 2017, Vol. 37
2017, Vol. 37文章信息
- 封焕英, 范少辉, 苏文会, 杜满义, 刘广路, 毛超
- FENG Huanying, FAN Shaohui, SU Wenhui, DU Manyi, LIU Guanglu, MAO Chao.
- 不同经营方式下毛竹光合特性分异研究
- Photosynthetic characteristics of Phyllostachys edulis under different management modes
- 生态学报. 2017, 37(7): 2307-2314
- Acta Ecologica Sinica. 2017, 37(7): 2307-2314
- http://dx.doi.org/10.5846/stxb201512012407
-
文章历史
- 收稿日期: 2015-12-01
- 网络出版日期: 2016-08-30





2. 国际竹藤中心 竹藤科学与技术重点实验室, 北京 100102
2. International Centre for Bamboo and Rattan, Key Laboratory on the Science and Technology of Bamboo and Rattan, Beijing 100102, China
光合作用是植物获得干物质和产量的基础, 也是森林生态系统碳收入的唯一途径和碳素循环开始的关键环节[1-2], 因此, 较高的光合能力是植物获得高产的前提, 也是决定森林生产力的重要生理生态过程[3]。光合作用是个复杂的过程, 受光照、温度、水分、养分等多种因素的影响和制约, 同一植物不同冠层因光照差异光合能力表现出明显的异质性[4-6];其他条件相同, 温度改变能显著影响植物光合光响应曲线[7-8];较重的干旱胁迫和淹水灌溉均会降低叶片光合能力[9-10], 土壤相对含水量为56.3%—80.9%时, 山杏能维持强光下较高的光合作用水平[11]等。实际生产中, 如何提高植物光合能力越来越受到人们的关注。相关研究表明, 人为干扰 (经营) 能显著影响植物的光合能力, 垄作、间作等经营方式能增大光的截获量, 提高土壤水分含量和肥料利用率, 延长农作物叶片功能期, 提高产量[12-17];合理的施肥能有效提高林木、农作物、灌木等叶片的光合性能[18-21], 但是由于不同植物的光合能力及光合作用所需条件不同[22-26], 因此应根据植物特性选择有效经营方式, 以利于实现植物生产力提高、多样性发展及可持续经营的要求。
毛竹 (Phyllostachys eduils) 作为一种人为干扰相对频繁的森林类型, 其林地光合特性必然受到经营方式的影响, 而目前对毛竹光合特性的研究, 仅限于施肥[27-28]或某单一经营方式[29-30], 尚缺乏毛竹经营整体趋势的了解。本研究从目前粗放经营、一般经营及集约经营的几类毛竹林为对象, 探讨不同程度的人为干扰下毛竹光合特性的适应性变化, 解析不同经营方式下毛竹的生态适应性, 以期评价不同经营方式对毛竹生长的影响, 筛选不同经营目标下最有效的经营方式, 为毛竹林高产、高效经营研究奠定理论基础。
1 研究区与研究方法 1.1 研究区概况安徽省黄山区位于安徽省南部, 118°14′—118°21′E, 32°4′—32°10′N, 竹资源分布集中。研究区位于谭家桥镇黄山公益林场, 全场毛竹34万株, 竹林面积约20hm2。平均海拔450—550 m, 坡度0—15 °, 成土母岩主要为千枚岩, 含部分花岗岩及少量石灰岩, 土壤自上及下分布为山地黄壤、山地黄棕壤、黄红壤。属亚热带湿润性气候, 年均温15.3 ℃, 月平均最低气温3 ℃, 月平均最高气温27.8 ℃, 极端最高气温达40.3 ℃, 极端最低气温可至-13.5 ℃, 年降水量1376—1649 mm, 以春夏多, 秋冬少, 年蒸发量约1120 mm, 干旱指数0.4—1.2之间, 相对湿度在80%以上, 无霜期220 d。年日照2281—2453 h, 占可照时数40%。