 2017, Vol. 37
2017, Vol. 37文章信息
- 周泓杨 , 张健 , 张丹桔 , 张捷 , 魏大平 , 赵燕波 , 赵波 , 李川北
- ZHOU Hongyang , ZHANG Jian , ZHANG Danju , ZHANG Jie , WEI Daping , ZHAO Yanbo , ZHAO Bo , LI Chuanbei .
- 不同郁闭度控制下马尾松(Pinus massoniana)人工林土壤动物群落特征
- Community characteristics of soil fauna for different canopy density of a Pinus massoniana plantation
- 生态学报. 2017, 37(6): 1939-1955
- Acta Ecologica Sinica. 2017, 37(6): 1939-1955
- http://dx.doi.org/10.5846/stxb201510232148
-
文章历史
- 收稿日期: 2015-10-23
- 网络出版日期: 2016-08-02






2. 四川农业大学林学院, 成都 611130
2. Forestry College of Sichuan Agricultural University, Chengdu 611130, China
土壤动物作为森林生态系统的重要组成部分, 与地下植物根系、微生物等一起构成完整稳定的食物网络, 在人工林土壤活性及养分循环方面发挥了积极作用[1]。土壤动物取食、掘穴等活动直接促进枯落物分解和粉碎, 增加了土壤有机元素的有效性, 促进微生物多样性和土壤微环境异质性, 能够加速养分循环[2], 影响土壤有机质稳定性和碳、氮平衡[3]。土壤动物与土壤微生物及地上部分的相互关系有利于森林生态系统功能的发挥[4]。因此, 研究森林生态系统土壤动物群落结构和功能具有十分重要的意义。目前, 关于人工林经营与生态系统中土壤动物群落结构和功能的研究已有大量报道, 如Andrey[5]等研究了云杉纯林经过4个演替阶段发展成混交林对土壤甲螨群落结构的影响。张良辉[6]研究了川西亚高山米亚罗林区云杉人工林不同大小林窗以及不同强度抚育间伐调控下植物和土壤动物多样性。而关于人工林不同郁闭度的生态功能研究还相对较少[7]。
马尾松 (Pinus massoniana) 作为我国南方和长江流域低山丘陵区主要造林树种, 面积超过200万hm2, 在人工林中占有很大比例[8]。然而, 马尾松人工林林下植被结构单一、生物多样性降低[9];马尾松凋落物腐化分解产生的有机酸等导致土壤酸化, 降低了部分养分元素有效性[10];连栽马尾松林使土壤速效氮、速效磷、代换性钙镁离子含量明显下降, 土壤酶类活性及微生物区系随之相应变化, 从而影响整个马尾松林下土壤代谢活动[11]。因此, 采取科学的抚育措施加强马尾松人工林经营的要求非常迫切。林分郁闭度作为反映森林结构和环境状态的重要因子, 在林分结构构建与调控过程中为确定抚育措施和抚育强度提供参考[12]。通过郁闭度调节林内光照和水分, 减小森林生态系统中光合有效辐射、土壤温度、土壤含水量、空气湿度和温度的极端分布, 为林下植被发育和生物活动创造稳定良好的小生境[13]。土壤动物对生存环境的感受与适应, 决定了郁闭度变化也显然会影响到土壤生态系统中土壤动物的群落结构组成。而目前关于不同郁闭度对土壤生态系统尤其是对土壤动物的研究还未见报道, 难以完整全面地解释郁闭度生态效应及其响应机制。
土壤动物群落受气温和降雨的季节变化, 在亚热带针叶林中通常于秋季大量发生[14]。为此, 本文选择四川宜宾11年生马尾松人工林作为研究对象, 通过抚育间伐结合修枝控制形成5个郁闭度梯度 (0.5、0.6、0.7、0.8、0.9), 于2014年10月在不同郁闭度下马尾松人工林进行土壤动物群落组成、结构及多样性研究, 以期为马尾松人工林的科学经营和管理提供依据。
1 研究区概况研究区位于四川省高县来复镇 (28°11′—28°47′ N, 104°21′—104°48′ E), 地处四川盆地南缘。该地属中亚热带湿润季风气候, 气候温和, 雨量充沛。年平均气温18.1 ℃, 最高气温 (7月)36.8 ℃, 最低气温 (1月)7.8 ℃, 年均降雨量1 021 mm。相对湿度81%, 年日照时数1 148 h。地貌以低山丘陵为主, 海拔412—567 m。土壤类型为发育在砂岩上的山地黄壤。植被绝大部分为原始植被受破坏后的次生植被, 马尾松人工林林分结构简单, 初植密度较大。常见林下灌木有枹栎 (Quercus serrate)、黄牛奶 (Symplocos laurina)、梨叶悬钩子 (Rubus pirifolius) 等, 草本层有淡竹叶 (Lophatherum gracile), 金星蕨 (Parathelypteris glanduligera)、芒萁 (Dicranopteris dichotoma)、山菅兰 (Dianella ensifolia) 等。
2 材料与方法 2.1 样地设置本次实验样地位于高县来复镇森林经营所毛颠坳Ⅴ1林班大窝场。2013年9月, 在已有马尾松人工林 (2.84 hm2, 郁闭度0.9, 林龄11 a, 栽植密度1.5 m×1.5 m) 的基础上通过间伐、修枝形成郁闭度为0.5、0.6、0.7、0.8、0.9梯度系列样地。郁闭度采用数字式植物冠层分析仪 (CI-110) 测定。每个处理林分面积约为20 m×20 m。每个郁闭度设置3个重复样地。每个样地间隔10 m, 样地边界离林缘不小于10 m。样地基本情况见表 1, 其中土壤性质的测定于2014年10月与土壤动物样品采集同步进行, 以“品”字形采样法采集0—15 cm层土壤混合样品并测定。
| 郁闭度 Canopy density |
海拔/m Altitude |
坡度/° Gradient |
坡位 Slope position |
坡向 Aspect |
含水量/% Moisture content |
土壤pH | 容重/(g/cm3) Bulk density |
有机碳/(g/kg) Soil organic carbon |
全氮/(g/kg) Total nitrogen |
全磷 (/g/kg) Total phosphorus |
| 0.5 | 488.57 | 30.33 | 中坡 | 西南 | 22.98±1.08a | 3.98±0.03b | 1.37 ±0.04 | 16.07 ±0.88a | 1.03 ±0.04a | 0.20 ±0.00ac |
| 0.6 | 456.67 | 29.00 | 中坡 | 东南 | 20.23±1.10ab | 4.00±0.01b | 1.36 ±0.06 | 12.64 ±0.92b | 0.73±0.03c | 0.17 ±0.02bc |
| 0.7 | 460.17 | 29.00 | 中坡 | 东南 | 21.90±1.23ab | 3.97±0.01bc | 1.36±0.05 | 15.67 ±1.33ab | 0.94 ±0.04ab | 0.24 ±0.02a |
| 0.8 | 462.45 | 30.67 | 中坡 | 东南 | 19.64±0.29b | 4.08±0.03a | 1.39 ±0.04 | 12.56 ±1.23b | 0.77±0.03c | 0.15 ±0.01b |
| 0.9 | 461.57 | 28.33 | 中坡 | 东南 | 20.00±0.56b | 3.91±0.01c | 1.31 ±0.05 | 17.92 ±0.46a | 0.91 ±0.04b | 0.15 ±0.01b |
| 同列不同小写字母表示差异显著 (P < 0.05) | ||||||||||
2014年10月, 在样方内以“五点法”进行采样, 样点选择较为平坦、人为活动少且避开斜坡地、洼地、岩石、倒木和大树根。每个采样点分为枯落物层、0—5 cm、5—10 cm 3个不同层次取样。土壤动物按体形、习性及其在食物分解过程中的作用分为大型和中小型两类[15-16]。其中, 大型土壤动物主要是指野外调查中肉眼可见的在浅层土壤活动的大型种类, 采用手捡法, 采样面积为50 cm×50 cm (0.25 m2), 将收集到的土壤动物放入盛有浓度为75%的酒精容器中杀死后带回实验室在解剖镜下分类计数;中小型土壤动物包括用湿漏斗法 (Baermann法) 收集的小型湿生土壤动物和用干漏斗法 (Tullgren法) 收集的中小型节肢动物。首先在采样点收集10 cm×10 cm (0.01 m2) 枯落物带回实验室用Tullgren法分离土壤动物, 再用环刀 (100 mL) 自下往上顺次取土, 每个土层取两个土样, 带回实验室分别用Tullgren干漏斗和Baermann湿漏斗分离。由于湿生土壤动物平均密度较大, 用于分离湿生的土壤样品采用“四分法”取四分之一 (25 mL) 进行分离[17]。
土壤动物的分离在烘虫箱中进行, 控制烘虫箱温度在35—50 ℃, 烘虫时间为48 h, 其中湿生土壤动物在分离4 h后观察一次, 以防线虫自溶。干漏斗法分离出的土壤动物用盛有75%浓度的酒精收集于培养皿中, 湿生土壤动物则用清水。对分离出的土壤动物进行分类鉴定并计数。
