 2017, Vol. 37
2017, Vol. 37文章信息
- 张旭龙, 马淼, 吴振振, 张志政, 高睿, 石灵玉
- ZHANG Xulong, MA Miao, WU Zhenzhen, ZHANG Zhizheng, GAO Rui, SHI Lingyu.
- 不同油葵品种对盐碱地根际土壤酶活性及微生物群落功能多样性的影响
- Effects of Helianthus annuus varieties on rhizosphere soil enzyme activities and microbial community functional diversity of saline-alkali land in Xinjiang
- 生态学报. 2017, 37(5): 1659-1666
- Acta Ecologica Sinica. 2017, 37(5): 1659-1666
- http://dx.doi.org/10.5846/stxb201510172095
-
文章历史
- 收稿日期: 2015-10-17
- 网络出版日期: 2016-07-13

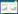




盐碱土会导致土壤肥力下降, 毒害作物根系, 影响作物生长, 是限制农业发展的瓶颈之一。据统计, 我国盐碱土地面积约为0.36亿hm2, 占全国可利用土地总面积的4.88%, 并以每年1.00%的速度增长[1]。研究报道称新疆有80.0%的耕地面积属于次生盐碱化土地[2], 严重制约了当地的经济发展。因此, 对盐碱土的改良已刻不容缓。目前, 普遍认为最有效的开发利用盐碱地的方法是在盐碱地上种植耐盐碱的作物[3-5], 其不仅能提高土地利用率和生产力水平, 而且能促进盐碱土环境中农业生态的良性循环和环境改善[6-8]。
油葵 (Helianthus annuusL.) 作为全球重要的油料植物, 在食品、能源、轻工业等领域备受国内外学者的青睐和关注。油葵具有耐盐碱性强等特点[9-12], 是用于开发盐碱地的少数经济作物之一。目前国内外关于油葵的研究报道较多, 但大都集中在油葵种子和幼苗的耐盐性研究[13-17]、不同程度盐碱化土壤对油葵生长发育和产量的影响[18-20]以及化感作用等方面的研究[18-20]。鲜有从种植油葵对盐碱地土壤质量改善效果的研究报道。本文以在新疆广为栽种的新葵4号、新葵6号、新葵10号和美国矮大头 (567DW) 等4个油葵品种为材料, 通过盆栽试验, 从土壤理化性质、相关酶活性和土壤微生物群落功能多样性等方面来评价种植不同油葵品种对盐碱土的改良效果, 筛选出较适宜在盐碱地种植的油葵品种, 以期为盐碱地的改良、生态系统的恢复与重建以及区域经济的发展提供科学依据。
1 材料与方法 1.1 供试土壤与材料该试验所需的土壤采自石河子市周边盐碱土, 土壤类型为轻度盐碱沙壤土。供试油葵品种 (新葵4号、新葵6号、新葵10号和美国矮大头) 由新疆农垦科学院作物所提供。盆栽试验在高30 cm, 直径20 cm的花盆中进行, 土壤厚度约为27 cm, 充分浇水, 隔夜备用。种子用温水浸泡过夜, 每盆点播2穴, 每穴3颗, 穴距10 cm, 设3次重复。然后, 将其置于温度为25 ℃、光强为450 μmol m-2 s-1、光照时间为12 h/d的光照培养箱 (GXZ-430D) 中培养。出苗后定苗, 每穴留1颗苗, 每3d等量浇水1次, 培养80多天至油葵开花时取样。
1.2 土壤理化性质和酶活性分析取油葵根际土, 去除残余根系和杂质后将相同处理不同重复的土壤样品混和均匀并过1 mm筛, 置于4℃冰箱中保存待用, 以未种植油葵的土壤为空白对照 (CK)。用梅特勒pH计 (FE20K) 测定根际土壤pH值 (水:土=2.5: 1), 全氮用高氯酸-硫酸消化法, 速效氮用碱解蒸馏法, 速效磷用0.5 mol/L NaHCO3浸提-钼锑抗比色法, 速效钾用NH4OAc浸提-火焰光度法;蔗糖酶、脲酶和磷酸酶的测定方法参照关松荫[21]的《土壤酶及其研究法》。