试验地土壤pH为4.7—5.2, 速效P为3.61 mg/kg, 有效N为89.55 mg/kg, 有机质含量为35.30 g/kg, 速效K为175.62 mg/kg。植被基本为毛竹纯林, 分布有少量的马尾松 (Pinus massoniana)、豹皮樟 (Litsea coreana Lévl. var. sinensis)、枫香 (Liquidambar formosa Hance)、枫杨 (Pterocary stenoptera)、杉木 (Cunninghamia lanceolata)、苦槠 (Castanopsis sclerophylla)、乌药 (Lindera aggregata) 等。
1.2 试验设计以黄山区目前主要经营类型的5类毛竹林为研究对象, 设置20 m×30 m标准样地, 并调查基本情况 (表 1)。每类毛竹林设3个重复, 每个重复内选3株生长健康、径粗为平均胸径的3年生毛竹作为标准样竹, 测第12—14轮枝条的3—5片健康叶片。
| 编号No. | 经营方式Management mode | 经营现状Management status | 大小年是否分明On-year or off-year | 经营年限Management age/a | 平均胸径Mean DBH/cm | 林分密度Stand density/(株/hm2) | 备注Remark |
| Ⅰ | 粗放经营 | 约6—8a采伐1次, 无挖笋、劈草等 | 是 | >40 | 8.82 | 3380 | 位于大坪, 以发挥生态功能为主 |
| Ⅱ | 一般经营 | 每年采伐1次, 皆伐2度以上生长良好竹子, 挖春笋量小于5%, 挖冬笋量小于3% | 是 | 25—30 | 8.35 | 3200 | 位于后背山, 以材用为主 |
| Ⅲ | 集约经营 | 每隔1年采伐1次, 按比例留养母竹, 挖冬笋量10%—15%, 挖春笋量约为15% | 是 | 25—30 | 8.50 | 4129 | 位于留东14队, 兼具笋材两用 |
| Ⅳ | 集约经营 | 每隔1年采伐1次, 按比例留养母竹, 挖冬笋量15%—20%, 挖春笋量约为15% | 否 | 20—25 | 8.53 | 3450 | 位于留东13队, 兼具笋材两用 |
| Ⅴ | 集约经营 | 每隔1年采伐1次, 每年9月施氮肥1次, 按比例留养母竹, 挖冬笋量约为10% | 是 | 30—35 | 9.60 | 3826 | 位于大坪, 以材用为主 |
光合参数测定:在毛竹大年生长旺季 (8月3—25日), 光照充足的晴朗天气, 每天7:30—11:00时, 用Li-6400标准叶室和红蓝光源, 测定各标准样竹光响应曲线。为保证其它条件尽量一致, 观测过程中, 根据外界环境条件控制叶室内温度为28 ℃, CO2浓度为400 μmol/μmol, 流速为500 μmol/s, 光合有效辐射从弱到强依次为0、30、50、100、200、300、500、800、1200、1500、1800、2200 μmol m-2 s-1, 每个光合有效辐射下适应3—5 min后进行测定。为避免毛竹叶片不能完全充满叶室造成的误差, 对测定叶片编号标记并剪掉带回, 通过方格纸计算叶面积, 用以重新计算各光合参数。由于毛竹具有较强的柔韧性, 本研究将不同经营方式下的目标竹提前一天选出, 拉弯竹梢以备活体测定其光合特性。测定主要参数有:(1) 光合特性因子:净光合速率Pn、蒸腾速率Tr、气孔导度Cd、胞间CO2浓度Ci、叶片温度Tleaf。(2) 环境因子:光合有效辐射PAR、空气温度Ta、空气相对湿度RH、空气CO2浓度Ca。
叶绿素含量的测定:用SPAD-520荧光仪, 选择目标竹12—14轮枝条上生长健康叶片, 30片为一组求平均值, 每竹测90片叶片。
叶面积指数 (LAI) 的测定:用Hemview数字冠层仪, 在每个标准样地的对角线及中心选5个固定样点 (尽量避免照片视野的交叉, 保证整个样地都在拍摄范围内), 于天气晴朗的6:30—8:30、16:30—18:00以及光线均匀的阴天全天进行测定 (避免强烈的太阳光直射产生的光斑对图片曝光率过高的影响), 保持冠层仪水平状态, 距地面约1.5 m (排除地面灌草层的干扰), 每个固定样点测3次, 然后通过自带分析软件计算叶面积指数。