土壤动物的分类鉴定, 将捕获所得的土壤动物置于双目解剖镜 (Leica, EZ4HD) 下, 主要参考《中国土壤动物检索图鉴》、《中国亚热带土壤动物》、《昆虫分类检索》和《幼虫分类学》进行分类鉴定。受分类限制, 采用大类进行分类。
2.3 数据处理与分析土壤动物类群数量等级划分依据:个体数量占捕获总量的10.0%以上者为优势类群 (+++), 占1.0%—10.0%者为常见类群 (++), 不足1.0%者为稀有类群 (+)。
群落多样性分析:土壤动物多样性特征主要采用Shannon-Wiener多样性指数 (H′)、Margalef丰富度指数 (D)、Pielou均匀度指数 (J) 和Simpson优势度指数 (C) 来计算。
Simpson优势度指数 (C):
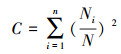
|
Shannon-Wiener多样性指数 (H′):
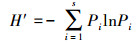
|
Pielou均匀度指数 (E):

|
Margalef丰富度指数 (D):
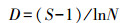
|
式中,Ni为第i类群的个体数;N为群落中所有类群的个体数;Pi=Ni/N, S为类群数。
群落相似性分析:采用Jaccard相似性指数
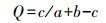
|
式中, a为A样地全部类群数, b为B样地全部类群数, c为A、B样地共有的类群数。主要用于反映林分之间土壤动物类群上的相似程度。
数据的整理与分析主要采用Excel 2010和SPSS 22.0, 采用Origin 8.1绘制图形。利用单因素方差分析 (one-way ANOVA) 比较不同郁闭度间土壤动物群落的差异性, 用最小显著差异法 (LSD) 进行两两比较。用Canoco for Windows 4.5对土壤动物和主要土壤环境因子进行CCA分析。
3 结果与分析 3.1 土壤动物群落组成本次实验共捕获土壤动物8 860只, 隶属于4门11纲25目111科。其中大型土壤动物589只, 隶属于2门8纲19目53科 (附表 1);以蜚蠊科、疣科、康叭科为优势类群, 占大型土壤动物捕获量的48.30%;以蚁科、螱科、步甲科、鞘翅目幼虫等为常见类群, 占大型土壤动物捕获量的39.12%;其余12.59%为稀有类群。中小型土壤动物共捕获8 271只, 隶属于4门7纲18目75科 (附表 2);其中优势类群为等节科和丽甲螨科, 占中小型土壤动物捕获量的53.54%;常见类群为懒甲螨科、矮蒲螨科、大翼甲螨科、美绥螨科、囊螨科、跳虫、线虫等, 占中小型土壤动物捕获量的36.27%;其余10.19%为稀有类群。
| 类群Taxa | 0.9 | 0.8 | 0.7 | 0.6 | 0.5 | 合计 | 多度 Abundance |
功能群 Functional group |
|||||||||||
| 个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
||||||||
| 大蚓类Megadrile oligochaetes | 1 | 0.70 | 9 | 9.00 | — | — | 3 | 2.27 | 2 | 1.89 | 15 | 2.55 | ++ | S | |||||
| 螲蟷科Ctenizidae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | Pr | |||||
| 盗蛛科Pisauridae | — | — | 1 | 1.00 | 1 | 0.93 | 2 | 1.52 | — | — | 4 | 0.68 | + | Pr | |||||
| 狼栉蛛科Zoridae | — | — | 1 | 1.00 | — | — | — | — | 1 | 0.94 | 2 | 0.34 | + | Pr | |||||
| 类球蛛科Nesticidae | — | — | — | — | — | — | 2 | 1.52 | — | — | 2 | 0.34 | + | Pr | |||||
| 漏斗蛛科Agelenidae | — | — | — | — | — | — | 1 | 0.76 | — | — | 1 | 0.17 | + | Pr | |||||
| 卵形蛛科Oonopidae | 4 | 2.80 | — | — | — | — | 1 | 0.76 | — | — | 5 | 0.85 | + | Pr | |||||
| 平腹蛛科Gnaphosidae | 1 | 0.70 | — | — | 1 | 0.93 | — | — | 3 | 2.83 | 5 | 0.85 | + | Pr | |||||
| 球蛛科Theridiidae | — | — | 1 | 1.00 | — | — | — | — | — | — | 1 | 0.17 | + | Pr | |||||
| 跳蛛科Salticidae | 3 | 2.10 | — | — | — | — | 3 | 2.27 | 4 | 3.77 | 10 | 1.70 | ++ | Pr | |||||
| 逍遥蛛科Philodromidae | — | — | 1 | 1.00 | — | — | 3 | 2.27 | 1 | 0.94 | 5 | 0.85 | + | Pr | |||||
| 蟹蛛科Thomisidae | 3 | 2.10 | 1 | 1.00 | 8 | 7.48 | 3 | 2.27 | 2 | 1.89 | 17 | 2.89 | ++ | Pr | |||||
| 园蛛科Araneidae | — | — | 2 | 2.00 | 3 | 2.80 | 2 | 1.52 | 2 | 1.89 | 9 | 1.53 | ++ | Pr | |||||
| 栅蛛科Hahniidae | — | — | — | — | — | — | 1 | 0.76 | — | — | 1 | 0.17 | + | Pr | |||||
| 栉足蛛科Ctenidae | — | — | — | — | — | — | 1 | 0.76 | 2 | 1.89 | 3 | 0.51 | + | Pr | |||||
| 美绥螨科Ameroseiidae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | S | |||||
| 寄螨科Parasitidae | 2 | 1.40 | — | — | — | — | — | — | — | — | 2 | 0.34 | + | S | |||||
| 绒螨科Trombidiidae | — | — | 1 | 1.00 | — | — | 2 | 1.52 | — | — | 3 | 0.51 | + | S | |||||
| 潮虫科Oniscidae | — | — | — | — | — | — | — | — | 2 | 1.89 | 2 | 0.34 | + | D | |||||
| 奇马陆科Paradoxosomatidae | — | — | 6 | 6.00 | 1 | 0.93 | 3 | 2.27 | — | — | 10 | 1.70 | ++ | Ph | |||||
| 奥地蜈蚣科Oryidae | 8 | 5.59 | 2 | 2.00 | 1 | 0.93 | 5 | 3.79 | — | — | 16 | 2.72 | ++ | Pr | |||||
| 跳虫科Poduridae | — | — | 1 | 1.00 | 1 | 0.93 | — | — | 1 | 0.94 | 3 | 0.51 | + | O | |||||
| 疣科Neanuridae | 26 | 18.18 | 8 | 8.00 | 14 | 13.08 | 10 | 7.58 | 24 | 22.64 | 82 | 13.95 | +++ | O | |||||
| 副铗叭科Parajapygidae | 1 | 0.70 | — | — | 1 | 0.93 | 2 | 1.52 | 2 | 1.89 | 6 | 1.02 | ++ | O | |||||
| 铗叭科Japygidae | — | — | 1 | 1.00 | — | — | — | — | — | — | 1 | 0.17 | + | O | |||||
| 康叭科Campodeidae | 29 | 20.28 | 9 | 9.00 | 6 | 5.61 | 19 | 14.39 | 7 | 6.60 | 70 | 11.90 | +++ | O | |||||
| 蜚蠊科Blattidae | 30 | 20.98 | 30 | 3— | 17 | 15.89 | 29 | 21.97 | 26 | 24.53 | 132 | 22.45 | +++ | O | |||||
| 鼻螱科Rhinotermitidae | 2 | 1.40 | — | — | — | — | — | — | — | — | 2 | 0.34 | + | D | |||||