1.3 土壤微生物功能多样性分析微生物群落功能多样性采用Biolog (Biolog-ECO PlateTM) 方法进行分析。称取相当于10 g烘干土的鲜土加入到已装有100 mL灭菌的0.05 mol/L的磷酸盐缓冲液 (PBS) 的三角瓶中, 室温震荡60 min, 然后用PBS缓冲液稀释至1000倍液, 再震荡30 min, 吸取稀释液接种到生态板 (ECO板) 中, 每孔加150 μL。将接种好的微平板置于25℃的恒温培养箱中避光培养, 每隔12 h在Biolog EmaxTM读数器上读取波长590 nm下的光密度值, 持续144 h。用31个孔的平均光密度值表示微生物代谢的整体活性 (Average well color development, AWCD), 并计算Shannon多样性指数 (H)、Shannon均匀度指数 (E)、Shannon优势度指数 (D) 和碳源利用丰富度指数 (S), 具体计算公式如下[22-24]:
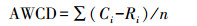
|
式中, Ci为每个孔的光密度值, Ri为对照孔的光密度值, n=31, 为ECO板的孔数。
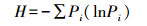
|
式中, Pi=(Ci-Ri)/∑(Ci-Ri)。
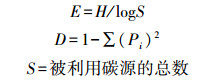
|
采用SPSS 16.0软件对实验数据进行单因素方差分析、主成分分析 (PCA) 和聚类分析。相关数据图制作在Excel中完成。
2 结果与分析 2.1 不同油葵品种对土壤化学性质的影响不同油葵品种对土壤理化性质的影响有显著的根际效应 (表 1)。种植4个油葵品种均能降低根际土壤pH值, 且新葵6号的降低效果最为显著。与其余3个油葵品种相比, 种植新葵4号后能显著提高根际土壤碱解氮、速效磷和速效钾的含量, 且以新葵4号的效果最佳。种植新葵10号后的根际土壤碱解氮含量最小且显著低于CK。种植新葵6号后, 其根际土壤全氮含量最高, 并显著高于CK。
| 油葵品种 Varieties | pH | 全氮 Total N/ (g/kg) | 碱解氮 Alkaline N/ (mg/kg) | 速效磷 Available P/ (mg/kg) | 速效钾 Available K/ (mg/kg) |
| CK | 7.88±0.11 a | 0.73±0.02 c | 51.45±1.39 c | 23.58±0.92 c | 213.26±7.63 d |
| 新葵4Xinkui 4 | 7.66±0.09 c | 0.99±0.06 a | 56.24±0.97 a | 25.39±1.25 a | 228.25±8.21 a |
| 新葵6号Xinkui 6 | 7.64±0.12 c | 1.02±0.08 a | 52.36±0.91 c | 21.33±1.04 d | 196.57±7.12 e |
| 新葵10号Xinkui 10 | 7.78±0.14 b | 0.72±0.03 c | 50.02±1.02 d | 24.73±1.01 b | 219.77±9.03 c |
| 美国矮大头Aidatou | 7.85±0.08 a | 0.79±0.03 b | 53.77±0.98 b | 23.89±1.32 c | 224.97±8.87 b |
| 同列不同字母表示相同处理间差异显著 (P < 0.05) | |||||
从表 2可以看出, 新葵6号和新葵10号的根际土壤蔗糖酶活性显著高于新葵4号、美国矮大头和CK, 且以新葵6号的根际土壤蔗糖酶活性最高, 种植新葵6号和新葵10号能显著提高根际土壤蔗糖酶的活性。种植新葵4号对根际土壤脲酶活性影响最大, 且与CK有显著性差异。新葵4号和美国矮大头的根际土壤磷酸酶活性高于CK, 且新葵4号的根际土壤磷酸酶活性最高, 并与CK有显著性差异, 种植新葵4号能显著提高其根际土壤磷酸酶活性。
| 油葵品种 Varieties | 蔗糖酶Invertase/ (mg g-1 d-1) | 脲酶Urease/ (mg g-1 d-1) | 磷酸酶Peroxidase/ (mg g-1 d-1) |
| CK | 9.43±0.24 b | 1.57±0.03 b | 10.42±0.44c |
| 新葵4号Xinkui 4 | 9.03±0.07 c | 1.63±0.02 a | 10.62±0.31 b |