1.4 数据处理在excel2010中进行数据初步分析, 计算各经营方式下不同光强的平均净光合速率, 然后用非线性模型拟合Pn-PAR, 求得相应的光响应曲线及光合参数。相关统计分析在SPSS 16.0统计软件完成。
非线性曲线模型:
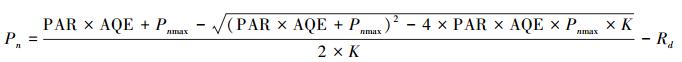
|
式中, Pn为净光合速率 (μmol m-2 s-1), PAR为光合有效辐射强度 (μmol m-2 s-1), Pnmax为最大净光合速率 (μmol m-2 s-1), AQE为表观量子效率 (μmol/μmol), K为曲角, Rd为暗呼吸速率 (μmol m-2 s-1)。
2 结果与分析 2.1 不同经营方式下毛竹叶面积指数和叶片SPAD特性 2.1.1 毛竹叶面积指数图 1可知, 毛竹叶面积指数在不同经营方式下依次表现为Ⅳ(1.76)>Ⅴ(1.63)>Ⅰ(1.56)>Ⅱ(1.54)>Ⅲ(1.39), 且差异显著 (P < 0.05)。其中Ⅳ显著高于Ⅰ—Ⅲ, 但与Ⅴ间差异不显著;Ⅲ的叶面积指数则显著低于其它四种经营。说明粗放经营 (Ⅰ) 和一般经营 (Ⅱ) 两种方式下毛竹林叶面积指数差异不大;而不同集约经营手段对毛竹林叶面积则产生较大差别, 其中, 对每年有计划挖笋伐竹的毛竹林 (Ⅲ和Ⅳ), 大小年是否分明显著区分了该种经营模式对竹林叶面积指数的影响, 大小年不分的毛竹林有更高的叶面积指数;对大小年分明的毛竹林而言 (Ⅲ和Ⅴ), 施肥方式下毛竹林的叶面积指数显著高于仅挖笋伐竹经营的毛竹林。

|
| 图 1 不同经营方式下毛竹叶面积指数 Fig. 1 Bamboo forest LAI index under different management modes Ⅰ:粗放经营;Ⅱ:一般经营;Ⅲ:集约经营,挖冬笋量10%—15%;Ⅳ:集约经营,挖冬笋量15%—20%;Ⅴ:集约经营,挖冬笋量约10% |
SPAD值是通过测定植物叶片叶绿素的相对含量或者“绿色程度”来了解植物真实的硝基需求量即氮的需求。图 2表明各经营方式下, Ⅱ和Ⅲ的SPAD值显著低于其它处理, 说明该两种经营模式下叶片中氮含量相对较低, 可能是因为大小年分明的毛竹林, 挖笋、伐竹强度过大, 对林内N的循环和利用有较大的影响。对于粗放经营的毛竹林来讲, 内部养分和物质的循环存在不被轻易打破的平衡。

|
| 图 2 不同经营方式下毛竹叶片SPAD值 Fig. 2 Bamboo leaf SPAD value under different management modes |
图 3为5种不同经营方式毛竹林光响应曲线拟合曲线, 各方程决定系数均大于0.96, 说明非线性曲线模型可有效反映不同经营方式毛竹的光合特性。从各曲线走向可知, 不同经营方式下毛竹的净光合速率 (Pn) 随光合有效辐射 (PAR) 变化趋势基本一致, 即均表现为随PAR增强, Pn先迅速增大, 达到一定水平后, Pn增速平缓, 直至Pn不再随PAR增强而增大。同一PAR下, 5种经营方式间Pn值总体表现为, Ⅴ最大, Ⅰ和Ⅱ次之, Ⅲ和Ⅳ较小。说明不同经营方式下毛竹对PAR响应存在差异, 其中经营方式Ⅴ下毛竹对PAR的响应最突出, 一定程度上说明适当补充养分 (Ⅴ) 能有效提高毛竹对光能利用能力, 粗放经营 (Ⅰ) 和一般经营 (Ⅱ) 对光能利用能力接近, 对毛竹光能利用能力稍强于对林地干扰相对较强的仅有计划挖笋伐竹的集约经营 (Ⅲ和Ⅳ)。

|
| 图 3 不同经营方式下毛竹的光响应曲线 Fig. 3 Bamboo light response curve under different management modes |