| 螱科Termitidae | — | — | 1 | 1.00 | 20 | 18.69 | 1 | 0.76 | 3 | 2.83 | 25 | 4.25 | ++ | D | |||||
| 蟋蟀科Gorillidae | — | — | 1 | 1.00 | — | — | 1 | 0.76 | — | — | 2 | 0.34 | + | Ph | |||||
| 蚱总科Tetrigoidae | — | — | — | — | 1 | 0.93 | — | — | — | — | 1 | 0.17 | + | Ph | |||||
| 螯螋科Chelisochidae | — | — | — | — | — | — | — | — | 3 | 2.83 | 3 | 0.51 | + | Ph | |||||
| 肥螋科Anisolabis | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | O | |||||
| 臀螋科Apachyidae | 2 | 1.40 | — | — | — | — | — | — | — | — | 2 | 0.34 | + | O | |||||
| 红蝽科Pyrrhocoridae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | Ph | |||||
| 奇蝽科Enicocephalidae | — | — | — | — | — | — | 1 | 0.76 | — | — | 1 | 0.17 | + | Ph | |||||
| 驼蝽科Mierophysidae | — | — | 2 | 2.00 | 1 | 0.93 | 1 | 0.76 | — | — | 4 | 0.68 | + | Ph | |||||
| 长蝽科Lygeaidae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | Ph | |||||
| 啮科Psocidae | — | — | — | — | — | — | 1 | 0.76 | — | — | 1 | 0.17 | + | O | |||||
| 鼠啮科Myopsocidae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | O | |||||
| 沼啮科Elipsocidae | 1 | 0.70 | 1 | 1.00 | 1 | 0.93 | — | — | — | — | 3 | 0.51 | + | O | |||||
| 步甲科Carabidae | 2 | 1.40 | 7 | 7.00 | 11 | 10.28 | 6 | 4.55 | 5 | 4.72 | 31 | 5.27 | ++ | Pr | |||||
| 拟步甲科Tenebrionidae | — | — | — | — | — | — | 1 | 0.76 | 2 | 1.89 | 3 | 0.51 | + | Ph | |||||
| 锹甲科Lucanidea | — | — | — | — | 1 | 0.93 | — | — | — | — | 1 | 0.17 | + | Ph | |||||
| 叶甲科Chrysomelidae | 1 | 0.70 | — | — | — | — | — | — | — | — | 1 | 0.17 | + | Ph | |||||
| 蚁甲科Pselaphidae | 4 | 2.80 | — | — | — | — | 2 | 1.52 | — | — | 6 | 1.02 | ++ | D | |||||
| 隐翅甲科Staphylinidae | 2 | 1.40 | 2 | 2.00 | 3 | 2.80 | 2 | 1.52 | 3 | 2.83 | 12 | 2.04 | ++ | S | |||||
| 长角象甲科Anthribidae | 1 | 0.70 | — | — | 2 | 1.87 | — | — | — | — | 3 | 0.51 | + | Ph | |||||
| 长角沼甲科Ptilodactylidae | — | — | — | — | — | — | 1 | 0.76 | — | — | 1 | 0.17 | + | Ph | |||||
| 粪金龟科Geotrupidae | — | — | — | — | — | — | 1 | 0.07 | — | — | 1 | 0.01 | + | Co | |||||
| 鞘翅目幼虫Coleoptera larvae | 1 | 0.70 | 5 | 5.00 | 5 | 4.67 | 10 | 7.58 | 7 | 6.60 | 28 | 4.76 | ++ | Ph | |||||
| 双翅目幼虫Diptera larvae | 5 | 3.50 | 1 | 1.00 | 7 | 6.54 | 1 | 0.76 | 3 | 2.83 | 17 | 2.89 | ++ | S | |||||
| 蚁科Formicidae | 8 | 5.59 | 6 | 6.00 | 1 | 0.93 | 12 | 9.09 | 1 | 0.94 | 28 | 4.76 | ++ | O | |||||
| 总个体数Total individuals | 143 | 100 | 107 | 133 | 106 | 589 | |||||||||||||
| 总类群数Total group number | 28 | 24 | 22 | 32 | 22 | 53 | |||||||||||||
| —表示无或未采集到, +++为优势类群, ++为常见类群, +为稀有类群; Ph:植食性Phytophage;F:菌食性Fungivorous forms;Pr:捕食性Predators; S:腐食性Saprozoic;O:杂食性Omnivores;D:枯食性Debris-feeder′ s;Co:粪食性Coprophages | |||||||||||||||||||
| 类群Taxa | 0.9 | 0.8 | 0.7 | 0.6 | 0.5 | 合计 | 多度 Abundance |
功能群 Functional group |
|||||||||||
| 个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
个体数 Individual/个 |
百分比 Percentage/% |
||||||||
| 轮虫Rotifera | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | Pr | |||||
| 线虫Nemata | 89 | 9.46 | 14 | 0.64 | 15 | 0.61 | 25 | 1.66 | 10 | 0.86 | 153 | 1.85 | ++ | O | |||||
| 线蚓科Enchyiraeidae | 43 | 4.57 | 12 | 0.55 | 4 | 0.16 | 1 | 0.07 | 4 | 0.34 | 64 | 0.77 | + | Ph | |||||
| 地蛛科Atypidae | — | — | — | — | — | — | 1 | 0.07 | — | — | 1 | 0.01 | + | Pr | |||||
| 卷叶蛛科Dictynidae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | Pr | |||||
| 卵形蛛科Oonopidae | — | — | 2 | 0.09 | — | — | 1 | 0.07 | — | — | 3 | 0.04 | + | Pr | |||||
| 跳蛛科Salticidae | — | — | 2 | 0.09 | 2 | 0.08 | — | — | — | — | 4 | 0.05 | + | Pr | |||||
| 幽灵蛛科Pholcidae | 1 | 0.11 | — | — | 4 | 0.16 | 2 | 0.13 | — | — | 7 | 0.08 | + | Pr | |||||
| 栅蛛科Hahniidae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | Pr | |||||
| 土伪蝎科Chthoniidae | 3 | 0.32 | 4 | 0.18 | 8 | 0.32 | 1 | 0.07 | 2 | 0.17 | 18 | 0.22 | + | Pr | |||||
| 美绥螨科Ameroseiidae | 26 | 2.76 | 51 | 2.34 | 31 | 1.25 | 48 | 3.19 | 11 | 0.94 | 167 | 2.02 | ++ | S | |||||
| 表刻螨科Epicridae | 10 | 1.06 | 10 | 0.46 | 5 | 0.20 | 1 | 0.07 | — | — | 26 | 0.31 | + | Ph | |||||