| 新葵6号Xinkui 6 | 11.61±0.48 a | 1.56±0.04 b | 8.31±1.22 a |
| 新葵10号Xinkui 10 | 8.87±0.28 c | 1.58±0.03 ab | 10.01±0.31 d |
| 美国矮大头Aidatou | 9.21±0.31 bc | 1.60±0.03 ab | 10.45±0.36 bc |
AWCD是表征土壤微生物对总体碳源的利用能力, 反映其生物活性的一个重要指标[25]。31种碳源的AWCD变化如图 1所示, 随着温育时间的延长, AWCD逐渐升高, 在0—12 h时, 不同处理间的AWCD值均为零, 在12 h之后, 种植4个油葵品种后的根际微生物均开始利用单一碳源进行代谢, 在24 h之后CK的微生物也开始进行代谢, 种植新葵4号后的根际微生物在24 h之后基本进入对数增长期。新葵6号、新葵10号、美国矮大头和CK的微生物在36 h之后基本进入对数增长期, 并在144 h时都趋于稳定。并且CK的AWCD在整个温育时期内都为最小, 种植不同油葵品种后均显著提高了第72 h的根际微生物代谢活性 (P<0.05), 且以种植新葵4号的最高, 较之CK增加了113.83%。

|
| 图 1 根际微生物AWCD随温育时间的变化 Fig. 1 AWCD changes with incubation time of different treatments |
Shannon多样性指数可以从不同侧面反映土壤微生物群落功能多样性, 是目前应用最广泛的群落多样性指数之一[26]。通常把颜色变化孔数作为根际微生物群落功能多样性的丰富度[27]。从图 2可以看出, 土壤微生物对碳源利用的H、D、S变化表现出类似的规律:新葵4号的最高而CK最小且有显著性差异。种植4个油葵品种后的H、D均高于CK且有显著性差异;种植4个油葵品种后的S均高于CK。Shannon均匀度指数 (E) 以CK最大, 新葵4号的最小。

|
| 图 2 不同油葵品种土壤微生物群落多样性指数 Fig. 2 Diversity indices for soil microbial communities in different varietiesHelianthus annuus |
对培养72 h的31种碳源利用情况进行主成分分析, 共提取3个主成分, 第一主成分 (PC1)、第二主成分 (PC2) 和第三主成分 (PC3) 分别可以解释所有变量的52.8%、24.2%和13.9%, 3个主成分累积方差贡献率达到90.9%, 可以较全面的概括31个变量的特征。新葵4号在PC1得分最高, 新葵6号在PC2上得分最高, 新葵10号在PC3上得分最高。
因子载荷反映了主成分与碳源利用的相关程度, 因子载荷绝对值越接近于1, 表示该碳源对主成分的影响越大。表 3所示, 与第一主成分相关性较高的碳源有17个, 主要包括酚酸类化合物 (1个)、聚合物 (3个)、羧酸类化合物 (4个)、碳水化合物 (9个);与第二主成分相关性较高的碳源有10个, 主要包括酚酸类化合物 (2个)、胺类化合物 (1个)、氨基酸 (5个)、羧酸类化合物 (1个)、碳水化合物 (1个);与第三主成分相关性较高的碳源有5个, 主要包括胺类化合物 (1个)、氨基酸 (1个)、羧酸类化合物 (2个)、碳水化合物 (1个)。其中, 聚合物、羧酸类化合物、氨基酸和碳水化合物是微生物利用的主要碳源。
| 碳源类别 Carbon source guild | 底物 Substrates | 主成分1 PC1 | 主成分2 PC2 | 主成分3 PC3 |
| 酚酸类化合物Phenolic acids | 2-羟基苯甲酸 | -0.852 | -0.509 | -0.080 |
| 4-羟基苯甲酸 | -0.433 | 0.874 | 0.113 | |
| 胺类化合物Amines/amids | 苯乙基胺 | -0.151 | 0.840 | 0.490 |
| 腐胺 | 0.475 | 0.308 | 0.683 | |
| 聚合物Polymers | 吐温40 | -0.948 | 0.303 | -0.001 |
| 吐温80 | 0.376 | 0.490 | -0.757 | |
| α-环式糊精 | 0.975 | -0.031 | 0.168 | |