随光合有效辐射增强, 不同经营方式下毛竹叶片的气孔导度均呈上升趋势, PAR小于200μmol m-2 s-1时, 气孔导度增速最快, PAR大于1200μmol m-2 s-1时, 气孔导度增速缓慢 (图 4)。从曲线趋势来看, 不同经营方式间气孔导度总体表现为Ⅱ(0.022—0.174μmol/μmol)> Ⅰ(0.042—0.168μmol/μmol)> Ⅴ(0.031—0.161μmol/μmol)> Ⅲ(0.027—0.141μmol/μmol)> Ⅳ(0.022—0.133μmol/μmol), Ⅰ、Ⅱ和Ⅴ 3种经营下气孔导度增大较高于Ⅲ和Ⅳ, 说明在无适当养分补充, 仅挖笋伐竹的集约经营 (Ⅲ和Ⅳ) 下, 毛竹气孔导度相对粗放经营 (Ⅰ) 和一般经营 (Ⅱ) 的毛竹林较低, 实际经营中可根据需要有效补充林地养分或降低干扰水平来改善毛竹气孔导度。

|
| 图 4 不同经营方式下毛竹的气孔导度 Fig. 4 Bamboo Cd under different management modes |
随光合有效辐射增强, 毛竹叶片的胞间CO2浓度总体呈下降趋势 (图 5)。当PAR小于200μmol m-2 s-1时, 下降迅速, 大于800μmol m-2 s-1时, 趋于稳定。5种经营方式中, Ⅴ在PAR小于200μmol m-2 s-1时胞间CO2浓度下降最快, 说明该经营方式下毛竹在弱光条件下具较高的光合效率。随PAR增强胞间CO2浓度趋于稳定后, Ⅰ、Ⅱ和Ⅴ经营方式下的值较接近, Ⅲ和Ⅳ经营方式下的值较接近, 但总体差异小于88μmol/μmol。

|
| 图 5 不同经营方式下毛竹的胞间CO2浓度 Fig. 5 Bamboo Ci under different management modes |
从图 6可知, 随光合有效辐射增强, 毛竹叶片蒸腾速率逐渐上升, 低光强 (小于200μmol m-2 s-1) 下, 蒸腾速率上升较快, 然后逐渐缓慢。不同经营方式下毛竹的蒸腾速率总体表现为, Ⅱ>Ⅰ>Ⅴ>Ⅳ>Ⅲ, 与气孔导度具相似的规律。

|
| 图 6 不同经营方式下毛竹的蒸腾速率 Fig. 6 Bamboo Tr under different management modes |
从表 2可知, 不同经营方式下毛竹光响应参数之间存在较大差异。其中最大净光合速率 (Pnmax) 表现为Ⅰ>Ⅴ>Ⅱ>Ⅳ>Ⅲ, 集约经营中的施肥经营 (Ⅴ) 与粗放经营 (Ⅰ) 的Pnmax明显高于其它3种处理, 且一般经营 (Ⅱ) 也高于集约经营中的Ⅲ和Ⅳ。表观量子效率在不同经营方式下为0.029—0.034, 各处理间差异较小, 说明毛竹在弱光下吸收、转换和利用光能的能力虽有不同, 但受经营方式的影响不明显。光补偿点 (LCP) 和光饱和点 (LSP) 在各经营下差异较大, 与粗放经营 (Ⅰ) 相比, 一般经营 (Ⅱ) 和集约经营 (Ⅳ、Ⅴ) 下的毛竹的LCP均较小, 总体表现为Ⅰ>Ⅲ>Ⅳ>Ⅱ>Ⅴ;LSP在各处理下的表现与LCP不同, Ⅰ和Ⅳ的值接近, 明显高于集约经营下的Ⅲ和Ⅴ, 一般经营下毛竹的LSP与Ⅳ接近, 即Ⅰ>Ⅳ>Ⅱ>Ⅴ>Ⅲ, 因此, 一定程度上说明长期不同经营方式下毛竹林内光照、温度、水分等环境因素的不同干扰到毛竹对光的利用特性, 根据不同的环境特性, 毛竹光合生理特性做出一定的调整。粗放经营的毛竹林可能更倾向于增强对强光利用的能力, 一般经营毛竹林对强光和弱光均有较强的适应性, 集约经营的毛竹林对光的反应差异较大, 这可能与集约经营侧重点不同, 从而使毛竹光合响应呈现多元化。暗呼吸速率Rd在各毛竹林中表现为Ⅰ最强, Ⅴ明显低于所有经营, 一定程度上说明粗放经营毛竹林无光照条件下仍具有较强的呼吸速率, 而集约经营中的施肥经营 (Ⅴ) 有效降低了毛竹无光照下的呼吸速率。
| 编号No. | Pnmax/(μmol m-2 s-1) | AQE/(μmol m-2 s-1) | LSP/(μmol m-2 s-1) | LCP/(μmol m-2 s-1) | Rd/(μmol m-2 s-1) |
| Ⅰ | 13.56±1.078 | 0.033±0.008 | 699.92±80.12 | 77.76±11.26 | 1.939±0.313 |
| Ⅱ | 12.77±0.372 | 0.033±0.005 | 649.62±63.74 | 49.95±10.32 | 1.316±0.222 |
