
| [15] | 朱文泉, 潘耀忠, 张锦水. 中国陆地植被净初级生产力遥感估算. 植物生态学报, 2007, 31(3): 413–424. |
| [16] | 朱文泉, 潘耀忠, 阳小琼, 宋国宝. 气候变化对中国陆地植被净初级生产力的影响分析. 科学通报, 2007, 52(21): 2535–2541. DOI:10.3321/j.issn:0023-074x.2007.21.012 |
| [17] | 国家林业局. 中国森林资源报告 (2009-2013). 北京: 中国林业出版社, 2014: 10–16. |
| [18] | Luo Y J, Wang X K, Zhang X Q, Booth T H, Lu F. Root:shoot ratios across China's forests:forest type and climatic effects. Forest Ecology and Management, 2012, 269: 19–25. DOI:10.1016/j.foreco.2012.01.005 |
| [19] | 侯振宏, 张小全, 徐德应, 于澎涛. 杉木人工林生物量和生产力研究. 中国农学通报, 2009, 25(5): 97–103. |
| [20] | Reich P B, Luo Y J, Bradford J B, Poorter H, Perry C H, Oleksyn J. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(38): 13721–13726. DOI:10.1073/pnas.1216053111 |
| [21] | 左舒翟, 任引, 王效科, 张小全, 罗云建. 中国杉木林生物量估算参数及其影响因素. 林业科学, 2014, 50(11): 1–12. |
| [22] | Poorter H, Jagodzinski A M, Ruiz-Peinado R, Kuyah S, Luo Y J, Oleksyn J, Usoltsev V A, Buckley T N, Reich P B, Sack L. How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents. New Phytologist, 2015, 208(3): 736–749. DOI:10.1111/nph.13571 |
| [23] | Olive R L. Satisfaction:A Behavioral Perspective on the Consumer. New York:McGraw Hill, 1997. |
| [24] | Woodruff R B, Gardial S F. Know Your Customer:New Approaches to Understanding Customer Value and Satisfaction. Cambridge, Massaehusetts:Blackwell Business, 1996. |
| [25] | 侯杰泰, 成子娟. 结构方程模型的应用及分析策略. 心理学探新, 1999, 19(1): 54–59. |
| [26] | Kang M Y, Dai C, Ji W Y, Jiang Y, Yuan Z Y, Chen H Y H. Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia Grasslands, China. PLoS One, 2013, 8(7): e69561. DOI:10.1371/journal.pone.0069561 |
| [27] | R Core Team. R:a language and environment for statistical computing, version 3.1.2. Lavaan, China:R Foundation for Statistical Computing. 2015, 11, 7. URL https://cran.r-project.org/web/packages/lavaan/index.html. |
| [28] | 戴尔阜, 李双元, 吴卓, 闫浩文, 赵东升. 中国南方红壤丘陵区植被净初级生产力空间分布及其与气候因子的关系——以江西省泰和县为例. 地理研究, 2015, 34(7): 1222–1234. |
| [29] | 聂道平. 不同立地条件的杉木人工林生产力和养分循环. 林业科学研究, 1993, 6(6): 643–648. |
| [30] | 孙长忠, 沈国舫. 我国主要树种人工林生产力现状及潜力的调查研究Ⅰ.杉木、马尾松人工林生产力研究. 林业科学研究, 2000, 13(6): 613–621. |
| [31] | 孙长忠, 沈国舫. 我国人工林生产力问题的研究Ⅱ.影响我国人工林生产力的人为因素与社会因素探讨. 林业科学, 2000, 37(4): 26–34. DOI:10.11707/j.1001-7488.20000405 |
| [32] | Enquist B J, Allen A P, Brown J H, Gillooly J F, Kerkhoff A J, Niklas K J, Price C A, West G B. Biological scaling:does the exception prove the rule?. Nature, 2007, 445(7127): E9–E10. DOI:10.1038/nature05548 |
| [33] | 侯杰泰, 温忠麟, 成子娟. 结构方程模型及其应用. 北京: 教育科学出版社, 2004. |
| [34] | Clough Y. A generalized approach to modeling and estimating indirect effects in ecology. Ecology, 2012, 93(8): 1809–1815. DOI:10.1890/11-1899.1 |
| [35] | Harrell F E Jr, Lee K L, Mark D B. Multivariable prognostic models:issues in developing models, evaluating assumptions and adequacy, and measuring and reducing errors. Statistics in Medicine, 1996, 15(4): 361–387. DOI:10.1002/(SICI)1097-0258(19960229)15:4<361::AID-SIM168>3.0.CO;2-4 |
| [36] | Coupland R T. Grassland Ecosystems of the World:Analysis of Grasslands and Their Uses:International Biological Programme Synthesis Series. Cambridge: Cambridge University Press, 1979: 41–45. |
| [37] | Lamb E G, Mengersen K L, Stewart K J, Attanayake U, Siciliano S D. Spatially explicit structural equation modeling. Ecology, 2014, 95(9): 2434–2442. DOI:10.1890/13-1997.1 |