| 肝糖 | 0.998 | -0.029 | 0.030 | |
| 氨基酸Amino acids | L-精氨酸 | -0.258 | -0.696 | 0.211 |
| L-天冬酰胺酸 | 0.444 | 0.729 | 0.266 | |
| L-苯基丙氨酸 | 0.450 | -0.853 | -0.256 | |
| L-丝氨酸 | 0.123 | 0.789 | 0.058 | |
| L-苏氨酸 | 0.260 | 0.831 | -0.400 | |
| 甘氨酰-L-谷氨酸 | 0.252 | -0.293 | 0.823 | |
| 羧酸类化合物Carboxylic acids | 丙酮酸甲脂 | 0.668 | -0.207 | 0.458 |
| D-半乳糖醛酸 | -0.965 | -0.104 | 0.233 | |
| γ-羟基丁酸 | 0.834 | 0.075 | -0.456 | |
| D-葡萄胺酸 | -0.419 | 0.496 | 0.739 | |
| 衣康酸 | 0.200 | 0.460 | -0.611 | |
| α-丁酮酸 | -0.927 | 0.184 | 0.241 | |
| D-苹果酸 | 0.469 | 0.802 | -0.282 | |
| 碳水化合物Carbohydrates | β-甲基D-葡萄糖苷 | 0.991 | -0.046 | -0.129 |
| D-半乳糖内酯 | -0.808 | 0.259 | -0.519 | |
| D-木糖 | 0.988 | -0.027 | 0.114 | |
| I-赤藻糖醇 | 0.905 | -0.072 | 0.104 | |
| D-甘露醇 | 0.078 | 0.938 | 0.290 | |
| N-乙酰基-D-葡萄胺 | 0.976 | -0.138 | 0.143 | |
| D-纤维二糖 | 0.951 | 0.119 | 0.097 | |
| 葡萄糖-1-磷酸盐 | 0.993 | -0.116 | 0.018 | |
| α-D-乳糖 | 0.988 | 0.049 | 0.100 | |
| D, L-α-甘油 | 0.999 | -0.025 | -0.022 |
对培养72 h的31种碳源的利用情况进行聚类分析, 共分成2类:新葵6号、新葵10号、美国矮大头和CK聚为一类, 新葵4号单独成类 (图 3)。

|
| 图 3 土壤微生物碳源利用聚类分析 Fig. 3 Cluster analysis of carbon utilization by soil microbial communities |
种植耐盐植物可较好的改良盐碱地, 提高盐碱地土壤肥力[28]。本研究发现, 在盐碱地中种植这4个油葵品种对根际土壤理化性质均有不同程度的改良效果, 种植新葵6号对根际土壤pH值的降低和全氮含量的提高效果最为显著, 种植新葵4号能显著提高根际土壤碱解氮、速效磷和速效钾的含量且效果最佳, 种植新葵6号显著降低了根际土壤速效磷和速效钾的含量。
土壤酶来自动、植物和微生物的活体或残体的分泌和释放, 对土壤中养分的循环至关重要, 已被用作微生物的活性指标[29]和土壤肥力指标[30-32]。土壤蔗糖酶、脲酶和磷酸酶广泛存在于土壤中且是土壤中碳、氮、磷和钾素循环的关键酶[21, 33], 其酶活性的大小与土壤营养呈正相关关系[34-35]。种植新葵6号后的根际土壤蔗糖酶活性最高, 种植新葵4号后的根际土壤脲酶活性和磷酸酶活性最高。较高的土壤酶活性可在一定程度上保证油葵生长所需的营养。
根系分泌物是构成植物不同根际微生态特征的关键因素[36], 不同植物的根际分泌物的种类和数量有差异, 对根际土壤微环境中的酶活性、养分循环, 甚至是根际微生物的生长代谢和多样性都有较大影响[37], 在4个油葵品种中, 种植新葵4号对根际土壤理化性质和酶活性的影响较大, 其余3个油葵品种的影响较小, 其可能的原因是, 4个不同的油葵品种在根系分泌物的种类和数量上有差异, 而这些差异使新葵4号对根际土壤微环境的影响最为显著。