| Ⅲ | 11.71±0.513 | 0.029±0.006 | 571.93±60.11 | 66.70±10.10 | 1.220±00319 |
| Ⅳ | 11.87±0.283 | 0.034±0.006 | 690.38±81.02 | 53.96±12.37 | 1.381±0.215 |
| Ⅴ | 13.36±0.484 | 0.031±0.004 | 584.55±55.46 | 44.42±9.12 | 1.057±0.215 |
毛竹具有生长快、周期短、用途广的特性, 因此以直接经济利益为导向的经营方式在毛竹林中最常见, 即通常以竹材或笋材为主要目的隔年采伐、挖笋, 经营方式相对粗放, 存在一定的盲目性。采伐影响毛竹林地土壤呼吸和养分盈亏, 相关研究表明, 生长季 (8月) 对毛竹林采伐, 能显著提高土壤温度、土壤呼吸和凋落物呼吸, 降低根系呼吸[31];年均采伐量为9.0 t·hm-2时, 林地内N、P、K、Ca、Mg、Si等6种元素的亏损总量将达到7.96 kg/hm2[32]。同时采伐亦改变了毛竹林空间结构和林分密度, 对林内光照和养分分配必将产生一定的影响。长期不同程度的采伐经营, 对毛竹林内土壤呼吸、养分分配、温度及光环境产生习惯性影响, 这些因素与毛竹光合作用的基本需求密不可分, 长期不同环境的适应对毛竹自身光合特性是如何影响的, 是一个值得深入思考的问题。本研究结果表明, 不同的干扰水平, 毛竹光合特性间存在着较大的差异, 粗放经营毛竹林 (Ⅰ) 的Pn、Cd、Tr、LSP、LCP和Rd较其他经营方式相对高;一般经营毛竹林 (Ⅱ) 的Pn、Cd和Tr比集约经营的Ⅲ和Ⅳ高;集约经营的3类林地间存在较大差别, 其中Ⅲ的LAI和SPAD显著低于Ⅳ和Ⅴ, Ⅳ和Ⅴ差异不显著, Ⅴ的Pn显著高于Ⅲ和Ⅳ, Rd显著低于Ⅲ和Ⅳ。说明人为干扰是毛竹林光合能力产生差异的重要原因, 在人为干扰程度低的粗放经营毛竹林中, 存在着系统的自我更新和平衡, 符合系统生态学的原则[33-34]。平衡的生态系统一旦加入人为因素, 发展也易出现多元化[35-37], 集约经营中3类林地光合能力的不同表现有力的印证了这一规律。结合毛竹自身特性和立地条件, 适当补充林地养分经营的毛竹林 (Ⅴ) 有较高的光合能力, 这与相关研究结论一致, 但对于施肥时间、施肥量和施肥频率的研究目前仍未有统一结论[28, 38]。
大小年现象是毛竹林的一个典型特点, 而产生这种现象的内在原因目前仍未有统一的认识。本研究以大小年是否分明的两类毛竹林为研究对象, 探讨其光合特性的分异, 结果表明, 相同经营方式下, 大小年分明毛竹林 (Ⅲ) 的LAI和SPAD值显著低于大小年不分明毛竹林 (Ⅳ), 二者的Pn、Cd、Tr及Ci间表现出一致的趋势, 但Ⅳ的LSP显著高于Ⅲ, 且LCP显著低于Ⅲ, 一定程度上反映了大小年不分的毛竹林 (Ⅳ) 对强光具有较强的适应性, 是否能作为区分毛竹林大小年的一个重要结论仍需进一步研究。大小年不分的毛竹林每年发笋成竹, 林内单竹生存空间变化相对频繁, 而林内空间的竞争强度与林分密度、叶面积指数呈正相关关系[39-40], 本文研究结果表明, 相同经营方式下, 大小年不分的毛竹林有更高的叶面积指数, 也进一步说明了大小年不分的林分类型可能具更高适应性。
隔年采伐、挖笋对毛竹林空间结构和土壤结构扰动较大, 林内环境具有不确定性[38]。本研究5种类型毛竹林的光合特性分异反映了不同经营方式下毛竹长期对光、水分、养分等环境的适应特性, 因此毛竹林经营中, 应根据实际情况, 选择有效经营措施, 充分发挥毛竹林的经济、生态和社会效益。
致谢: 感谢刘碧桃、陈红老师对本研究给予的支持和帮助。| [1] | Gifford R M, Evans L T. Photosynthesis, carbon partitioning, and yield. Annual Review of Plant Physiology, 1981, 32(1): 485–509. DOI:10.1146/annurev.pp.32.060181.002413 |
