 2017, Vol. 37
2017, Vol. 37文章信息
- 曹阳, 高梅香, 张雪萍, 董承旭
- CAO Yang, GAO Meixiang, ZHANG Xueping, DONG Chengxu.
- 黑龙江省不同纬度梯度农田大型土壤动物群落分布特征
- Distribution characteristics of soil macro-faunal communities along a latitudinal gradient in farmland of Heilongjiang Province
- 生态学报. 2017, 37(5): 1677-1687
- Acta Ecologica Sinica. 2017, 37(5): 1677-1687
- http://dx.doi.org/10.5846/stxb201510142073
-
文章历史
- 收稿日期: 2015-10-15
- 网络出版日期: 2016-07-13





2. 黑龙江省普通高等学校地理环境遥感监测重点实验室, 哈尔滨师范大学, 哈尔滨 150025
2. Key Laboratory of Remote Sensing Monitoring of Geographic Environment, College of Heilongjiang Province, Harbin Normal University, Harbin 150025, China
土壤动物数量丰富, 分布广泛, 处于整个土壤食物链的顶端, 是农田生态系统重要的分解者, 在分解残体、改变土壤理化性质、土壤形成与发育、促进土壤物质循环与能量转化等方面起着关键的作用, 并且能够对环境的变化做出迅速的反应[1-5]。研究不同纬度梯度农田大型土壤动物群落空间分布特征, 分析农田大型土壤动物对气候变化的响应规律, 是全球变化背景下进行生物多样性保护的重要基础, 有助于人们在区域尺度揭示地下生态系统的生物多样性格局特征, 同时也是进行农田生态系统结构功能管理的重要依据。
大量研究表明, 土壤动物的研究主要集中在重金属等导致的农田污染对土壤动物的影响, 以及气候变化和人为干扰下生态系统退化对土壤动物的影响。21世纪以后, 对土壤动物研究范围有了进一步扩展, 涉及到各种自然灾害、旅游活动和全球变化等对土壤动物群落的影响等。不同土地利用方式下土壤动物的群落组成和数量变化、生态特征和动态变化、多样性及其评价指标分析、大型土壤动物与环境因子的相互关系等也已在国内广泛展开, 部分研究结果表明, 土壤动物随纬度的增高数量变少[6-11]。近年来, 有关土壤动物对环境的指示作用和修复作用也时常有报道[12-14], 国内外对生物多样性的纬度梯度分布规律及物种丰富度有所研究, 部分学者对不同纬度引起的气候变量与物种的丰富度之间的关系等进行了研究[15-18], 但对不同纬度大型土壤动物的群落分布研究甚少。张雪萍等[19-25]对塔河、带岭、帽儿山的土壤动物做过大量细致的研究, 但都是对某一地区样地的土壤动物进行研究, 对不同纬度梯度之间大型土壤动物的群落组成变化研究较少[5, 26-29]。
本研究目的在于探究由不同纬度而引起的气候变化下, 大型土壤动物的区系组成、分布特征和规律及其影响机制, 对于黑龙江省不同纬度大型土壤动物区系组成和生态地理分布变化规律, 及其在生态系统中的功能作用具有重要的意义, 以期反映黑龙江省不同纬度的大型土壤动物的群落特征, 为区域尺度农田生态系统土壤动物空间格局研究奠定基础, 也为区域尺度生物多样性维持机制研究提供数据支持。
1 研究区自然概况试验地点设在黑龙江省, 按照纬度从高到低选取具有代表性的研究区, 分别为黑龙江省东部山地 (帽儿山)—小兴安岭 (带岭)—大兴安岭 (塔河)。塔河县地处黑龙江省北部、大兴安岭伊勒呼里山北坡, 地理坐标为52°09′—53°23′N, 123°19′—125°48′E, 平均海拔约为600m, 属寒温带大陆性季风气候, 冬季漫长而寒冷, 夏季短暂而湿热, 年平均气温-2.4℃, 年降水量为428mm, 年无霜期在80—100d, 地带性土壤为棕色针叶林土。带岭研究区位于黑龙江省伊春市带岭区境内, 地理位置为:47°10′—47°14′N, 128°53′—128°55′ E, 本区地处小兴安岭山脉的南段达里带岭支脉的东坡, 平均海拔约为400m。本区具有明显的温带大陆性季风气候特征。年平均气温1.4℃, 年平均最高气温7.5℃, 年平均最低气温-6.6℃, 年平均降水量为676mm。地带性土壤为暗棕色森林土为主, 草甸土、沼泽土和泥炭土少量。帽儿山研究地点设在东北林业大学帽儿山实验林场, 帽儿山位于东北东部山地张广才岭余脉, 位于黑龙江省尚志市境内, 地理位置为45°20′—45°25′N, 127°30′—127°34′E, 主要为低山丘陵, 地势北高南低, 平均海拔为300m, 本区气候属于大陆性季风气候。年平均气温2.7℃, 最冷月平均气温为-19.7℃, 最热月平均气温为20.9℃, 极端最高温度是31.8℃, 极端最低温度是-31.9℃, 暗棕壤是本地区地带性土壤[16-18]。