3.2 不同油葵品种对AWCD和多样性指数的影响光密度平均颜色变化率AWCD反应土壤微生物利用碳源的能力和代谢活性的大小, AWCD越大, 其代谢活性越强[38]。本研究表明, 种植4个油葵品种后能缩短微生物调整期的时长, 与其余4个处理相比, 种植新葵4号使其根际微生物提前12 h进入对数期增长。在144 h的温育期内, 各处理间的AWCD表现出如下规律:新葵4号>新葵6号>美国矮大头>新葵10号>CK, 说明种植油葵可以提高盐碱地土壤微生物的代谢活性, 改善微生物利用碳源的能力, 其中以种植新葵4号的效果最优。
碳源代谢的多样性指数分析可以准确地反映微生物群落功能多样性的变化, Shannon指数能客观地反映物种的丰富度和均匀度。H、D、E和S是表示群落多样性的最常用指标。种植4个油葵品种后均能不同程度地提高H、D、S, 并表现出相似的规律:4个油葵品种的H、D、S均高于CK, 且以种植新葵4号的指数最高, 说明在盐碱地种植油葵能显著提高土壤微生物群落功能多样性, 且以种植新葵4号后的提高效果最为明显。种植新葵4号后的E最小, 而CK的最大, 与其余3种指数变化趋势不一致, 可能的原因是在肥力贫瘠的盐碱土中, 由于新葵4号的根系分泌物的种类和数量间的差异而引起的, 这些差异打破了原有微环境中的生态平衡, 使新葵4号的根际微生物发生较大变化。
3.3 微生物碳源利用的主成分分析和聚类分析主成分分析采用降维的思想, 分析种植不同油葵品种之后, 对其根际土壤微生物碳源利用的影响是否存在差异[39-40]。不同的作物对根际土壤微生物群落的影响不同[41]。主成分分析表明种植不同油葵品种对盐碱地土壤微生物的碳源利用能力影响显著, 改变了盐碱地土壤微生物群落的代谢特征。样本在主成分上的得分和微生物对碳源底物的利用能力相关[42]。在第一主成分上, 解释了大部分的变异, 新葵4号分布PC1正方向, 得分最高, 分别是新葵6号、新葵10号、美国矮大头和CK的4.4、2.85、5.08和6.38倍。种植新葵4号对盐碱地土壤微生物群落功能影响最大。种植不同油葵品种通过影响盐碱地根际土壤微环境, 改变其微生物群落结构, 最终使盐碱地土壤微生物碳源代谢发生改变, 这与Meng Q J[43]等人研究不同植被对土壤微生物群落功能多样性所得结果相似。
聚类分析是一种多元分类的统计方法, 根据样本的数据特征按照性质上的亲疏程度进行自动分类, 类内部的样本在特征上具有相似性, 不同类间样本特征差异性较大[44]。新葵6号、新葵10号、美国矮大头和CK聚为一类, 说明种植这3个油葵品种对盐碱地根际土壤微生物群落功能多样性的影响程度与CK相似。新葵4号单独成类, 且欧氏距离大, 说明新葵4号与新葵6号、新葵10号、美国矮大头和CK差异性较大, 表明种植新葵4号对盐碱地土壤微生物群落功能多样性有较大影响。这与根际土壤理化性质、酶活性分析以及主成分分析所得结果相同。
4 结论土壤微生物是土壤中最活跃的组分之一, 通过参与土壤形成、有机质分解、腐殖质形成、土壤养分转化和循环等影响土壤环境[45-47], 也是土壤酶的来源之一[48], 土壤酶活性和土壤微生物群落功能多样性对土壤微生态环境的改善和土壤的可持续利用至关重要。
本实验研究表明, 种植新葵4号后能有效降低盐碱地根际土壤pH值, 显著提高土壤中碱解氮、速效磷和速效钾的含量, 并且显著提高根际土壤脲酶和磷酸酶的活性, 对盐碱地根际微环境的养分循环有积极意义。种植四个品种的油葵均显著 (72 h, P<0.05) 提高了盐碱地土壤微生物对总碳源的利用率, 显著提高土壤微生物功能多样性指数 (H、D、S), 且以种植新葵4号的提高效果最为显著。主成分分析和聚类分析表明, 种植新葵4号后其根际土壤微生物对碳源的利用能力最强。在盐碱地中种植不同品种的油葵能有效提高相关土壤理化性质和酶活, 促进土壤养分循环和微生物代谢, 优化土壤微生物群落功能多样性, 其中以新葵4号的效果最为显著, 建议在盐碱地的改良过程中优先选择种植新葵4号。
致谢: 感谢新疆联强农业资产经营有限责任公司对本研究的支持。| [1] | 杨劲松. 中国盐渍土研究的发展历程与展望. 土壤学报, 2008, 45(5): 837–845. |
| [2] | 田长彦, 周宏飞, 刘国庆. 21世纪新疆土壤盐渍化调控与农业持续发展研究建议. 干旱区地理, 2000, 23(2): 177–181. |
| [3] | 牛东玲, 王启基. 盐碱地治理研究进展. 土壤通报, 2002, 33(6): 449–455. |