| [2] | Tuittila E S, Vasander H, Laine J. Sensitivity of C sequestration in reintroduced Sphagnum to water-level variation in a cutaway peatland. Restoration Ecology, 2004, 12(4): 483–493. DOI:10.1111/rec.2004.12.issue-4 |
| [3] | 许大全. 光合作用效率. 上海: 上海科学技术出版社, 2002. |
| [4] | Cao Y H, Zhou B Z, Wang X M, Wang G. The Photosynthetic characteristics of Moso Bamboo (Phyllostachys pubescens) for different canopy leaves. Advanced Materials Research, 2013, 726-731: 4274–4279. DOI:10.4028/www.scientific.net/AMR.726-731 |
| [5] | Van de Weg M J, Meir P, Grace J, Ramos G D. Photosynthetic parameters, dark respiration and leaf traits in the canopy of a Peruvian tropical montane cloud forest. Oecologia, 2012, 168(1): 23–34. DOI:10.1007/s00442-011-2068-z |
| [6] | Niinemets V. Optimization of foliage photosynthetic capacity in tree canopies:towards identifying missing constraints. Tree Physiology, 2012, 32(5): 505–509. DOI:10.1093/treephys/tps045 |
| [7] | 郑益兴, 彭兴民, 张燕平. 印楝不同种源对温度变化的光合生理生态响应. 林业科学研究, 2008, 21(2): 131–138. |
| [8] | Yamori W, Hikosaka K, Way D A. Temperature response of photosynthesis in C3, C4, and CAM plants:temperature acclimation and temperature adaptation. Photosynthesis Research, 2014, 119(1/2): 101–117. |
| [9] | 王唯逍, 刘小军, 田永超, 姚霞, 曹卫星, 朱艳. 不同土壤水分处理对水稻光合特性及产量的影响. 生态学报, 2012, 32(22): 7053–7060. |
| [10] | Hosseinzadeh S R, Amiri H, Ismaili A. Effect of vermicompost fertilizer on photosynthetic characteristics of chickpea (Cicer arietinum L.) under drought stress. Photosynthetica, 2015: 1–6. |
| [11] | 郎莹, 张光灿, 张征坤, 刘顺生, 刘德虎, 胡小兰. 不同土壤水分下山杏光合作用光响应过程及其模拟. 生态学报, 2011, 31(16): 4499–4508. |
| [12] | 马丽, 李潮海, 付景, 郭学良, 赵霞, 高超, 王磊. 垄作栽培对高产田夏玉米光合特性及产量的影响. 生态学报, 2011, 31(23): 7141–7150. |
| [13] | 张春明, 张耀文, 郭志利, 赵雪英, 闰虎斌, 朱慧君, 高伟. 间作模式下小豆光合特征及产量效益研究. 中国农学通报, 2014, 30(3): 226–231. DOI:10.11924/j.issn.1000-6850.2013-0498 |
