 2017, Vol. 37
2017, Vol. 37文章信息
- 安静 , 吴玲 , 王海娟 , 段呈 , 王绍明
- AN Jing , WU Ling , WANG Haijuan , DUAN Cheng , WANG Shaoming .
- 不同干扰程度下沙生植物异翅独尾草的种群结构与动态特征
- Structural and dynamic characteristics of Eremurus anisopterus populations in areas with different levels of disturbance
- 生态学报. 2017, 37(6): 2023-2032
- Acta Ecologica Sinica. 2017, 37(6): 2023-2032
- http://dx.doi.org/10.5846/stxb201512242572
-
文章历史
- 收稿日期: 2015-12-24
- 网络出版日期: 2016-07-21






生境破碎化是指在人为活动和自然干扰下,使原来大块连续分布的自然生境被分割、破碎,形成分散、孤立的岛状生境或生境碎片的现象[1], 从而导致原生境总面积减小,产生隔离的异质种群,影响种间基因交流、种群存活力、物种间相互作用及一系列生态过程[2-5]。生境变化是自然、社会和生物因素相互作用的结果,人类活动加剧了生境的破碎化[6-7]。生境破碎化过程引起生境小气候、边缘效应等生物与非生物条件的变化,进而影响到植物异质种群的动态[2, 8-9]。栖息地的碎化、退化和消失是目前公认的导致物种消失的重要原因,那么从人类与自然界相互作用的过程中寻找保护地方特有种的规律和方法变得越来越重要。
异翅独尾草 (Eremurus anisopterus (Kar.et Kir.) Regel) 是生长在古尔班通古特沙漠南部与天山北麓相邻的固定、半固定沙丘上的一种多年生类短命植物,是早春时节固定沙区重要的植物之一[10]。在自然生境中,该物种地下芽能形成新的个体,其利用早春 (3月底至4月初) 地面积雪融化、春雨开始、土壤湿润的有利条件,在60 d内迅速完成从萌芽到开花结果形成种子的整个生长发育过程[11-12]。虽然生活周期短暂,却是古尔班通古特沙漠早春防风固沙、稳定沙面的主要贡献者以及维持荒漠生态系统的主要参与者之一,具有十分重要的生态学意义[10, 13-14]。近年来由于大面积垦荒、过度樵采、放牧以及新疆北部油气资源的开发、道路等工程项目的建设对荒漠生态系统产生重大影响,异翅独尾草所在生境斑块化现象明显,斑块数目、斑块隔离程度增加。以往有关异翅独尾草的研究主要集中于繁殖对策、化学成分、核型分析等方面[15-19],而研究者对生境破碎后该植物种群的数量分布与动态知之甚少。王海娟等[16]分析了荒漠斑块生境中异翅独尾草种子形态特征,发现在被干扰的生境中,种子有尺寸小、质量轻、种翅长且小种子比率较高的特点,说明干扰条件已经对异翅独尾草种子的形态特征产生了影响,那么持续的人为干扰可能也会对异翅独尾草种群的结构与动态产生影响,例如遭受持续的严重的人为干扰可能造成该生境异翅独尾草的退化或消失,而未受到或受到轻度人为干扰的种群将呈稳定的增长趋势。种群结构与种群动态是种群最基本的特征,也是种群重要的特征之一,它运用种群统计学方法分析种群现有状态,不仅能反映种群动态及其发展趋势,在一定程度上也反映了种群与环境间的相关关系以及它们在种群中的地位[20-21]。其相关研究可以反映种群数量特征、存活状态,展现种群生物学特性对环境条件的适应结果,还能对种群的过去干扰和未来趋势做出估计与预测[22-27]。
本文基于对古尔班通古特沙漠中不同破碎生境下异翅独尾草种群的样地调查,分析研究其年龄结构、静态生命表、存活曲线、生存分析,了解现有异翅独尾草种群生存现状,分析种群的年龄结构特征与受干扰情况及其种群动态与内在的生活史特征和外在的干扰状况之间的互动关系,探讨其种群更新与维持机制,为揭示生境破碎化后异翅独尾草种群数量动态变化规律,合理利用和保护沙生独尾草种群提供科学依据。
1 材料与方法 1.1 研究区概况古尔班通古特沙漠 (44°11′—46°20′N, 81°31′—90°00′E) 地处半封闭的准噶尔盆地中,面积约为4.88万km2,整个沙漠面积的95%以上为固定、半固定风沙土,是我国最大的固定与半固定沙漠。地貌类型以沙垄及树枝状沙垄为主,沙丘相对高度一般为10—50 m。该沙漠年积温3000—3500 ℃,年降水量70—150 mm,年潜在蒸发量约2000 mm,为典型的内陆干早气候。该沙漠冬春两季降水量占全年30%—45%,冬季稳定积雪日数一般在100—160 d,最大积雪深度多在20 cm以上,而地下水埋藏多在30 m以下,植物生长主要靠降水而难以利用地下水,因此3月中旬气温回升、积雪融化,为早春短命植物生长发育提供良好条件[28-29]。
研究区位于新疆古尔班通古特沙漠南缘,异翅独尾草分布比较集中的炮台镇、西古城镇、十户滩镇三地。该地区地形有沙丘、丘间平地和开垦地,实验样地海拔在300—350 m之间。植被主要是新疆沙漠常见的植物,有异翅独尾草、粗柄独尾草 (Eremurus inderiensis)、角果藜 (Ceratocarpus arenarius)、木本猪毛菜 (Salsola arbuscula)、沙大戟 (Chrozophora sabulosa)、琵琶柴 (Reaumuria soongorica)、白茎绢蒿 (Seriphidium terraealbae) 等。近几十年来由于人为干扰,使该区植被景观遭到破坏,导致过去广阔连片分布的植物种群被分隔成一个个小种群。