| [4] | 王斌, 马兴旺, 单娜娜, 杨涛, 刘骅, 牛新湘. 新疆盐碱地土壤改良剂的选择与应用. 干旱区资源与环境, 2014, 28(7): 111–115. |
| [5] | 高伟, 邵玉翠, 杨军, 高贤彪. 盐碱地土壤改良剂筛选研究. 中国农学通报, 2011, 27(21): 154–160. |
| [6] | 吕晓, 徐慧, 李丽, 赵雲泰. 盐碱地农业可持续利用及其评价. 土壤, 2012, 44(2): 203–207. |
| [7] | 林学政, 陈靠山, 何培青, 沈继红, 黄晓航. 种植盐地碱蓬改良滨海盐渍土对土壤微生物区系的影响. 生态学报, 2006, 26(3): 801–807. |
| [8] | 枚德新, 张德顺, 王振. 滨海盐碱地生态修复现状及趋势. 中国农学通报, 2013, 29(5): 167–171. |
| [9] | 王景艳, 邓力群, 隆小华, 刘玲, 刘兆普. 滨海盐渍化土壤引种油葵的试验研究. 土壤, 2008, 40(1): 121–124. |
| [10] | 郑青松, 陈刚, 刘玲, 刘兆普, 徐文君. 盐胁迫对油葵种子萌发和幼苗生长及离子吸收、分布的效应. 中国油料作物学报, 2005, 27(1): 60–64. |
| [11] | 石必显, 雷中华, 向理军. 利用盆栽试验对4个油葵品种的耐盐性鉴定. 新疆农业科学, 2010, 47(3): 523–526. |
| [12] | 杜社妮, 白岗栓, 于健, 任志宏. 沙封覆膜种植孔促进盐碱地油葵生长. 农业工程学报, 2014, 05: 82–90. |
| [13] | 穆俊丽, 李建科, 杨静慧, 付志芳, 桂毓, 何斌琼, 刘太林. 不同油葵品种种子萌发期的耐盐性研究. 北方园艺, 2009(5): 26–30. |
| [14] | 陈玉梁, 裴怀弟, 石有太, 陈炳东, 王蒂. 温度和盐胁迫对油葵种子萌发及幼苗生长的影响. 中国油料作物学报, 2011, 33(4): 374–378. |
| [15] | 崔云玲, 王生录, 陈炳东, 杨思存, 吕晓东. 不同品种油葵对盐胁迫响应研究. 土壤学报, 2011, 48(5): 1051–1058. |
| [16] | 何承刚. 油葵品种在盐碱地的生态适应性研究. 种子, 2004, 23(5): 6–7. |
| [17] | 孔东, 史海滨, 陈亚新, 张丽辉, 李延林, 王长生. 水盐胁迫对向日葵幼苗生长发育的影响. 灌溉排水学报, 2004, 23(5): 32–35. |
| [18] | Alsaadawi I S, Sarbout A K, Al-Shamma L M. Differential allelopathic potential of sunflower (Helianthus annuus L.) genotypes on weeds and wheat (Triticum aestivum L.) crop. Archives of Agronomy and Soil Science, 2012, 58(10): 1139–1148. DOI:10.1080/03650340.2011.570335 |
| [19] | Kamal J. Impact of allelopathy of sunflower (Helianthus annuus L.) roots extract on physiology of wheat (Triticum aestivum L.). African Journal of Biotechnology, 2011, 10(65): 14465–14477. |
| [20] | Kamal J. Quantification of alkaloids, phenols and flavonoids in sunflower (Helianthus annuus L.). African Journal of Biotechnology, 2011, 10(16): 3149–3151. |
| [21] | 关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986. |
| [22] | Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 2001. |