| [14] | 焦念元, 宁堂原, 杨萌珂, 付国占, 尹飞, 徐国伟, 李增嘉. 玉米花生间作对玉米光合特性及产量形成的影响. 生态学报, 2013, 33(14): 4324–4330. |
| [15] | Wang H X, Liu Y F, Song Q B, Lu Y, Han X R, Shi Q W. Effects of different planting modes on peanut photosynthetic characteristics, leaf area index and yield in the sandy Area//International Conference on Mechatronics, Electronic, Industrial and Control Engineering. Harold:Atlantis Press, 2015. |
| [16] | 张倩, 张洪生, 宋希云, 姜雯. 种植方式和密度对夏玉米光合特征及产量的影响. 生态学报, 2015, 35(4): 1235–1241. |
| [17] | Xiao J L, Wang G J, Zhao M, Yin J, Li W, Bi Y D, Li W, Lai Y C, Shu X T, Zhao Y. Effect of cultivation pattern on the light radiation of group canopy and yield of spring soybean (Glycine Max L. Merrill). American Journal of Plant Sciences, 2013, 4(6): 1204–1211. DOI:10.4236/ajps.2013.46148 |
| [18] | 赵燕, 董雯怡, 张志毅, 李吉跃, 聂立水, 沈应柏. 施肥对毛白杨杂种无性系幼苗生长和光合的影响. 林业科学, 2010, 46(4): 70–77. DOI:10.11707/j.1001-7488.20100411 |
| [19] | 杨自立, 马履一, 贾忠奎, 王梓. 不同供氮水平对栓皮栎播种苗光响应曲线的影响. 北京林业大学学报, 2011, 33(5): 56–60. |
| [20] | 李学刚, 宋宪亮, 孙学振, 陈二影, 张美玲, 赵庆龙, 刘峰. 控释氮肥对棉花叶片光合特性及产量的影响. 植物营养与肥料学报, 2010, 16(3): 656–662. DOI:10.11674/zwyf.2010.0320 |
| [21] | 王帅, 韩晓日, 战秀梅, 杨劲峰, 刘轶飞, 王月, 李娜. 不同氮肥水平下玉米光响应曲线模型的比较. 植物营养与肥料学报, 2014, 20(6): 1403–1412. DOI:10.11674/zwyf.2014.0610 |
| [22] | 师生波, 李惠梅, 王学英, 岳向国, 徐文华, 陈桂琛. 青藏高原几种典型高山植物的光合特性比较. 植物生态学报, 2006, 30(1): 40–46. |
| [23] | 王暾, 郭晋平, 刘宁, 张芸香. 森林光环境对4种天然灌木的光合作用和形态的影响. 林业科学, 2011, 47(6): 56–63. DOI:10.11707/j.1001-7488.20110609 |
| [24] | 张娇, 施拥军, 朱月清, 刘恩斌, 李梦, 周建平, 李建国. 浙北地区常见绿化树种光合固碳特征. 生态学报, 2013, 33(6): 1740–1750. |
| [25] | Gail Smith B, Stephens W, Burgess P J, Carr M K V. Effects of light, temperature, irrigation and fertilizer on photosynthetic rate in Tea (Camellia Sinensis). Experimental Agriculture, 1993, 29(3): 291–306. DOI:10.1017/S001447970002086X |