1.2 样地的选择经充分踏查,考虑生境特点和人为干扰等因素,于2015年4月—6月分别在有异翅独尾草分布的炮台镇、西古城镇和十户滩镇设定样地,在沙垄或丘间地选择不同破碎化程度及不同受干扰程度,不同规模大小、相互之间间距大的斑块共计19个,分别记为P1、P2、P3、…、P19,走访调查各斑块主要干扰因素、干扰成因及干扰年限等,记录各样地如下信息:GPS位点、海拔高度、地形条件、斑块形状、斑块面积、斑块周长、边缘密度 (ED,单位面积上斑块的长度,其大小直接影响景观边缘效应,能反映斑块的破碎程度)、斑块敞闭度 (指斑块与周围环境的镶嵌关系,分为全敞型、三围一敞型、敞闭相齐型、三敞一闭型、全闭型)、是否与近缘种粗柄独尾草混生等,并根据实地情况对斑块破碎程度在0—1内进行赋值。用SPSS 17.0对斑块初始数据进行标准转换,采用Q分层聚类法对样地进行类型划分。
1.3 样地类型的划分基于3个样区19个斑块的10个环境因子,用SPSS 17.0分析软件的欧式距离离差平方和法做树状聚类分析 (图 1)。结果表明,聚类分析方法可以将19个斑块划分为两种类型或细化为3种类型,本文采用3种类型的分类方法分别记为a类型、b类型、c类型。

|
| 图 1 欧式距离离差平方和法分层聚类谱系树状图 Fig. 1 The Classify Deprogram of patches in Euclidean distance Ward′s method |
a类型为小面积-近全敞式-高破碎度的样地组合,有6个斑块组成:P1、P5、P7、P15、P17、P19。这些斑块受众多因素干扰,除沙漠道路、农田、放牧等干扰活动外,还有周边灌溉井、养殖场和酒庄等场所的干扰;斑块内有近缘种出现,和近缘种粗柄独尾草混合生长;斑块面积在196—682 m2之间,属于小面积斑块;斑块敞闭方式上近全敞开型;ED值高显示出边缘发育程度高,破碎程度赋值在0.80以上,破碎化程度较高。
b类型为中等面积-中等破碎度的斑块组合,包括的斑块有P2、P3、P4、P8、P13、P18。归属该类型的样地斑块特点是:样地地形近长方形;样地面积中等,面积值在1464—2925 m2之间;边缘密度 (ED) 值中等,即边缘发达程度中等;样地内有少量近缘种分布;破碎度值在0.5—0.6之间,破碎程度中等。
c类型为大面积-近全闭合-低破碎程度的斑块组合,包括P6、P9、P10、P11、P12、P14、P16的7个斑块。该类型斑块面积中等偏大,各组成斑块的面积在0.5 hm2以上;斑块边缘不发达;斑块以四周近闭合的形态存在;样地内少有近缘种生长;破碎度赋值在0.5以下,属于低破碎度的斑块类型。
1.4 研究方法 1.4.1 野外调查采用样方法对不同类型样地中异翅独尾草种群进行研究。每样地内划定3个10 m×10 m样方,调查各样方内异翅独尾草的株数,并对样方内异翅独尾草进行每株调查,调查指标为轮叶数、叶片长度以及株高等。
1.4.2 龄级划分植物种群具有其群体特征,这些特征一般都是用统计数据进行描述的,其中种群年龄 (或龄级) 分布对于种群动态的研究具有重要意义。不同的物种其龄级的确定可采用不同的方法,高大的木本植物可采用胸径或树高作为个体大小的指标,对于多年生草本植物来讲,这一工作在多数情祝下还是存在较大的困难。检索相关文献得知,百合科植物叶长和轮叶数均可作为其龄级大小的指标[30-31]。结合本次调查的实际情况,本文采用以轮叶数代替年龄的方法,用实测轮叶数来划分龄级,以三片轮叶为一个龄级,共划分为12个龄级。统计各龄级的植株数,以各龄级代表相对年龄作为纵轴,以各龄级植物个体数为横轴,绘制种群年龄结构椎体图。
1.4.3 静态生命表由于异翅独尾草属于多年生类短命植物,故采用“空间代替时间”的方法编制种群静态生命表[25-27],进而分析其动态变化。特定时间生命表各参数含义如下[32]:x为单位时间内年龄等级的中值;ax为在x龄级内现有个体数;lx为在x龄级开始时标准化存活个体数 (一般转换为1000);dx为从x到x+1龄级间隔期内标准化死亡数;qx为从x到x+1龄级间隔期间死亡率;Lx为从x到x+1龄级间隔期间还存活的个体数;Tx为从x龄级到超过x龄级的个体总数;ex为进入x龄级个体的生命期望或平均期望寿命;Kx为亏损率 (损失度)。以上各项相互关联,通过实测值ax或dx求得其余各项的数值。
由于静态生命表用同一时期收集的种群所有个体的径级编制而成,反映了多个时代重叠的年龄动态历程中的一个特定时间,而不是对这一种群全部生活史的追踪,并且在调查中存在系统误差,在生命表中会出现死亡率为负的情况。对于这种情况,Wratten认为,生命表分析中产生一些负值,这与数学假设技术不符,但仍能提供有用的生态学记录,表明种群并非静止不动,而是在迅速发展或衰落之中[33]。
1.4.4 存活曲线和死亡曲线以龄级为横轴,以计算得到的标准化存活数和死亡率为纵坐标,绘制得到该种群的存活曲线和死亡曲线。
1.4.5 生存分析方法为了更好地分析异翅独尾草种群动态,阐明其生存规律,本文把生存分析中的4个函数引入种群生存分析中,即种群的生存率函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)、危险率函数λ(ti),这4个函数用下列公式计算[34]:
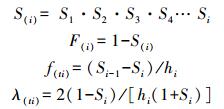
|
式中, Si为存活率;hi为龄级宽度。根据上述4个生存函数的估算值,绘制生存曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。数据的处理与作图均使用Microsoft Office Excel 2003与Origin进行。
2 结果与分析 2.1 年龄结构分析通过统计各样地内各龄级异翅独尾草植株数量,计算得a、b、c3种类型斑块的平均数,绘制不同类型样地异翅独尾草种群的年龄结构图 (图 2)。首先,从图 2中可以看出:a类型样地中异翅独尾草植株数量最少,平均仅有134株,出现某一或多个龄级植株缺失的情况,尤其是高龄级植株缺失严重;b类型各样地异翅独尾草除P4、P13、P18样地有龄级缺失外,其他样地植株在每一龄级都有一定数量的个体分布,种群年龄结构为纺锤型,呈两头小中间大的正态分布;c类型7个样地异翅独尾草总数平均达到540株,其中第Ⅰ、Ⅱ龄级的植株就占植株总数的43.85%,幼苗个体数占有绝对的优势,各样地异翅独尾草种群的年龄结构呈金字塔型,属于典型的增长型种群,说明种群能够很好地适应当地的自然条件,能够稳定维持发展下去。总体来说,a类型样地受人为干扰较强,种群规模小且Ⅸ龄级以上的植株仅有一株,种群有波动、发展不稳定,衰退风险较大;b类型样地受中等程度的干扰,虽然异翅独尾草种群表现为稳定型,但其幼苗死亡率高且储量较少,因此将其归属于稳定至衰退型种群,只要对现存的种群实行严格的保护和科学的抚育,可以在较短的时间内恢复;c类型样地中异翅独尾草种群受人为干扰较轻,种群规模大且幼苗数量丰富,种群维持有足够的后续资源,更新潜力较强,处于一种良好的增长状态。

|
| 图 2 3种类型样地中异翅独尾草种群年龄结构 Fig. 2 The age structure of three type plots of Eremurus anisopterus population |
根据静态生命表的编制要求,用每一种斑块类型中各样地平均存活数编制了异翅独尾草种群的静态生命表 (表 1)。它反映了该种群年龄动态历程中的一个特定时间段内的特点,代表了该种群生死规律的基本属性。由表 1可以看出a类型斑块的第Ⅱ龄级和b类型斑块的第Ⅰ、Ⅲ、Ⅳ、Ⅴ龄级均为低龄级的存活数小于高其一龄级的植株,使得生命表中相应龄级的dx、qx、Kx出现负值,反映了种群在该龄级段里的幼植株的缺乏程度,尤其是低龄级植株。相反,在c类型斑块中异翅独尾草种群幼苗数量较大,第Ⅰ、Ⅱ龄生命期望ex来看,种群的期望寿命在Ⅱ、Ⅲ龄级和Ⅴ、Ⅵ龄级较高,生命期望值达到3.48、3.49、3.24、3.21,随后到较高龄级开始小幅度下滑,说明异翅独尾草在中龄级阶段生存质量较高,在第Ⅴ、Ⅵ龄级的生存能力最强,并在经过种内竞争和自然筛选之后,存活下来的植株具有较强的生命期望寿命。
| 斑块类型 Plots type |
X | ax | lx | lnlx | dx | qx | Lx | Tx | ex | Kx |
| a类型 | Ⅰ | 29.500 | 1000.000 | 6.908 | 282.475 | 0.282 | 858.763 | 4031.085 | 4.031 | 0.332 |
| A type | Ⅱ | 21.167 | 717.525 | 6.576 | -152.542 | -0.213 | 793.797 | 3172.322 | 4.421 | -0.193 |
| Ⅲ | 25.667 | 870.068 | 6.769 | 310.746 | 0.357 | 714.695 | 2378.525 | 2.734 | 0.442 | |
| Ⅳ | 16.500 | 559.322 | 6.327 | 39.559 | 0.071 | 539.542 | 1663.831 | 2.975 | 0.073 | |
| Ⅴ | 15.333 | 519.763 | 6.253 | 50.847 | 0.098 | 494.339 | 1124.288 | 2.163 | 0.103 | |
| Ⅵ | 13.833 | 468.915 | 6.150 | 265.525 | 0.566 | 336.153 | 629.949 | 1.343 | 0.835 | |
| Ⅶ | 6.000 | 203.390 | 5.315 | 56.508 | 0.278 | 175.136 | 293.797 | 1.445 | 0.325 | |
| Ⅷ | 4.333 | 146.881 | 4.990 | 107.322 | 0.731 | 93.220 | 118.661 | 0.808 | 1.312 | |
| Ⅸ | 1.167 | 39.559 | 3.678 | 39.559 | 1.000 | 19.780 | 25.441 | 0.643 | ||
| Ⅹ | 0.000 | 0.000 | 0.000 | 0.000 | 5.661 | |||||
| Ⅺ | 0.000 | 0.000 | -5.661 | 2.831 | 5.661 | |||||