2 研究方法 2.1 野外研究方法2011年8月分别在塔河、带岭和帽儿山随机选择3块施用无机化肥的玉米农田样地, 3块样地彼此之间至少间隔50m以上的距离。在每个纬度区域, 首先在每个农田样地中分别随机设置4个5m×5m的样方, 然后在每个样方中随机选取4个面积为50cm×50cm的采样点作为重复。使用铁锹沿土壤剖面获得每个采样点0—5、5—10、10—15cm的大型土壤动物群落, 每个纬度农田生态系统分别获得48个样品, 即3层×4采样点×4样方。采用手捡法对大型土壤动物样品进行就地分拣, 然后放入75%酒精中固定, 带回实验室内进行分类鉴定。
在调查大型土壤动物样品的同时, 获得土壤理化性质分析样品。在每个纬度区域, 分别在选择的3个农田样地中随机选择2个样方, 在该大型土壤动物的采样点附近, 使用小铁锹沿着3个层次 (0—5、5—10、10—15cm) 分别挖取10cm×10cm的土方, 将土方放入自封袋带回室内待测土壤理化性质, 同时使用铝盒获得测量土壤含水量的样品。在每个纬度农田生态系统, 分别获得土壤理化性状样品18个, 即3层×2采样点×3样地。

|
| 图 1 不同纬度梯度样地分布 Fig. 1 Distribution of sites in different latitude gradient |
依据尹文英的《中国土壤动物检索图鉴》, 在显微镜下对大型土壤动物进行分类鉴定, 物种鉴定到科, 部分物种鉴定到目, 将成虫和幼虫分开计数, 在本文分析过程中只统计成虫。测量土壤pH值、有机质、全氮、全磷、速效磷和速效钾6个主要的理化指标[30]。采用电位法测定土壤pH值, 采取丘林法测定土壤有机质, 采取重铬酸钾-硫酸消化法测定土壤全氮含量, 采用SMT方法测定全磷含量, 采用0.5mol/L NaHCO3浸提-钼锑抗比色法测定土壤速效磷含量, 采取双酸浸提、钼锑抗比色法测定速效钾。土壤环境特征见表 1。
| 土壤特性 Soil characteristics | 塔河Tahe | 带岭Dailing | 帽儿山Maoer Mountain | ||||||||
| 0—5cm | 5—10cm | 10—15cm | 0—5cm | 5—10cm | 10—15cm | 0—5cm | 5—10cm | 10—15cm | |||
| 速效钾AK/(g/kg) | 0.059±0.005 | 0.054±0.005 | 0.053±0.005 | 0.062±0.003 | 0.058±0.003 | 0.056±0.005 | 0.042±0.292 | 0.045±0.003 | 0.043±0.003 | ||
| 全磷TP/(g/kg) | 27.801±2.957 | 29.200±2.841 | 28.532±3.675 | 28.004±2.600 | 27.576±3.353 | 27.550±3.475 | 17.737±2.259 | 16.839±2.298 | 15.746±2.181 | ||
| 速效磷AP/(g/kg) | 1.700±0.519 | 1.656±0.422 | 1.333±0.362 | 1.639±0.431 | 1.925±0.398 | 1.376±0.370 | 0.678±0.519 | 0.632±0.256 | 0.559±0.243 | ||
| 全氮TN/(g/kg) | 0.005±0.001 | 0.004±0.001 | 0.004±0.002 | 0.003±0.000 | 0.003±0.000 | 0.003±0.000 | 0.003±0.000 | 0.002±0.000 | 0.003±0.000 | ||
| 有机质SOM/(g/kg) | 0.090±0.013 | 0.087±0.012 | 0.082±0.014 | 0.069±0.003 | 0.069±0.003 | 0.066±0.003 | 0.055±0.005 | 0.047±0.003 | 0.045±0.003 | ||