| [26] | 吴志庄, 杜旭华, 熊德礼, 王树东, 邹跃国, 周妍. 不同类型竹种光合特性的比较研究. 生态环境学报, 2013, 22(9): 1523–1527. |
| [27] | 高培军, 邱永华, 周紫球, 何仁华, 徐佳. 氮素施肥对毛竹生产力与光合能力的影响. 浙江农林大学学报, 2014, 31(5): 697–703. DOI:10.11833/j.issn.2095-0756.2014.05.006 |
| [28] | 宋艳冬, 金爱武, 金晓春, 胡元斌, 杜亮亮, 江志友. 施肥对毛竹叶片光合生理的影响. 浙江林学院学报, 2010, 27(3): 334–339. |
| [29] | 施建敏, 郭起荣, 杨光耀. 毛竹光合动态研究. 林业科学研究, 2005, 18(5): 551–555. |
| [30] | 王星星, 刘琳, 张洁, 王玉魁, 温国胜, 高荣孚, 高岩, 张汝民. 毛竹出笋后快速生长期内茎秆中光合色素和光合酶活性的变化. 植物生态学报, 2012, 36(5): 456–462. |
| [31] | 唐晓鹿, 范少辉, 漆良华, 刘广路, 官凤英, 杜满义, 许庆标. 采伐对幕布山区毛竹林土壤呼吸的影响. 林业科学研究, 2013, 26(1): 52–57. |
| [32] | 蒋俊明, 刘大雷, 范少辉, 费世民, 陈德松. 川南毛竹林生态系统养分动态分析. 南京林业大学学报:自然科学版, 2010, 34(2): 31–36. |
| [33] | 封焕英. 毛竹林健康评价指标体系构建及实证研究[D]. 北京: 中国林业科学研究院, 2014. |
| [34] | Millar C I, Stephenson N L. Temperate forest health in an era of emerging megadisturbance. Science, 2015, 349(6250): 823–826. DOI:10.1126/science.aaa9933 |
| [35] | Lindenmayer D B, Blanchard W, McBurneny L, Blair D, Banks S, Linkens G E, Franklin J F, Laurance W F, Stein J A R, Gibbons P. Interacting factors driving a major loss of large trees with cavities in a forest ecosystem. PLoS One, 2012, 7(10): e41864. DOI:10.1371/journal.pone.0041864 |
| [36] | Parmesan C, Burrows M T, Duarte C M, Poloczanska E S, Richardson A J, Schoeman D S, Singer M C. Beyond climate change attribution in conservation and ecological research. Ecology Letters. 2013: 58–71. |
| [37] | 袁菲, 张星耀, 梁军. 基于有害干扰的森林生态系统健康评价指标体系的构建. 生态学报, 2012, 32(3): 964–973. |
| [38] | 苏文会. 基于生长和养分积累规律的毛竹林施肥理论与实践研究[D]. 北京: 中国林业科学研究院, 2012. |
| [39] | 仇建习, 汤孟平, 沈利芬, 娄明华, 庞春梅. 近自然毛竹林空间结构动态变化. 生态学报, 2014, 34(6): 1444–1450. |
| [40] | 陈永刚, 汤孟平, 杨春菊, 马天午, 王礼. 天然毛竹林竞争空间关系分析. 植物生态学报, 2015, 39(7): 726–735. DOI:10.17521/cjpe.2015.0069 |