| Ⅻ | 0.167 | 5.661 | 1.734 | 2.831 | 2.831 | 0.500 | ||||
| b类型 | Ⅰ | 38.167 | 566.839 | 6.340 | -146.035 | -0.258 | 639.857 | 6271.063 | 11.063 | -0.229 |
| B type | Ⅱ | 48.000 | 712.875 | 6.569 | -89.109 | -0.125 | 757.429 | 5631.206 | 7.899 | -0.118 |
| Ⅲ | 54.000 | 801.984 | 6.687 | -37.129 | -0.046 | 820.549 | 4873.777 | 6.077 | -0.045 | |
| Ⅳ | 56.500 | 839.113 | 6.732 | -89.109 | -0.106 | 883.668 | 4053.228 | 4.830 | -0.101 | |
| Ⅴ | 62.500 | 928.222 | 6.833 | -71.778 | -0.077 | 964.111 | 3169.560 | 3.415 | -0.074 | |
| Ⅵ | 67.333 | 1000.000 | 6.908 | 331.680 | 0.332 | 834.160 | 2205.449 | 2.205 | 0.403 | |
| Ⅶ | 45.000 | 668.320 | 6.505 | 225.254 | 0.337 | 555.693 | 1371.289 | 2.052 | 0.411 | |
| Ⅷ | 29.833 | 443.067 | 6.094 | 133.664 | 0.302 | 376.235 | 815.596 | 1.841 | 0.359 | |
| Ⅸ | 20.833 | 309.403 | 5.735 | 173.258 | 0.560 | 222.773 | 439.361 | 1.420 | 0.821 | |
| Ⅹ | 9.167 | 136.144 | 4.914 | 34.664 | 0.255 | 118.812 | 216.588 | 1.591 | 0.294 | |
| Ⅺ | 6.833 | 101.481 | 4.620 | 54.446 | 0.537 | 74.258 | 97.775 | 0.963 | 0.769 | |
| Ⅻ | 3.167 | 47.035 | 3.851 | 23.517 | 23.517 | 0.500 | ||||
| c类型 | Ⅰ | 142.143 | 1000.000 | 6.908 | 333.671 | 0.334 | 833.164 | 3299.997 | 3.300 | 0.406 |
| C type | Ⅱ | 94.714 | 666.329 | 6.502 | 158.791 | 0.238 | 586.934 | 2466.833 | 3.702 | 0.272 |
| Ⅲ | 72.143 | 507.538 | 6.230 | 78.393 | 0.154 | 468.342 | 1879.899 | 3.704 | 0.168 | |
| Ⅳ | 61.000 | 429.145 | 6.062 | 121.610 | 0.283 | 368.340 | 1411.557 | 3.289 | 0.333 | |
| Ⅴ | 43.714 | 307.535 | 5.729 | 84.422 | 0.275 | 265.324 | 1043.217 | 3.392 | 0.321 | |
| Ⅵ | 31.714 | 223.113 | 5.408 | 29.147 | 0.131 | 208.540 | 777.893 | 3.487 | 0.140 | |
| Ⅶ | 27.571 | 193.967 | 5.268 | 39.193 | 0.202 | 174.370 | 569.353 | 2.935 | 0.226 | |
| Ⅷ | 22.000 | 154.774 | 5.042 | -28.141 | -0.182 | 168.844 | 394.983 | 2.552 | -0.167 | |
| Ⅸ | 26.000 | 182.914 | 5.209 | 103.515 | 0.566 | 131.157 | 226.138 | 1.236 | 0.835 | |
| Ⅹ | 11.286 | 79.399 | 4.374 | 41.205 | 0.519 | 58.796 | 94.982 | 1.196 | 0.732 | |
| Ⅺ | 5.429 | 38.194 | 3.643 | 21.106 | 0.553 | 27.641 | 36.185 | 0.947 | 0.804 | |
| Ⅻ | 2.429 | 17.088 | 2.838 | 8.544 | 8.544 | |||||
| x:龄级;ax:在x龄级内现有个体数;lx:在x龄级开始时标准化存活个体数 (一般转换为1000);dx:从x到x+1龄级间隔期内标准化死亡数;qx:从x到x+1龄级间隔期间死亡率;Lx:从x到x+1龄级间隔期间还存活的个体数;Tx:从x龄级到超过x龄级的个体总数;ex:进入x龄级个体的生命期望或平均期望寿命;Kx:消失率 (损失度) | ||||||||||