| pH | 5.969±0.346 | 5.941±0.341 | 6.031±0.392 | 6.204±0.255 | 6.200±0.252 | 6.290±0.230 | 6.391±0.233 | 6.430±0.246 | 6.571±0.221 | ||
多样性指数:选用Shannon-wiener多样性指数 (H′)、Pielou均匀度指数 (E)、Simpson优势度指数 (C) 和Margalef丰富度指数 (D) 对土壤动物群落结构多样性进行分析。计算公式如下[21]:
Shannon-wiener多样性指数
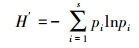
|
(1) |
Pielou均匀度指数
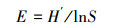
|
(2) |
Simpson优势度指数
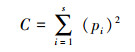
|
(3) |
Margalef丰富度指数
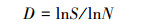
|
(4) |
式中, pi指i类群在群落中的个体比例, 即pi=ni/N, ni为样区内第i个类群的个体数量, N为样区内所有类群的个体数量, S为样区内类群的数目。
所有数据的分析与制图采用SPSS 19.0、Origin 7.0和CANOCO 5.0软件。利用SPSS 19.0软件中的单因素方差分析 (One way ANOVA, LSD) 比较不同纬度大型土壤动物个体数、类群数差异的显著性, 双变量相关分析 (Bivariate correlation analysis) 方法对环境因子与土壤动物群落指标的相关性进行分析;用CANOCO 5.0软件中的典范对应分析 (Canonical correspondence analysis, CCA) 对本研究中大型土壤动物数量及优势类群和常见类群的数量与土壤环境因子进行相关分析。
3 结果与分析 3.1 大型土壤动物群落组成本研究共获得大型土壤动物35类2339只, 隶属于2门6纲14目35科, 平均密度为778.33只/m2, 其中优势类群 (>总个体数10%)4类, 为线蚓科 (Enchytraeidae)、正蚓科 (Lumbricidae)、步甲科 (Carabida)、隐翅虫科 (Staphylinidae), 占总个体数的58.84%;常见类群 (占总个体数1%—10%)8类, 为蚁科 (Formicidae)、蜘蛛目 (Araneida)、地蜈蚣目 (Geophilomorpha)、石蜈蚣目 (Lithobiomorpha)、马陆目 (Juliformia)、叩甲科 (Elateridae)、金龟甲科 (Scarabaeoidae)、蝙蝠蛾科 (Hepialidae), 占总个体数的32.79%;稀有类群 (<总个体数1%)23类, 为象甲科 (Curculionidae)、舞虻科 (Empidadae)、鹬虻科 (Rhagionidae)、大蚊科 (Tipulidae)、叶甲科 (Ghrysomelidae)、剑虻科 (Therevidae) 等, 占总个体数的8.37%(表 2)。
| 序号 Order | 类群 Group | 塔河Tahe | 带岭Dailing | 帽儿山Maoer Mountain | 总Total | ||||||||
| 密度 Density /(只/m2) | % | 密度 Density /(只/m2) | % | 密度 Density /(只/m2) | % | 密度 Density /(只/m2) | % | 优势度 Abundence | |||||
| 1 | 线蚓科Enchytraeidae | 129.00±104.06 | 21.46 | 281.00±230.13 | 21.46 | 158.0±79.84 | 23.87 | 568.00±301.98 | 24.28 | + + + | |||
| 2 | 正蚓科Lumbricidae | 43.00±27.00 | 7.15 | 210.00±112.98 | 19.52 | 30.0±24.68 | 4.53 | 283.00±143.37 | 12.10 | + + + | |||
| 3 | 蜘蛛目Araneida | 65.00±14.55 | 10.82 | 51.00±13.99 | 4.74 | 44.0±9.93 | 6.65 | 160.00±21.73 | 6.84 | + + | |||