| 虫穴螨科Zerconidae | 4 | 0.43 | — | — | — | — | — | — | — | — | 4 | 0.05 | + | S | |||||
| 厚厉螨科Pachylaelapidae | — | — | — | — | 3 | 0.12 | 1 | 0.07 | — | — | 4 | 0.05 | + | Ph | |||||
| 寄螨科Parasitidae | 27 | 2.87 | 33 | 1.51 | 19 | 0.77 | 10 | 0.66 | 5 | 0.43 | 94 | 1.14 | ++ | S | |||||
| 巨螯螨科Macrochelidae | — | — | 2 | 0.09 | — | — | — | — | — | — | 2 | 0.02 | + | S | |||||
| 厉螨科Lacclapidae | — | — | — | — | 3 | 0.12 | — | — | — | — | 3 | 0.04 | + | Pr | |||||
| 囊螨科Ascidae | 22 | 2.34 | 53 | 2.43 | 93 | 3.76 | 65 | 4.32 | 10 | 0.86 | 243 | 2.94 | ++ | S | |||||
| 派盾螨科Parholaspididae | 1 | 0.11 | 5 | 0.23 | 18 | 0.73 | 10 | 0.66 | 5 | 0.43 | 39 | 0.47 | + | S | |||||
| 绥螨科Sejidae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | S | |||||
| 维螨科Veigaiidae | 1 | 0.11 | 1 | 0.05 | 10 | 0.40 | — | — | — | — | 12 | 0.15 | + | S | |||||
| 尾足螨科Uropodidae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | S | |||||
| 真伊螨科Eriphididae | — | — | — | — | 1 | 0.04 | — | — | — | — | 1 | 0.01 | + | S | |||||
| 足角螨科Podocinidae | 4 | 0.43 | 7 | 0.32 | 3 | 0.12 | — | — | 2 | 0.17 | 16 | 0.19 | + | S | |||||
| 矮蒲螨科Pygmephoridae | 16 | 1.70 | 48 | 2.20 | 72 | 2.91 | 27 | 1.79 | 31 | 2.66 | 194 | 2.35 | ++ | S | |||||
| 跗线螨科Tarsonmidae | 4 | 0.43 | 4 | 0.18 | 1 | 0.04 | 6 | 0.40 | — | — | 15 | 0.18 | + | S | |||||
| 绒螨科Trombidiidae | 1 | 0.11 | 1 | 0.05 | 1 | 0.04 | — | — | — | — | 3 | 0.04 | + | S | |||||
| 肉食螨科Cheylrtidae | 7 | 0.74 | 4 | 0.18 | 3 | 0.12 | 25 | 1.66 | 3 | 0.26 | 42 | 0.51 | + | S | |||||
| 微离螨科Microdispidae | — | — | 1 | 0.05 | — | — | 3 | 0.20 | — | — | 4 | 0.05 | + | S | |||||
| 吸螨科Bdellidae | 16 | 1.70 | 26 | 1.19 | 19 | 0.77 | 27 | 1.79 | 12 | 1.03 | 100 | 1.21 | ++ | S | |||||
| 叶螨科Tetranychidae | 2 | 0.21 | — | — | — | — | — | — | — | — | 2 | 0.02 | + | S | |||||
| 隐颚螨科Cryptognathidae | 2 | 0.21 | 4 | 0.18 | 12 | 0.48 | 1 | 0.07 | — | — | 19 | 0.23 | + | S | |||||
| 奥甲螨Oppiidae | 2 | 0.21 | 40 | 1.83 | 63 | 2.54 | 32 | 2.13 | 22 | 1.89 | 159 | 1.92 | ++ | S | |||||
| 步甲螨Carabodidae | 1 | 0.11 | 11 | 0.50 | 3 | 0.12 | 30 | 1.99 | 47 | 4.03 | 92 | 1.11 | ++ | O | |||||
| 大翼甲螨科Galumnidae | 43 | 4.57 | 79 | 3.62 | 39 | 1.58 | 27 | 1.79 | 37 | 3.17 | 225 | 2.72 | ++ | S | |||||
| 耳头甲螨科Otocepheidae | — | — | — | — | 11 | 0.44 | 1 | 0.07 | 2 | 0.17 | 14 | 0.17 | + | S | |||||
| 缝甲螨科Hypochthoniidae | — | — | 2 | 0.09 | 10 | 0.40 | 5 | 0.33 | — | — | 17 | 0.21 | + | S | |||||
| 古甲螨科Palaeacaridae | 8 | 0.85 | 2 | 0.09 | — | — | — | — | — | — | 10 | 0.12 | + | S | |||||
| 尖棱甲螨科Ceratozetidae | — | — | 4 | 0.18 | 5 | 0.20 | — | — | — | — | 9 | 0.11 | + | S | |||||
| 懒甲螨科Nothridae | 128 | 13.60 | 96 | 4.40 | 204 | 8.24 | 88 | 5.85 | 87 | 7.46 | 603 | 7.29 | ++ | S | |||||
| 丽甲螨科Liacaridae | 46 | 4.89 | 191 | 8.75 | 464 | 18.74 | 110 | 7.31 | 96 | 8.23 | 907 | 10.96 | +++ | S | |||||
| 罗甲螨科Lohmannidae | 10 | 1.06 | 4 | 0.18 | 4 | 0.16 | 5 | 0.33 | 1 | 0.09 | 24 | 0.29 | + | S | |||||
| 裸珠甲螨科gymnodamaeoid mites | — | — | 1 | 0.05 | — | — | 1 | 0.07 | — | — | 2 | 0.02 | + | S | |||||
| 若甲螨科Oribatulidae | 14 | 1.49 | 11 | 0.50 | 16 | 0.65 | 7 | 0.47 | 44 | 3.77 | 92 | 1.11 | ++ | S | |||||
| 沙甲螨科Eremulidae | 1 | 0.11 | 20 | 0.92 | 14 | 0.57 | 18 | 1.20 | — | — | 53 | 0.64 | + | S | |||||
| 鲜甲螨科Cepheidae | — | — | 1 | 0.05 | — | — | 9 | 0.60 | — | — | 10 | 0.12 | + | S | |||||
| 小棱甲螨科Microzetidae | — | — | 11 | 0.50 | — | — | — | — | — | — | 11 | 0.13 | + | S | |||||
| 折甲螨科ptychoid oribatid mites | 8 | 0.85 | 7 | 0.32 | 27 | 1.09 | 18 | 1.20 | 8 | 0.69 | 68 | 0.82 | + | S | |||||
| 珠甲螨科Damaeidae | 2 | 0.21 | 6 | 0.27 | 6 | 0.24 | 2 | 0.13 | 4 | 0.34 | 20 | 0.24 | + | S | |||||
| 地蜈蚣科Geophilidae | — | — | — | — | 1 | 0.04 | — | — | — | — | 1 | 0.01 | + | Pr | |||||
| 原尾纲Protura | — | — | — | — | 2 | 0.08 | — | — | 4 | 0.34 | 6 | 0.07 | + | F | |||||
| 等节科Isotomidae | 170 | 18.07 | 933 | 42.74 | 1000 | 40.39 | 775 | 51.50 | 644 | 55.18 | 3522 | 42.58 | +++ | F | |||||
| 棘科Onychiuridae | 19 | 2.02 | 10 | 0.46 | 15 | 0.61 | 16 | 1.06 | 13 | 1.11 | 73 | 0.88 | + | Ph | |||||