以该群体的轮叶数作为横坐标,分别以计算得到的标准化存活量lx和死亡率qx为纵坐标,绘制得到该独尾草种群的存活曲线和死亡曲线 (图 3)。从图 3A可以看出c类型斑块种群的存活曲线呈DeeveyⅢ型,即幼苗时死亡率很高,种群数量急剧减少,随着年龄的增加,死亡率逐渐下降,种群规模开始稳定。异翅独尾草幼年个体面临激烈的种内竞争,从而造成很高的死亡率,经过严酷的环境筛选以后,种群实现了定居,死亡率维持在较低水平,直至其生理衰退年龄,整个生活史中受人为干扰较轻。b类型斑块种群的存活曲线则表现为先上升,到第Ⅵ龄级达最大存活数后又开始下降的趋势,说明严酷的自然条件使得幼苗的死亡率很高,又遭受到放牧、土地开垦与道路、勘探等工程的人类活动影响,导致幼苗更难存活。而受到人类活动严重干扰的a类型种群已严重缺乏高龄级植株,说明人类干扰强度大且持续时间较长。

|
| 图 3 异翅独尾草种群存活曲线 (A) 与死亡曲线 (B) Fig. 3 Survival curves (A) and mortality curves (B) of Eremurus anisopterus population |
死亡率曲线能够反映种群死亡率的动态变化,图 3B为异翅独尾草的死亡曲线。a类型斑块种群的死亡率极不稳定,是由于此类型斑块受人为干扰程度强,斑块破碎程度高,种群已遭到严重破坏,植株数量急剧下降,各龄级植株参差不齐,分布不全;a类型斑块的第Ⅱ龄级和b类型斑块的第Ⅰ、Ⅲ、Ⅳ、Ⅴ龄级的死亡率qx为负值,说明该龄级植株的存活数小于高其一龄级植株的存活数,同时也说明a类型与b类型斑块种群低龄级植株较少,缺乏幼苗补充;c类型斑块中从Ⅰ— Ⅲ龄级种群的死亡率逐渐下降,第Ⅳ龄级出现一个小高峰,说明在此龄级之后存活下来的个体,随着年龄的增长,营养发育阶段有一定的筛选强度,而环境筛的作用强大,只有少数幼苗完成营养生长后能进入下一生长阶段,之后死亡率又有所下降到第Ⅷ龄级开始急剧上升。实地调查中发现,极少数异翅独尾草的最大轮叶数能达到37片,这在一定程度上说明,样地内异翅独尾草龄级越接近第Ⅻ龄级,就表示越接近其实际寿命。
2.4 生存分析根据4个生存函数估算值,以龄级为横坐标,绘制生存率曲线、累积死亡率曲线死亡密度曲线和危险率曲线得到图 4、图 5。由图 4可知a类型斑块中异翅独尾草种群的生存率曲线和累计死亡率曲线除第Ⅱ龄级外,都呈逐级下降或增加的趋势,二者互补;b类型斑块中异翅独尾草种群在第Ⅱ—Ⅴ龄级的生存率增加,相应的累计死亡率下降,Ⅴ龄级开始增加;c类型斑块中异翅独尾草种群的生存率曲线和累计死亡率曲线呈单调递减或递增的趋势,变化平缓稳定。结合死亡密度函数可以看出,死亡密度曲线基本持平,危险率曲线呈单调递增的趋势。4个生存函数曲线表明,a类型种群前、中期稳定,后期衰退;b类型种群前、中期衰退,后期稳定;c类型种群稳定增长。

|
| 图 4 异翅独尾草种群生存率 (Si) 和累计死亡率 (Fi) 曲线 Fig. 4 Survival rate (Si) and cumulative mortality rate (Fi) functional curves of Eremurus anisopterus population |

|
| 图 5 异翅独尾草种群死亡密度 (f(ti)) 和危险率 (λ(ti)) 曲线 Fig. 5 Mortality density (f(ti)) and hazard rate (λ(ti)) functional curves of Eremurus anisopterus population |
种群的年龄结构可以较直观反映种群与环境间的相互关系,同时呈现出其在群落中的作用与地位,而静态生命表则可以描述种群的存亡动态。因此,研究一个物种的种群结构和生命表特征,对深刻分析其种群的现状、动态并预测未来具有重要意义[35]。在异翅独尾草的分布区内,不同强度人为干扰的斑块中,种群的结构也有不同的表现。其中,人类干扰较强和中等程度的a类型与b类型的斑块中出现了种群幼苗缺乏、比例较低和某些龄级的植株缺失的现象,尤其是a类型斑块,在开垦放牧、人为破坏以及缺乏相应保护措施下,死亡或消失的风险较高,使得个体较难通过强烈的环境筛进入种群的更替层,种群整体上表现出一定的不稳定性。c类型种群结构表现为稳定型,虽然幼苗的死亡率较高,但其基数庞大足以维持幼苗更新。Manuel认为,以种子繁殖为主的植物种群若缺少幼苗,一般判断该种群趋于衰退型[36]。虽然一个物种的种群自身具有独特的更新方式,但气候变化、人为干扰以及种间竞争都会对特定年龄段的个体产生影响,使种群的年龄结构偏离原有形状[37],并对种群的自然更新带来不同程度的影响[38]。
对于a类型斑块中的种群,属重度干扰,种群的年龄结构波动较大,种群在未来的发展中趋于不稳定,如果干扰强度加大并缺乏保护,种群中的低龄级面临死亡的风险较高,种群的更新困难,甚至导致该种群消失。在b类型斑块中,践踏及牲畜啃食导致低龄级的个体较少,种群个体主要集中在Ⅳ—Ⅶ级间,而高龄级个体也有缺失或不足,导致种群趋于稳定衰退。因此,对异翅独尾草而言,大尺度、高强度的干扰对种群的更新产生严重制约作用,并使得种群趋向于不稳定,甚至衰退。c类型斑块位于连片沙垄内,少有干扰,种群的个体主要集中在Ⅰ—Ⅲ龄级,中等龄级植株数量适中,高龄级较少,种群属显著的增长型。