| [23] | Zak J C, Willig M R, Moorhead D L, Wildman H G. Functional diversity of microbial communities:a quantitative approach. Soil Biology and Biochemistry, 1994, 26(9): 1101–1108. DOI:10.1016/0038-0717(94)90131-7 |
| [24] | Zhang N L, Wan S Q, Li L H, Bi J, Zhao M M, Ma K P. Impacts of urea N addition on soil microbial community in a semi-arid temperate steppe in northern China. Plant and Soils, 2008, 311(1/2): 19–28. |
| [25] | Weber K P, Grove J A, Gehder M, Anderson W A, Legge R L. Data transformations in the analysis of community-level substrate utilization data from microplates. Journal of Microbiological Methods, 2007, 69(3): 461–469. |
| [26] | Larkin R P. Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles. Soil Biology and Biochemistry, 2003, 35(11): 1451–1466. DOI:10.1016/S0038-0717(03)00240-2 |
| [27] | Choi K H, Dobbs F C. Comparison of two kinds of Biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. Journal of Microbiological Methods, 1999, 36(3): 203–213. |
| [28] | 哈玲津, 马媛媛, 杨静慧. 四种野生植物对天津盐碱地土壤改良效果的研究. 北方园艺, 2009(4): 181–184. |
| [29] | 刘占锋, 傅伯杰, 刘国华, 朱永官. 土壤质量与土壤质量指标及其评价. 生态学报, 2006, 26(3): 901–913. |
| [30] | García-Ruiz R, Ochoa V, Hinojosa M B, Carreira J A. Suitability of enzyme activities for the monitoring of soil quality improvement in organic agricultural systems. Soil Biology and Biochemistry, 2008, 40(9): 2137–2145. DOI:10.1016/j.soilbio.2008.03.023 |
| [31] | 靳正忠, 雷加强, 徐新文, 李生宇. 沙土微生物多样性与土壤肥力质量的咸水滴灌效应. 生态学报, 2014, 34(13): 3720–3727. |
| [32] | 解丽娜, 贡璐, 朱美玲, 刘曾媛. 塔里木盆地南缘绿洲土壤酶活性与理化因子相关性. 环境科学研究, 2014, 27(11): 1306–1313. |
| [33] | 李志萍, 吴福忠, 杨万勤, 徐振锋, 苟小林, 熊莉, 殷睿, 黄莉. 川西亚高山森林林窗不同时期土壤转化酶和脲酶活性的特征. 生态学报, 2015, 35(12): 3919–3925. |
| [34] | Alkorta I, Aizpurua A, Riga P, Albizu I, Amézaga I, Garbisu C. Soil enzyme activities as biological indicators of soil health. Review of Environmental Health, 2003, 18(1): 65–73. |