| 4 | 地蜈蚣目Geophilomorpha | 7.00±4.12 | 1.16 | 130.00±63.93 | 12.08 | 69.0±20.55 | 10.42 | 206.00±86.79 | 8.81 | + + | |||
| 5 | 石蜈蚣目Lithobiomorpha | 16.00±10.95 | 2.66 | 10.00±3.46 | 0.93 | 28.0±21.42 | 4.23 | 54.00±24.08 | 2.31 | + + | |||
| 6 | 马陆目Juliformia | 28.00±26.68 | 2.60 | 9.00±9.00 | 1.35 | 37.00±35.68 | 1.58 | + + | |||||
| 7 | 鼠妇科Porcellio spinicornis | 23.00±23.00 | 3.83 | 23.00±23.00 | 0.98 | + | |||||||
| 8 | 步甲科Carabidae | 83.00±32.80 | 13.81 | 110.00±32.60 | 10.22 | 54.00±34.12 | 8.16 | 247.00±58.13 | 10.56 | + + | |||
| 9 | 隐翅虫科Staphylinidae | 152.00±56.31 | 25.29 | 93.00±39.41 | 8.64 | 35.00±12.48 | 5.29 | 280.00±48.58 | 11.97 | + + | |||
| 10 | 象甲科Curculionidae | 1.00±1.00 | 0.17 | 1.00±1.00 | 0.04 | + | |||||||
| 11 | 叩甲科Elateridae | 3.00±1.91 | 0.50 | 24.00±5.65 | 2.23 | 3.00±3.00 | 0.45 | 30.00±6.22 | 1.28 | + + | |||
| 12 | 锹甲科Lucanidae | 2.00±1.15 | 0.19 | 2.00±1.15 | 0.30 | 4.00±0.00 | 0.17 | + | |||||
| 13 | 金龟甲科Scarabaeoidae | 3.00±3.00 | 0.50 | 28.00±16.25 | 2.60 | 29.00±8.70 | 4.38 | 60.00±21.23 | 2.57 | + + | |||
| 14 | 叶甲科Ghrysomelidae | 1.00±1.00 | 0.09 | 1.00±1.00 | 0.04 | + | |||||||
| 15 | 虎甲科Cicindelidae | 1.00±1.00 | 0.17 | 4.00±2.83 | 0.37 | 13.00±5.51 | 1.96 | 18.00±4.76 | 0.77 | + | |||
| 16 | 拟步甲科Tenebrionidae | 1.00±1.00 | 0.09 | 1.00±1.00 | 0.04 | + | |||||||
| 17 | 葬甲科Silphidae | 9.00±9.00 | 1.36 | 9.00±9.00 | 0.38 | + | |||||||
| 18 | 冬大蚊科Trichoceridae | 1.00±1.00 | 0.17 | 1.00±1.00 | 0.04 | + | |||||||
| 19 | 毛蚊科Bibionidae | 2.00±2.00 | 0.33 | 1.00±1.00 | 0.09 | 1.00±1.00 | 0.15 | 4.00±1.63 | 0.17 | + | |||
| 20 | 蚤蝇科Phoridae | 2.00±1.15 | 0.33 | 2.00±1.15 | 0.09 | + | |||||||
| 21 | 大蚊科Tipulidae | 3.00±1.91 | 0.50 | 10.00±8.72 | 0.93 | 3.00±1.91 | 0.45 | 16.00±9.93 | 0.68 | + | |||
| 22 | 舞虻科Empidadae | 9.00±5.26 | 1.50 | 4.00±2.83 | 0.37 | 9.00±7.72 | 1.36 | 22.00±9.59 | 0.94 | + | |||