静态生命表就是在同一时间或某个调查期内,用收集到的植物样地内一个种群所有个体的年龄数据编制而成的生命表,生命表结构分析是解释种群变化的前提,根据生命表可以预测出该物种在某些特定条件下存活与繁殖的可能性,了解种群的现存状态,分析过去种群的结构与受干扰状态,预测种群未来的动态[33]。由于所调查的异翅独尾草种群属于自然生境中受不同程度人为扰动后的一个真实的种群,而且是以“空间推时间”的方法进行生命表的编制中,故在生命表中会出现死亡率为负值的情况[39],其中a类型斑块的第Ⅱ龄级和b类型斑块的第Ⅰ、Ⅲ、Ⅳ、Ⅴ龄级的死亡率qx为负值。对这种情况,Wratten和Fry认为“生命表分析中产生的一些负的dx值,这与数学假设不符,但仍能提供有用的生态学记录,即表明种群并非静止不动,而是在迅速发展或衰落之中”[33]。种群的存活曲线能够较直观地反映出种群发育过程中各个龄级个体的死亡率,从而体现出种群动态趋势,研究结果显示,异翅独尾草种群的存活曲线整体呈现为DeeveyⅢ型,幼体的死亡率较高,且种群的数量波动较大。分析结果表明前期异翅独尾草种群密度大,死亡率较高,后期竞争强度显著下降,种群趋于平稳。从个体水平来看,前期高死亡率对种群个体不利,但对于整个种群而言,这个过程由于淘汰了较弱的个体,保存了较强的个体,有利于种群的进化与繁荣。结合野外实地调查,由于历年来开荒、公路、油田工程建设的影响,以及牛、羊等对当年生、多年生的植株的毁灭性取食为种群的波动提供了较好解释,且这种波动在没有及时和足够的幼龄个体补充的前提下,导致异翅独尾草种群趋于衰退的风险增高。张文辉等指出,维持种群稳定的关键因素是要为种子萌发、幼苗的定居和生长创造适宜条件[40],一旦异翅独尾草的生存环境条件改善,则能为异翅独尾草种子的萌发和幼苗的形成、定居及其生长发育创造条件。
自然界生境中每一个斑块资源都有限,生境破碎化造成物种的部分生境丧失,种群原有生境面积减少,即生境容纳量减少,所能维持的平均物种个体数量随之降低。同时,种群扩散受到限制导致种群分布范围缩小,进而影响种群的未来发展动态。Benítez-Malvido等人[41]对亚马逊河连续生境和破碎化生境中物种多样性、幼苗生长速率的研究发现,破碎化森林中的再生乔木、藤本、草本、灌木的物种丰富度比连续生境中的乔木、藤本、草本、灌木的物种丰富度低,除了藤本外其它植物个体数比原来生境中的植物减少40%;胡世俊等人[42]通过对缙云卫矛 (Euonymus chloranthoides Yang) 的研究也发现,生境破碎导致种群大小的降低,而生境破碎后小种群不利于该物种的生殖成功,导致小种群的坐果率与幼苗比例降低;Helm等人[43]调查了爱沙尼亚石灰质草地上维管束植物对长期生境丧失和破碎化度的反应,结果发现斑块中的生境专一物种与70年前的斑块面积和空间连通度有很大的关联性,预计每个斑块中约有20种维管束植物将消失;Wu等人[44]在科尔沁破碎化沙丘生态系统的研究中指出,生境破碎化对物种丰富度、植物功能型与稀有物种的影响存在阈值,且不同功能型物种对生境破碎化的响应各异。与本研究结果一致,大部分研究结果都表明,生境破碎化限制了种群分布,降低了种群数量,甚至还可能改变种群结构,生境破碎化对物种的生存与发展具有负作用,因此,对于人类及放牧干扰较重生境中的异翅独尾草种群,应积极进行限制或禁牧及围封育林措施,减少人类干扰,发挥其有性生殖与通过地下部分无性繁殖实现自然恢复的潜力。
4 结论古尔班通古特沙漠南缘异翅独尾草种群的年龄与动态因人类干扰与生境破碎化程度的差异,呈现为不同的结构特征及变化趋势。上述4个方面的分析结果都一致反映了受人类活动影响强、生境破碎化程度高的异翅独尾草种群衰退速度较快,在未来趋于衰退的风险更大,而受人为干扰较轻、低破碎程度的种群呈稳定增长的趋势。异翅独尾草种群作为古尔班通古特沙漠短命植物层片重要的组成部分之一,对春夏之际植被稳定沙面、防沙固沙具有重要作用,而且在荒漠植被的恢复与重建以及维持生态系统的平衡中占据十分重要的生态地位。结合本文研究结果,现对异翅独尾草的保护应结合不同干扰程度、不同破碎程度生境中种群的生存现状,制定切实可行的保护、恢复策略。
| [1] | Harris L D. The Fragmented Forest: Island Biogeography Theory and the Preservation of Biotic Diversity. Chicago, IL: University of Chicago Press, 1984. |
| [2] | Saunders D A, Hobbs R J, Margules C R. Biological consequences of ecosystem fragmentation: a review. Conservation Biology, 1991, 5(1): 18–32. DOI:10.1111/cbi.1991.5.issue-1 |
| [3] | Debinski D M, Holt R D. A survey and overview of Habitat fragmentation experiments. Conservation Biology, 2001, 14(2): 342–355. |