| 球角科Hypogastruridae | 98 | 10.41 | 20 | 0.92 | — | — | 1 | 0.07 | — | — | 119 | 1.44 | ++ | F | |||||
| 跳虫科Poduridae | 15 | 1.59 | 55 | 2.52 | 80 | 3.23 | 36 | 2.39 | 25 | 2.14 | 211 | 2.55 | ++ | O | |||||
| 疣科Neanuridae | 4 | 0.43 | 12 | 0.55 | 2 | 0.08 | 5 | 0.33 | — | — | 23 | 0.28 | + | O | |||||
| 圆科Sminthurinae | 2 | 0.21 | 21 | 0.96 | 27 | 1.09 | 12 | 0.80 | 9 | 0.77 | 71 | 0.86 | + | Ph | |||||
| 长角长科Orchesellidae | 18 | 1.91 | 56 | 2.57 | 3 | 0.12 | 4 | 0.27 | — | — | 81 | 0.98 | + | F | |||||
| 康叭科Campodeidae | 1 | 0.11 | — | — | 4 | 0.16 | — | — | 2 | 0.17 | 7 | 0.08 | + | O | |||||
| 蜚蠊科Blattidae | — | — | 1 | 0.05 | — | — | — | — | — | — | 1 | 0.01 | + | O | |||||
| 鼻螱科Rhinotermitidae | 3 | 0.32 | — | — | — | — | — | — | — | — | 3 | 0.04 | + | D | |||||
| 螱科Termitidae | 2 | 0.21 | — | — | — | — | — | — | — | — | 2 | 0.02 | + | D | |||||
| 半翅目若虫Hemiptera nymphae | — | — | — | — | — | — | 1 | 0.07 | — | — | 1 | 0.01 | + | ||||||
| 上啮科Epilohmanniidae | 3 | 0.32 | 2 | 0.09 | — | — | 1 | 0.07 | — | — | 6 | 0.07 | + | O | |||||
| 鼠啮科Myopsocidae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | O | |||||
| 跳啮科Psyllipsocidae | — | — | — | — | 1 | 0.04 | — | — | — | — | 1 | 0.01 | + | O | |||||
| 沼啮科Elipsocidae | — | — | — | — | 12 | 0.48 | 1 | 0.07 | 1 | 0.09 | 14 | 0.17 | + | O | |||||
| 重啮科Amphientomidae | — | — | — | — | 1 | 0.04 | — | — | — | — | 1 | 0.01 | + | O | |||||
| 管蓟马科Phlaeothripidae | 4 | 0.43 | 27 | 1.24 | 31 | 1.25 | 23 | 1.53 | 13 | 1.11 | 98 | 1.18 | ++ | Ph | |||||
| 蚁甲科Pselaphidae | 1 | 0.11 | — | — | 1 | 0.04 | 1 | 0.07 | 1 | 0.09 | 4 | 0.05 | + | D | |||||
| 鞘翅目幼虫Coleoptera larvae | 1 | 0.11 | 2 | 0.09 | 1 | 0.04 | 1 | 0.07 | 1 | 0.09 | 6 | 0.07 | + | Ph | |||||
| 步甲科幼虫Carabidae larvae | 1 | 0.11 | — | — | — | — | — | — | — | — | 1 | 0.01 | + | Ph | |||||
| 鳞翅目幼虫Lepidoptera larvae | 1 | 0.11 | 4 | 0.18 | 2 | 0.08 | — | — | — | — | 7 | 0.08 | + | Ph | |||||
| 双翅目幼虫Diptera larvae | 35 | 3.72 | 58 | 2.66 | 23 | 0.93 | 16 | 1.06 | 9 | 0.77 | 141 | 1.70 | ++ | S | |||||
| 蚁科Formicidae | 15 | 1.59 | 212 | 9.71 | 77 | 3.11 | 3 | 0.20 | 2 | 0.17 | 309 | 3.74 | ++ | O | |||||
| 总个体数Total individuals | 941 | 2183 | 2476 | 1504 | 1167 | 8271 | |||||||||||||
| 总类群数Total group number | 55 | 51 | 53 | 48 | 34 | 76 | |||||||||||||
| —表示无或未采集到, +++为优势类群, ++为常见类群, +为稀有类群; Ph:植食性Phytophage;F:菌食性Fungivorous forms;Pr:捕食性Predators; S:腐食性Saprozoic;O:杂食性Omnivores;D:枯食性Debris-feeder′ s;Co:粪食性Coprophages | |||||||||||||||||||
5种郁闭度类型下土壤动物群落优势类群存在一定差异。在0.9郁闭度下, 大型土壤动物以蜚蠊科、康叭科、疣科为优势类群, 占59.44%;中小型以等节科、球角科、懒甲螨为优势类群, 占42.08%;在0.8郁闭度下, 大型土壤动物以蜚蠊科为优势类群, 占30.00%, 中小型以等节科为优势类群, 占42.74%;在0.7郁闭度下, 大型土壤动物以步甲科、蜚蠊科、疣科和螱科为优势类群, 占57.94%;中小型以等节科和丽甲螨科为优势类群, 占59.13%;在0.6郁闭度下, 大型土壤动物以蜚蠊科和康叭科为优势类群, 占36.36%, 中小型以等节科为优势类群, 占51.50%;在0.5郁闭度下, 大型土壤动物以蜚蠊科和疣科为优势类群, 占47.17%, 中小型以等节科为优势类群, 占55.18%。
3.2 土壤动物功能类群根据林英华[18]等, 将该研究区土壤动物划分为腐食性、杂食性、植食性、捕食性、菌食性、枯食性和粪食性土壤动物。大型土壤动物以杂食性为主, 占56.03%, 其次是捕食性 (19.19%)>植食性 (10.19%)>腐食性 (8.49%)>枯食性 (5.94%)>粪食性 (0.17%)(图 1A)。其中杂食性和捕食性在0.5—0.8无显著差异, 在0.8—0.9捕食性显著减少, 杂食性显著增加。中小型土壤动物以菌食性 (45.08%) 和腐食性 (40.19%) 为主, 其次为杂食性 (9.90%)>植食性 (4.23%)>捕食性 (0.48%)>枯食性 (0.11%)(图 1B)。其中捕食性在0.5—0.7显著增加, 0.7—0.9无显著变化;随郁闭度增大, 腐食性土壤动物逐渐增加, 菌食性逐渐减少, 且都在0.9郁闭度下形成显著性差异 (P < 0.05;P < 0.01)。

|
| 图 1 不同郁闭度马尾松人工林土壤动物功能类群 Fig. 1 Functional group of soil fauna for different canopy density of a Pinus massoniana Plantation A:大型土壤动物各功能群个体数百分比;B:中小型土壤动物各功能群个体数百分比 |
统计分析结果显示, 不同郁闭度处理对大型土壤动物个体数和类群数均无显著影响 (P > 0.05)。中小型土壤动物个体数随郁闭度在0.5—0.7下显著增加 (P < 0.05), 在0.7—0.9郁闭度下显著降低 (P < 0.05)。中小型土壤动物类群数在0.6—0.9郁闭度无显著变化 (P > 0.05), 在0.5郁闭度则显著降低 (P < 0.05)(图 2)。

|
| 图 2 不同郁闭度马尾松人工林土壤动物类群及数量 (平均值±标准误) Fig. 2 Soil fauna individuals and number of groups for different canopy density of a Pinus massoniana Plantation (mean±SE) A:大型土壤动物类群及数量;B:中小型土壤动物类群及数量; 不同小写字母表示显著差异 (P < 0.05);不同大写字母表示极显著差异 (P < 0.01) |