| 23 | 鹬虻科Rhagionidae | 3.00±3.00 | 0.50 | 2.00±1.15 | 0.19 | 5.00±3.79 | 0.21 | + | |||||
| 24 | 长足虻科Dolichopodadae | 2.00±2.00 | 0.33 | 2.00±2.00 | 0.19 | 1.00±1.00 | 0.15 | 5.00±5.00 | 0.21 | + | |||
| 25 | 剑虻科Therevidae | 4.00±2.83 | 0.67 | 4.00±2.83 | 0.17 | + | |||||||
| 26 | 食木虻科Xylophagiae | 14.00±14.00 | 2.11 | 14.00±14.00 | 0.60 | + | |||||||
| 27 | 蚁科Formicidae | 32.00±10.71 | 5.32 | 45.00±32.10 | 4.18 | 111.00±32.10 | 16.78 | 188.00±13.95 | 8.04 | + + | |||
| 28 | 土蝽科Cydnidae | 8.00±1.63 | 1.33 | 3.00±1.91 | 0.28 | 3.00±1.00 | 0.45 | 14.00±1.15 | 0.60 | + | |||
| 29 | 椿象若虫Hemiptera | 3.00±1.00 | 0.50 | 7.00±2.52 | 0.65 | 11.00±9.71 | 1.66 | 21.00±12.15 | 0.90 | + | |||
| 30 | 蝙蝠蛾科Hepialidae | 5.00±3.79 | 0.83 | 6.00±4.76 | 0.56 | 21.00±13.40 | 3.17 | 32.00±17.81 | 1.37 | + + | |||
| 31 | 尺蛾科Geometridae | 1.00±1.00 | 0.15 | 1.00±1.00 | 0.04 | + | |||||||
| 32 | 天蛾科Sphingidae | 2.00±2.00 | 0.19 | 2.00±2.00 | 0.09 | + | |||||||
| 33 | 球螋科Forficulidae | 1.00±1.00 | 0.09 | 3.00±1.91 | 0.45 | 4.00±2.31 | 0.17 | + | |||||
| 34 | 蝼蛄科Gryllotalpidae | 20.00±14.79 | 1.86 | 1.00±1.00 | 0.15 | 21.00±15.78 | 0.90 | + | |||||
| 35 | 蜢总科Eumastacoidea | 1.00±2.00 | 0.17 | 1.00±1.00 | 0.04 | + | |||||||
| 总密度Total density | 601.00±317.98 | 100 | 1076.00±628.85 | 100 | 662.00±325.03 | 100 | 2339.00±893.62 | 100 | |||||
| 总类群数Total group number | 25 | 26 | 25 | 35 | |||||||||
| + + +:优势类群 (>10%);+ +:常见类群 (1%—10%);+:稀有类群 ( < 1%) | |||||||||||||
塔河实验样地共获得大型土壤动物25类、601只, 隶属于2门5纲13目。其中优势类群4类, 为隐翅虫科 (Staphylinidae)、线蚓科 (Enchytraeidae)、步甲科 (Carabidae) 和蜘蛛目 (Araneida), 占总个体数的71.38%, 常见类群有正蚓科 (Lumbricidae)、地蜈蚣目 (Geophilomorpha)、石蜈蚣目 (Lithobiomorpha) 等7类, 占总个体数的22.96%, 稀有类群有蜢总科 (Eumastacoidea)、象甲科 (Curculionidae)、虎甲科 (Cicindelidae) 等14类, 占个体数的5.66%;带岭实验样地共获得大型土壤动物26类、1076只, 隶属于2门5纲13目。其中优势类群4类, 为线蚓科 (Enchytraeidae)、正蚓科 (Lumbricidae)、地蜈蚣科 (Geophilomorpha) 和步甲科 (Carabidae), 占总个体数的67.94%, 常见类群有蜘蛛目 (Araneida)、马陆目 (Juliformia)、隐翅虫科 (Staphylinidae) 等7类, 占总个体数的26.86%, 稀有类群有石蜈蚣目 (Lithobiomorpha)、锹甲科 (Lucanidae)、叶甲科 (Ghrysomelidae) 等15类, 占总个体数的5.2%;帽儿山实验样地共获得大型土壤动物25类、662只, 隶属于2门5纲13目。