| [4] | Fahrig L. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics, 2003, 34(1): 487–515. DOI:10.1146/annurev.ecolsys.34.011802.132419 |
| [5] | 杨芳, 贺达汉. 生境破碎化对生物多样性的影响. 生态科学, 2006, 25(6): 564–567. |
| [6] | Turner M G, Gardner R H. Quantitative Methods in Landscape Ecology. New York: Springer-Verlag, 1991. |
| [7] | 汤萃文, 张海风, 陈银萍, 姚雪玲, 王丽娟, 肖笃宁. 祁连山南坡植被景观格局及其破碎化. 生态学杂志, 2009, 28(11): 2305–2310. |
| [8] | 张育新, 马克明, 牛树奎. 异质种群动态模型:破碎化景观动态模拟的新途径. 生态学报, 2003, 23(9): 1877–1890. |
| [9] | 覃凤飞, 安树青, 卓元午, 项华均, 郑建伟, 陈兴龙. 景观破碎化对植物种群的影响. 生态学杂志, 2003, 22(3): 43–48. |
| [10] | 钱亦兵, 吴兆宁, 张立运, 赵锐锋, 王小燕, 李有民. 古尔班通古特沙漠短命植物的空间分布特征. 科学通报, 2007, 52(19): 2299–2306. |
| [11] | 刘媖心. 中国沙漠植物志 (3). 北京: 科学出版社, 1985: 223–225. |
| [12] | 崔乃然, 崔大方, 刘国钧, 毛祖美, 冷巧珍, 李学禹, 阎平. 新疆植物志 (6). 乌鲁木齐: 新疆科技卫生出版社, 1996: 480–482. |
| [13] | 王雪芹, 蒋进, 雷加强, 张伟民, 钱亦兵. 古尔班通古特沙漠短命植物分布及其沙面稳定意义. 地理学报, 2003, 58(4): 598–605. |
| [14] | 张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点. 生态学报, 2002, 22(11): 1923–1932. |
| [15] | 马淼, 范俊峰, 李静. 类短命植物异翅独尾草的传粉特性. 植物生态学报, 2006, 30(6): 1012–1017. |
| [16] | 王海娟, 段呈, 安静, 吴玲, 张霞, 王绍明. 荒漠斑块生境中异翅独尾草种子形态特征研究. 种子, 2015, 34(5): 17–21. |
| [17] | Xiao J Q, Eshbakova K A, Aisa H A. Chemical constituents of Eremurus anisopterus. Chemistry of Natural Compounds, 2014, 50(4): 730–731. DOI:10.1007/s10600-014-1065-y |
| [18] | Hu C F, Kong Q J, Yang D Y, Pan Y J. Isolation and structural characterization of a novel galactomannan from Eremurus anisopterus (Ker. et Kir) Regel roots. Carbohydrate Polymers, 2011, 84(1): 402–406. DOI:10.1016/j.carbpol.2010.11.054 |
| [19] | 吴玲, 魏凌基, 马淼, 邱爱军. 异翅独尾草的核型分析. 石河子大学学报:自然科学版, 2004, 22(5): 417–418. |
| [20] | Harper J L. Population Biology of Plants. London: Academic Press, 1977: 599–643. |
| [21] | 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. |
| [22] | 韩路, 席琳乔, 王家强, 王海珍, 宇振荣. 塔里木河上游灰胡杨种群生活史特征与空间分布格局. 生态学报, 2013, 33(19): 6181–6190. |
| [23] | 李先琨, 苏宗明, 向悟生, 宁世江, 唐润琴, 欧祖兰, 李瑞棠. 濒危植物元宝山冷杉种群结构与分布格局. 生态学报, 2002, 22(12): 2246–2253. |
| [24] | 张文辉, 卢志军, 李景侠, 刘国彬. 秦岭北坡栓皮栎种群动态的研究. 应用生态学报, 2003, 14(9): 1427–1432. |
| [25] | 刘仲健, 陈利君, 饶文辉, 李利强, 张玉婷. 长瓣杓兰 (Cypripedium lentiginosum) 种群数量动态与生殖行为的相关性. 生态学报, 2008, 28(1): 111–121. |
| [26] | 解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家. 荒漠绿洲过渡带沙拐枣种群结构及动态特征. 生态学报, 2014, 34(15): 4272–4279. |