郁闭度变化对不同土壤层次间大型土壤动物个体数以及枯落物层、0—5 cm层类群数没有显著影响 (P > 0.05), 对5—10 cm层大型土壤动物类群数影响显著 (P < 0.05)。其中, 在0.5—0.8郁闭度下大型土壤动物类群数无显著变化 (P > 0.05), 至0.9郁闭度则显著增多 (P < 0.05)(图 3)。枯落物层中小型土壤动物个体数和5—10 cm土层类群数在0.5—0.7郁闭度显著增高 (P < 0.05), 0.7—0.9郁闭度显著降低 (P < 0.05)。枯落物层类群数在0.5—0.8郁闭度显著升高 (P < 0.05), 且在0.7郁闭度达到极显著水平 (P < 0.01), 0.8郁闭度下枯落物层中小型土壤动物类群数最为丰富, 而0.8—0.9郁闭度则显著降低 (P < 0.05)。0—5 cm土层中小型土壤动物类群数随郁闭度变化不显著 (P > 0.05)(图 3)。

|
| 图 3 不同郁闭度马尾松人工林土壤动物剖面分布 (平均值±标准误) Fig. 3 Profile distribution of soil fauna for different canopy density of a Pinus massoniana Plantation (mean±SE) A:大型土壤动物类群及数量剖面分布;B:中小型土壤动物类群及数量剖面分布; 不同小写字母表示显著差异 (P < 0.05);不同大写字母表示极显著差异 (P < 0.01) |
5种郁闭度处理对大型土壤动物多样性影响不显著 (P > 0.05), 对中小型土壤动物群落多样性产生显著影响 (P < 0.05)。其中, 中小型土壤动物Simpson优势度指数随着郁闭度增大而显著减小 (P < 0.05), Shannon-Wiener多样性指数和Pielou均匀性指数则显著增大 (P < 0.05)。0.9郁闭度下优势度最低, 而多样性指数和均匀性指数则显著高于其他郁闭度 (P < 0.05)。Margalef丰富度指数在0.5—0.6郁闭度显著增高 (P < 0.05), 在0.6—0.9郁闭度则无显著变化 (P > 0.05)(图 4)。

|
| 图 4 不同郁闭度马尾松人工林土壤动物多样性特征 (平均值±标准误) Fig. 4 Diversity features for different canopy density of a Pinus massoniana Plantation (mean±SE) 不同小写字母表示显著差异 (P < 0.05);不同大写字母表示极显著差异 (P < 0.01) |
大型土壤动物多样性指数在枯落物层随郁闭度变化无显著影响 (P > 0.05)。但在0—5 cm层大型土壤动物Simpson优势度和Shanon-Wiener多样性指数差异均显著 (P < 0.05)。5—10 cm土层Shanon-Wiener多样性指数、Pielou均匀性和Margelef丰富性差异显著 (P < 0.05), 其中, 均匀性随郁闭度增大而显著增加 (P < 0.05)(图 5A)。中小型土壤动物多样性特征在0—5 cm均无显著影响 (P > 0.05)。Simpson优势度在枯落物层随郁闭度增大而显著减小 (P < 0.05), Shanon-Wiener多样性和Pielou均匀性显著增大 (P < 0.05), Margelef丰富度在0.5—0.8郁闭度下显著增大 (P < 0.05), 0.8—0.9无显著变化 (P > 0.05)。在5—10 cm土壤层中, 优势度在0.5—0.8无显著变化 (P > 0.05), 0.8—0.9郁闭度时显著增大 (P < 0.05)。Shanon-Wiener多样性在郁闭度0.5—0.7显著增大, 0.7—0.9时显著降低 (P < 0.05), Margelef丰富度在0.5—0.6显著增大, 0.6—0.9显著降低 (P < 0.05)(图 5B)。

|
| 图 5 不同郁闭度马尾松人工林土壤动物土壤不同层次间多样性 (平均值±标准误) Fig. 5 Soil fauna Diversity features for different canopy density of a Pinus massoniana Plantation (mean±SE) A:大型土壤动物多样性指数;B:中小型土壤动物多样性指数; 不同小写字母表示显著差异 (P < 0.05);不同大写字母表示极显著差异 (P < 0.01) |
通过计算, 5个郁闭度梯度下不同体型土壤动物群落Jaccard相似性指数不同, 大型土壤动物相似性指数低于中小型, 在0.3—0.59之间, 属于中等相似或中等不相似, 并以0.7与0.8相似程度最高;中小型土壤动物群落Jaccard相似性指数在0.51—0.69之间, 属中等相似, 并以0.6与0.8相似程度最高 (表 2)。此外, 在枯落物及土壤不同层次之间Jarcard相似性指数也有所差异。其中大型土壤动物除了枯落物层0.8郁闭度与0.7郁闭度、5—10 cm层0.8郁闭度与0.9、0.5郁闭度相似性指数等于0.5(中等相似) 以外, 在其他各层均属中等不相似或极不相似, 最低相似性指数达到0.18(0—5 cm, 郁闭度0.7与0.8;极不相似)。中小型土壤动物0.9郁闭度与0.5、0.6郁闭度Jaccard相似性系数在5—10 cm层极不相似, 在其他各层均属中等不相似或中等相似。
| 郁闭度 Canopy density |
大型Macro | 郁闭度 Canopy density |
中小型Meso or micro | |||||||||
| 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | |||
| 枯落物层Litter layer | 0.5 | 1 | — | — | — | — | 0.5 | 1 | — | — | — | — |
| 0.6 | 0.4 | 1 | — | — | — | 0.6 | 0.63 | 1 | — | — | — | |
| 0.7 | 0.38 | 0.42 | 1 | — | — | 0.7 | 0.59 | 0.64 | 1 | — | — | |
| 0.8 | 0.42 | 0.36 | 0.5 | 1 | — | 0.8 | 0.48 | 0.63 | 0.65 | 1 | — | |
| 0.9 | 0.29 | 0.3 | 0.31 | 0.23 | 1 | 0.9 | 0.5 | 0.56 | 0.59 | 0.67 | 1 | |
| 0—5 cm | 0.5 | 1 | — | — | — | — | 0.5 | 1 | — | — | — | — |
| 0.6 | 0.25 | 1 | — | — | — | 0.6 | 0.45 | 1 | — | — | — | |
| 0.7 | 0.31 | 0.18 | 1 | — | — | 0.7 | 0.41 | 0.42 | 1 | — | — | |
| 0.8 | 0.25 | 0.45 | 0.31 | 1 | — | 0.8 | 0.38 | 0.42 | 0.35 | 1 | — | |
| 0.9 | 0.19 | 0.22 | 0.33 | 0.28 | 1 | 0.9 | 0.39 | 0.44 | 0.32 | 0.45 | 1 | |
| 5—10 cm | 0.5 | 1 | — | — | — | — | 0.5 | 1 | — | — | — | — |
| 0.6 | 0.25 | 1 | — | — | — | 0.6 | 0.39 | 1 | — | — | — | |
| 0.7 | 0.33 | 0.25 | 1 | — | — | 0.7 | 0.3 | 0.37 | 1 | — | — | |
| 0.8 | 0.5 | 0.4 | 0.2 | 1 | — | 0.8 | 0.25 | 0.33 | 0.32 | 1 | — | |
| 0.9 | 0.25 | 0.22 | 0.25 | 0.5 | 1 | 0.9 | 0.14 | 0.23 | 0.33 | 0.35 | 1 | |
| 合计Total | 0.5 | 1 | — | — | — | — | 0.5 | 1 | — | — | — | — |
| 0.6 | 0.47 | 1 | — | — | — | 0.6 | 0.59 | 1 | — | — | — | |
| 0.7 | 0.47 | 0.43 | 1 | — | — | 0.7 | 0.63 | 0.64 | 1 | — | — | |
| 0.8 | 0.48 | 0.53 | 0.59 | 1 | — | 0.8 | 0.51 | 0.69 | 0.65 | 1 | — | |
| 0.9 | 0.35 | 0.34 | 0.39 | 0.3 | 1 | 0.9 | 0.53 | 0.57 | 0.61 | 0.63 | 1 | |