| [35] | 王娟, 谷雪景, 赵吉. 羊草草原土壤酶活性对土壤肥力的指示作用. 农业环境科学学报, 2006, 25(4): 934–938. |
| [36] | 吴林坤, 林向民, 林文雄. 根系分泌物介导下植物-土壤-微生物互作关系研究进展与展望. 植物生态学报, 2014, 38(3): 298–310. |
| [37] | 何欢, 王占武, 胡栋, 张翠绵, 李洪涛, 贾楠. 根系分泌物与根际微生物互作的研究进展. 河北农业科学, 2011, 15(3): 69–73. |
| [38] | Garland J L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiology Ecology, 1997, 24(4): 289–300. |
| [39] | Preston-Mafham J, Boddy L, Randerson P F. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles-a critique. FEMS Microbiology Ecology, 2002, 42(1): 1–14. |
| [40] | Cookson W R, Murphy D V, Roper M M. Characterizing the relationships between soil organic matter components and microbial function and composition along a tillage disturbance gradient. Soil Biology and Biochemistry, 2008, 40(3): 763–777. DOI:10.1016/j.soilbio.2007.10.011 |
| [41] | Bünemann E K, Bossio D A, Smithson P C, Frossard E, Oberson A. Microbial community composition and substrate use in a highly weathered soil as affected by crop rotation and P fertilization. Soil Biology and Biochemistry, 2004, 36(6): 889–901. DOI:10.1016/j.soilbio.2004.02.002 |
| [42] | Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology, 1991, 57(8): 2351–2359. |
| [43] | 孟庆杰, 许艳丽, 李春杰, 韩晓增, 裴希超. 不同植被覆盖对黑土微生物功能多样性的影响. 生态学杂志, 2008, 27(7): 1134–1140. |
| [44] | 鲍艳, 胡振琪, 柏玉, 郭瑞珊. 主成分聚类分析在土地利用生态安全评价中的应用. 农业工程学报, 2006, 22(8): 87–90. |
| [45] | 周丽霞, 丁明懋. 土壤微生物学特性对土壤健康的指示作用. 生物多样性, 2007, 15(2): 162–171. |
| [46] | Miya R K, Firestone M K. Phenanthrene-degrader community dynamics in rhizosphere soil from a common annual grass. Journal of Environmental Quality, 2000, 29(2): 584–592. |
| [47] | 龙健, 黄昌勇, 滕应, 姚槐应. 矿区废弃地土壤微生物及其生化活性. 生态学报, 2003, 23(3): 496–503. |
| [48] | 张丽莉, 张玉兰, 陈利军, 武志杰. 稻-麦轮作系统土壤糖酶活性对开放式CO2浓度增高的响应. 应用生态学报, 2004, 15(6): 1019–1024. |