| [27] | 李豪, 张钦弟, 苗艳明, 杨兆静, 毕润成. 历山自然保护区秃山白树天然种群生命表. 生态学杂志, 2013, 32(1): 52–58. |
| [28] | 季方, 叶玮, 魏文寿. 古尔班通古特沙漠固定与半固定沙丘成因初探. 干旱区地理, 2000, 23(1): 32–36. |
| [29] | 周宏飞, 李彦, 汤英, 周宝佳, 徐宏伟. 古尔班通古特沙漠的积雪及雪融水储存特征. 干旱区研究, 2009, 26(3): 312–317. |
| [30] | 张治国. 青岛百合 (Lilium tsingtauense) 复合种群研究[D]. 上海: 华东师范大学, 2002. |
| [31] | 张文辉, 李景侠, 李红, 刘祥君. 独叶草种群年龄结构及动态分析. 应用生态学报, 2004, 15(4): 561–565. |
| [32] | Harcombe P A. Tree life tables. Bioscience, 1987, 37(8): 557–568. DOI:10.2307/1310666 |
| [33] | Wratten S D, Fry G L A. Field and Laboratory Exercises in Ecology. London: Edward Arnold, 1980. |
| [34] | 杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153–158. |
| [35] | Svensson J S, Jeglum J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline. Forest Ecology and Management, 2001, 151(1-3): 67–79. DOI:10.1016/S0378-1127(00)00697-6 |
| [36] | Molles M C. Ecology: Concept and Application. Beijing: Science Press, 2000: 162–266. |
| [37] | Agren J, Zackrisson O. Age and size structure of Pinus sylvestris populations on mires in central and northern Sweden. Journal of Ecology, 1990, 78(4): 1049–1062. DOI:10.2307/2260951 |
| [38] | Dang H S, Jiang M X, Zhang Y J, Dang G D, Zhang Q F. Dendroecological study of a subalpine fir (Abies fargesii) forest in the Qinling Mountains, China. Plant Ecology, 2009, 201(1): 67–75. DOI:10.1007/s11258-008-9491-1 |
| [39] | Begon M, Mortimer M, Thompson D J. Population Ecology: A Unified Study of Animals and Plants. London: Black-well Scientific Publication, 1981. |
| [40] | 张文辉, 祖元刚, 刘国彬. 十种濒危植物的种群生态学特征及致危因素分析. 生态学报, 2002, 22(9): 1512–1520. |
| [41] | Benítez-Malvido J, Martínez-Ramos M. Impact of forest fragmentation on understory plant species richness in Amazonia. Conservation Biology, 2003, 17(2): 389–400. DOI:10.1046/j.1523-1739.2003.01120.x |
| [42] | 胡世俊, 何平, 张春平, 张益锋. 破碎生境中种群大小对缙云卫矛生殖成功的影响. 生态学杂志, 2013, 32(4): 821–824. |
| [43] | Helm A, Hanski I, Pärtel M. Slow response of plant species richness to habitat loss and fragmentation. Ecology Letters, 2006, 9(1): 72–77. |
| [44] | Wu J, Liu Z M, Qian J Q. Non-linear effect of habitat fragmentation on plant diversity: evidence from a sand dune field in a desertified grassland in northeastern China. Ecological Engineering, 2013, 54: 90–96. DOI:10.1016/j.ecoleng.2013.01.020 |