对不同郁闭度下土壤动物主要类群与土壤环境因子进行CCA排序 (置换系数499), 结果经Monte Carfo Test检验, 第一排序轴和其余所有排序轴均显著 (F=1.876, P=0.036;F=1.372, P=0.022)。其中第一排序轴与郁闭度呈极显著正相关, 与含水量和全P呈显著负相关, 第二排序轴与N含量呈显著正相关 (表 3)。前两个排序轴分别解释了36.5%和53.8%的土壤动物类群与几个土壤环境因子的关系, 在一定程度上反映了不同土壤动物类群和环境间的关系。由排序图 (图 6) 可知, 不同郁闭度土壤动物对环境因子的响应存在差异。其中, 寄螨科、双翅目幼虫、大蚓类、美绥螨科受郁闭度和含水量影响较大;园蛛科、鞘翅目幼虫、矮蒲螨科、跳虫科、管蓟马科受pH、全P影响明显, 有机碳主要影响康叭科、线虫、蚁甲科、疣科和大翼甲螨科。第二排序轴主要反映全N对土壤动物主要类群的影响, 其中蜚蠊科、跳蛛科和蚁科受N含量影响较大。球角科、奇马陆科、副铗叭科和若甲螨科受土壤环境因子的影响最小。吸螨科、隐翅甲螨科、懒甲螨科、等节科、丽甲螨科、步甲螨科受各个土壤环境因子的影响均较大。
| 环境变量 Envionment variables |
排序轴Ordination axes | 环境变量 Envionment variables |
排序轴Ordination axes | ||
| Axis1 | Axis2 | Axis1 | Axis2 | ||
| 郁闭度Canopy density | 0.7773** | -0.4304 | 有机碳Soil organic carbon | 0.2706 | 0.1642 |
| 土壤pH | -0.3409 | -0.1274 | 全N Total nitrogen | -0.0266 | 0.5459* |
| 含水量Moisture content | -0.4768* | 0.2464 | 全P Total phosphorus | -0.5032* | -0.1048 |
| 容重Bulk density | -0.4341 | -0.0564 | |||

|
| 图 6 不同郁闭度马尾松人工林土壤动物与土壤环境因子CCA分析 Fig. 6 CCA analysis between soil fauna groups and environmental variables for different canopy density of a Pinus massoniana Plantation CD:郁闭度canopy density; pH:土壤pH; MC:含水量moisture content; SOC:有机碳soil organic carbon; TN:全氮total nitrogen; TP:全磷total phosphorus; Pygm:矮蒲螨; Oryi:奥地蜈蚣; Oppi:奥甲螨; Cabi:步甲; Cabo:步甲螨; Galu:大翼甲螨; Isot:等节; Blat:蜚蠊; Para:副铗叭; Phla:管蓟马; Parasi:寄螨; Campo:康叭; Noth:懒甲螨; Liac:丽甲螨; Amer:美绥螨; Asci:囊螨科; Paradox:奇马陆; Cole:鞘翅目幼虫; Lumb:大蚓; Hypogas:球角; Orib:若甲螨; Dipt:双翅目幼虫; Podu:跳虫; Salt:跳蛛; Term:螱科; Bdel:吸螨; Nema:线虫; Thom:蟹蛛; Psel:蚁甲; Form:蚁科; Stap:隐翅甲; Nean:疣; Aran:圆蛛 |
在森林生态系统中, 由于郁闭度不同, 改变了系统的群体结构, 相应地引起林内微生境的变化, 包括光强、空气温度、空气相对湿度、土壤含水率、地温的日变化特和空间分布规律, 以及降水的再分配等[19]。研究表明, 林下微生态环境因子与林分郁闭度的相关性达到了显著水平, 且高郁闭度林下极值变化范围更小[20-21]。郁闭度的减小对应着林隙的增大和增多, 改变了光照在林分中的分布, 进而改变了林木各部位对光资源的利用和再分配。同时, 林内光照增强使土壤温度升高, 加快了土壤中有机质的分解, 使土壤速效养分增加, 从而提高了林地土壤肥力[22]。土壤环境因子的变化引起土壤动物类群和个体数分布差异, 决定了郁闭度变化也会影响到土壤生态系统中土壤动物的群落结构组成[23-24]。具体来讲, 土壤动物个体密度与土壤有机质含量、TN、TP呈极显著正相关, 与土壤含水量呈显著正相关, 与土壤容重呈显著负相关[25]。本研究表明, 在不同的郁闭度马尾松人工林中, 土壤动物主要类群与林下土壤P含量和含水量显著相关, 同时也受有机质、pH、容重和TN的共同影响。可见, 郁闭度对林下微环境的改变是土壤动物群落结构发生变化的主要原因。
在0.5郁闭度下, 虽然土壤含水量、有机质及N、P含量均较高, 灌木和草本恢复较好, 但该郁闭度下土壤动物个体数、类群数均较少。这是因为林冠开阔度相对较大, 温湿度等极值变化大, 一些对环境敏感的稀有种如表刻螨科、虫穴螨科、寄螨科、卷叶蛛、漏斗蛛科等减少或消失。光强直接使土壤动物向更深的土壤层次或周围环境迁移;穿透雨对地面的冲刷给土壤动物造成干扰, 且光照相对充足, 凋落物分解速度快, 林下植被恢复消耗了部分养分, 使腐食性土壤动物减少。等节科依靠体表呼吸, 受含水量和光照的影响较大[26-27], 占了该郁闭度下46.37%的个体, 因而优势度明显, 而多样性和均匀性则降低。一般认为, 生活在土壤水膜中的线虫在含水量高的土壤中数量偏高, 当超过15%会抑制线虫、双翅目的个体数量[28]。本实验样地含水量均在19.64%以上, 随着郁闭度增大, 土壤含水量显著降低 (表 1), 而线虫数量增加。这是因为含水量超过峰值后的抑制作用, 还是受到其他土壤肥力如有机碳和全P含量的影响, 还需要做进一步研究。随着郁闭度增加, 中小型土壤动物总类群数在郁闭度大于0.6时显著增加, 总个体数在0.7郁闭度之后显著减小。枯落物层中小型土壤动物个体数以及5—10 cm土层类群数在0.5—0.7显著升高, 而0.7—0.9下降 (P < 0.05)。在0.7郁闭度时土壤动物个体数最大, 这是因为该郁闭度不论是光照, 还是土壤湿度、营养条件等方面都比较适中, 更有利于土壤动物生存, 受郁闭度影响较大的懒甲螨科、奥甲螨科、丽甲螨科、鞘翅目幼虫、等节科、隐翅甲科等数量增多。而至0.9郁闭度, 此时林内光照不足, 林冠截留使其含水量偏低 (表 1), 凋落物长期堆积, 分解缓慢;林下植被缺失, 使食物源单一, 也不利于土壤动物的生存;且该郁闭度下土壤pH最低, 限制了园蛛科、鞘翅目幼虫、跳虫、管蓟马科等土壤动物的分布。因此该郁闭度下中小型土壤动物个体数和5—10 cm层多样性和丰富度均有所下降。说明0.7郁闭度更有利于保持马尾松人工林下土壤动物数量及种类丰度。
本研究中郁闭度显著改变了中小型土壤动物群落结构, 引起类群数、个体数及多样性指数发生变化, 但对大型土壤动物个体数、类群数均无显著性影响, 只有5—10 cm土层类群数在0.9郁闭度表现出显著增多的趋势。这说明中小型土壤动物能够更敏感地反映林分郁闭度变化, 而大型土壤动物移动力较强, 营养结构更为复杂, 在生境中具有相对稳定性[29], 受郁闭度影响较弱。
郁闭度处理使林分内生物种间关系发生了变化。杂食性土壤动物生态位广泛, 生存适应能力强, 在本研究中所占比例较高。捕食性动物是土壤生物群落中的消费者, 以植食性动物和腐食性动物为食, 营养级别高。本研究所采集到的捕食性土壤动物个体所占的比例高于植食性, 说明群落内部竞争和捕食作用使群落内各物种达到动态平衡, 减少部分植食性害虫的生存和繁衍[30]。而韩宝瑜[31]等的研究表明马尾松纯林 (林下植被盖度0.9) 中植食性昆虫种数最多、个体数最大, 天敌的种数和个体数较少。这主要是受到林下植被丰富程度的影响。随着郁闭度增大, 中小型腐食性土壤动物个体数百分比显著增加, 而菌食性显著减少。这是因为郁闭度越大, 土壤温度降低, 土壤微生物数量和活性均下降[32], 导致菌食性土壤动物可食性食物源减少, 故而随之减少。腐食性随郁闭度增大其比例逐渐增多, 是因为高郁闭度下枯落物大量堆积[33]。而郁闭度越小, 林下温度升高使枯落物很快分解, 加之林下植被恢复消耗了土壤中的养分, 故囊螨、维螨及其他受有机质影响的腐食性土壤动物减少。
土壤动物种类的多样性反映了其生活环境的稳定性和群落营养通道的复杂程度, 是衡量群落结构水平和生态学特征的重要量度指标[34]。多样性越高, 代表采样区生态结构越稳定, 则有利于土壤动物的生存与繁衍[35]。在本次实验中, 郁闭度变化对大型土壤动物在各土壤层次中的多样性影响显著, 且均匀性逐渐增强。随着郁闭度增大, 中小型土壤动物群落的Simpson优势度指数逐渐减小, Shannon-Wiener多样性指数和Pielou均匀性指数则显著增大。可能是因为林冠荫蔽作用使林下环境条件得到改善, 能够适宜更多类群的土壤动物生存, 因此多样性指数也逐渐增大, 均匀性增强。Margalef指数也表现在高郁闭度下更加丰富。然而, 郁闭度过大会限制林下植被恢复, 植食性土壤动物所占比例太小, 使整个林分的营养结构单一, 不利于生态系统的长期稳定。
5 结论郁闭度变化引起林下环境条件相应改变, 对大型土壤动物群落结构影响较小, 主要对中小型土壤动物类群及数量、多样性、功能类群等在各郁闭度下及不同土壤层次之间影响显著, 郁闭度过高或过低都不利于保持土壤动物群落多样性和稳定性。0.7郁闭度马尾松人工林下土壤动物优势度、丰富度都保持在适中水平, 虽然均匀性和多样性还不太理想, 但随着林下植被恢复的时间尺度增加, 该郁闭度马尾松人工林可能更有利于土壤动物结构和功能的发挥。但是, 本研究仅是郁闭度控制后马尾松人工林土壤动物群落的初期调查, 研究结果反应的仅是一种趋势, 这种趋势是否会随着郁闭度控制时间的推移而发生改变还有待进一步研究。
| [1] | Brussaard L, Pulleman M M, Ouédraogo É, Mando A, Six J. Soil Fauna and Soil Function in the Fabric of the Food Web. Pedobiologia, 2007, 50(6): 447–562. DOI:10.1016/j.pedobi.2006.10.007 |
| [2] | 武海涛, 吕宪国, 杨青, 姜明. 土壤动物主要生态特征与生态功能研究进展. 土壤学报, 2006, 43(2): 314–323. |
| [3] | Peng Y, Yang W Q, Li J, Wang B, Zhang C, Yue K, Wu F Z. Contribution of Soil Fauna to Foliar Litter-Mass Loss in Winter in an Ecotone between Dry Valley and Montane Forest in the Upper Reaches of the Minjiang River. Plos one, 2015, 10(4): e0124605. DOI:10.1371/journal.pone.0124605 |
| [4] | Huang W, Siemann E, Xiao L, Yang X F, Ding J Q. Species-Specific Defence Responses Facilitate Conspecifics and Inhibit Heterospecifics in Above-Belowground Herbivore Interactions. Nature Communications, 2014, 5: 4851. DOI:10.1038/ncomms5851 |
| [5] | Zaitsev A S, Chauvat M, Wolters V. Spruce Forest Conversion to a Mixed Beech-Coniferous Stand Modifies Oribatid Community Structure. Applied Soil Ecology, 2014, 76(2): 60–67. |
| [6] | 张良辉. 川西亚高山云杉人工林分结构调控下植物与土壤动物多样性研究[D]. 雅安: 四川农业大学, 2010. http://cdmd.cnki.com.cn/Article/CDMD-10626-2010244424.htm |
| [7] | 刘茂秀, 史军辉, 王新英, 彭秋梅. 林分郁闭度和龄级对防护林碳汇功能及固碳价值的影响. 东北林业大学学报, 2011, 39(6): 30–32. |
| [8] | 国家林业局. 中国森林资源简况——第八次全国森林资源清查, 2013[2015]. http://www.forestry.gov.cn/gjslzyqc.html. |
| [9] | 洪伟, 吴承祯. 马尾松人工林经营模式及其应用. 北京: 中国林业出版社, 1999. |
| [10] | 杨平平, 徐仁扣, 黎星辉. 淋溶条件下马尾松针对土壤的酸化作用. 生态环境学报, 2012, 21(11): 1817–1821. |
| [11] | 杨承栋, 孙启武, 焦如珍, 卢立华. 大青山一二代马尾松土壤性质变化与地力衰退关系的研究. 土壤学报, 2003, 40(2): 267–273. |
| [12] | 李永宁, 张宾兰, 秦淑英, 李帅英, 黄选瑞. 郁闭度及其测定方法研究与应用. 世界林业研究, 2008, 21(1): 40–56. |
| [13] | 周彬, 韩海荣, 康峰峰, 程小琴, 宋娅丽, 刘可, 李勇. 太岳山不同郁闭度油松人工林降水分配特征. 生态学报, 2013, 33(5): 1645–1653. |
| [14] | 王邵军, 阮宏华, 汪家社, 徐自坤, 吴焰玉. 武夷山典型植被类型土壤动物群落的结构特征. 生态学报, 2010, 30(19): 5174–5184. |
| [15] | 尹文英. 中国亚热带土壤动物. 北京: 科学出版社, 1992. |
| [16] | Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Berkeley and Los Angeles: University of California Press, 1979. |
| [17] | 尹文英. 中国土壤动物. 北京: 科学出版社, 2000. |
| [18] | 林英华, 贾旭东, 徐演鹏, 李慧仁, 刘学爽, 徐永波, 韦昌雷, 刘三章, 王立中. 大兴安岭典型森林沼泽类型地表土壤动物群落与生态位分析. 林业科学, 2015, 51(12): 53–62. |
| [19] | 宋小帅, 康峰峰, 韩海荣, 于晓文, 周彬, 程小琴. 太岳山不同郁闭度油松人工林枯落物及土壤水文效应. 水土保持通报, 2014, 34(3): 102–108. |
| [20] | 康文星, 邓湘雯, 赵仲辉. 林冠截留对杉木人工林生态系统物质循环的影响. 林业科学, 2006, 42(12): 1–5. |
| [21] | 成向荣, 冯利, 虞木奎, 吴统贵, 孙海菁, 王宗星, 张翠. 间伐对生态公益林冠层结构及土壤养分的影响. 生态环境学报, 2010, 19(2): 355–359. |
| [22] | 李岩. 阔叶红松林林隙小气候及土壤物理性质特征比较研究[D]. 哈尔滨: 东北林业大学, 2007. |
| [23] | Taki H, Inoue T, Tanaka H, Makihara H, Sueyoshi M, Isono M, Okabe K. Responses of Community Structure, Diversity, and Abundance of Understory Plants and Insect Assemblages to Thinning in Plantations. Forest Ecology and Management, 2010, 259(3): 607-613. |
| [24] | 刘任涛, 赵哈林, 赵学勇. 科尔沁沙地不同造林类型对土壤动物多样性的影响. 应用生态学报, 2012, 23(4): 1104–1110. |
| [25] | 武海涛, 吕宪国, 杨青, 姜明, 佟守正. 三江平原湿地岛状林土壤动物群落结构特征及影响因素. 北京林业大学学报, 2008, 30(2): 50–58. |
| [26] | Ponge J F. Biocenoses of Collembola in Atlantic Temperate Grass-woodland Ecosystems. Pedobiologia, 1993, 37(4): 223–244. |
| [27] | Nickerl J, Helbig R, Schulz H J, Werner C, Neinhuis C. Diversity and Potential Correlations to the Function of Collembola Cuticle Structures. Zoomorphology, 2013, 132(2): 183–195. DOI:10.1007/s00435-012-0181-0 |
| [28] | 张瑞益. 北方主要草地类型土壤线虫群落结构及多样性的比较研究[D]. 长春: 东北师范大学, 2015. |
| [29] | 黄丽荣, 张雪萍. 大兴安岭寒温带地区中小型土壤动物群落特征. 应用与环境生物学报, 2008, 14(3): 388-393. http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_yyyhjswxb200803020 |
| [30] | 朱新玉, 高宝嘉, 胡云川. 森林草原交错带土壤节肢动物同功能种团组成及分布. 江苏农业科学, 2011, 39(3): 478–581. |
| [31] | 韩宝瑜. 马尾松林节肢动物群落的组成及多样性. 生物多样性, 2001, 9(1): 62–67. |
| [32] | 张鼎华, 叶章发, 范必有, 危廷林. 抚育间伐对人工林土壤肥力的影响. 应用生态学报, 2001, 12(5): 672–676. |
| [33] | 张雪萍, 侯威岭, 陈鹏. 东北森林土壤动物同功能种团及其生态分布. 应用与环境生物学报, 2001, 7(4): 370–374. |
| [34] | 谭波, 吴福忠, 杨万勤, 张健, 徐振锋, 刘洋, 苟小林. 不同林龄马尾松人工林土壤节肢动物群落结构. 应用生态学报, 2013, 24(4): 1118–1124. |
| [35] | 王振中, 张友梅, 李忠武. 黄山森林生态系统土壤动物群落结构特征及其多样性. 林业科学, 2009, 45(10): 168–173. |