其中优势类群3类, 为线蚓科 (Enchytraeidae)、蚁科 (Formicidae) 和地蜈蚣科 (Geophilomorpha), 占总个体数的52.06%, 常见类群有正蚓科 (Lumbricidae)、蜘蛛目 (Araneida)、金龟甲科 (Scarabaeoidae) 等13类, 占总个体数的46.25%;稀有类群有锹甲科 (Lucanidae)、叩甲科 (Elateridae)、毛蚊科 (Bibionidae) 等, 占总个体数的1.69%(表 2)。
同一物种在不同纬度农田生境中的分布特征存在差异, 如线蚓科 (Enchytraeidae) 在不同纬度中均为优势类群, 毛蚊科 (Bibionidae)、长足虻科 (Dolichopodadae) 在不同纬度中均为稀有类群, 正蚓科 (Lumbricidae) 在塔河和帽儿山样地中为常见类群, 在带岭样地中为优势类群, 蜘蛛目 (Araneida) 在带岭和帽儿山样地为常见类群, 在塔河样地为优势类群, 石蜈蚣目 (Lithobiomorpha) 在塔河和帽儿山样地中为常见类群, 在带岭样地中为稀有类群;马陆目 (Juliformia) 在带岭和帽儿山样地中为常见类群, 而在塔河样地没有分布等 (表 2)。
3.2 不同纬度的土壤动物的水平分布特征带岭样地的大型土壤动物的个体密度和丰富度 (即类群数) 均最多, 与塔河和帽儿山样地无显著差异 (表 3)。但少部分土壤动物在不同样地中的个体密度存在不同的差异, 如地蜈蚣目 (Geophilomorpha) 在塔河样地中有7.00只/m2, 在带岭样地中高达130.00只/m2, 存在显著差异, 而毛蚊科 (Bibionidae)(1—2只/m2)、长足虻科 (Dolichopodadae)(1—2只/m2) 等在不同样地中的个体密度差异不显著。
| 项目Item | N/(只/m2) | S | H′ | E | C | D |
| 样地Sites | ||||||
| 塔河Tahe | 601.00±317.98 | 14.00±2.94 | 1.90±0.11 | 0.13±0.02 | 0.21±0.02 | 0.55±0.02 |
| 带岭Dailing | 1076.00±628.85 | 16.75±1.03 | 1.99±0.16 | 0.71±0.05 | 0.21±0.05 | 0.53±0.04 |
| 帽儿山Maoer Mountain | 662.00±325.03 | 15.50±1.44 | 2.11±0.13 | 0.77±0.03 | 0.18±0.03 | 0.54±0.02 |
| 土层Siol layer | ||||||
| 0—5cm | 395.32±77.44 | 11.67±0.99 | 1.86±0.08 | 0.78±0.02 | 0.21±0.02 | 0.57±0.02 |
| 5—10cm | 282.00±45.68 | 11.00±0.92 | 1.90±0.77 | 0.81±0.02 | 0.21±0.02 | 0.63±0.02 |
| 10—15cm | 156.32±37.80 | 8.17±0.90 | 1.60±0.09 | 0.80±0.03 | 0.27±0.03 | 0.61±0.02 |
| 同一列不同字母表示显著性差异组别 (P < 0.05) N:个体密度;S:类群数;H′:多样性指数;E:均匀度指数;C:优势度指数;D:丰富度指数 | ||||||
同一种类群在不同样地中优势度是不同的, 如隐翅虫科 (Staphylinidae) 在塔河样地为优势类群, 在带岭和帽儿山样地为常见类群。不同纬度的3个样地共有类群18类, 线蚓科 (Enchytraeidae) 为3个样地共有优势类群, 蜢总科 (Eumastacoidea) 为塔河样地的特有类群, 天蛾科 (Sphingidae) 只出现在带岭样地, 帽儿山样地才有尺蛾科 (Geometridae)、葬甲科 (Silphidae)(表 2)。
通过单因素方差分析方法可知, 3个不同样地的多样性指数均无显著差异, 个体密度和丰富度的大小顺序均为:带岭>帽儿山>塔河, H′指数和E指数均表现为:帽儿山>带岭>塔河, C指数表现为塔河和带岭最高, 帽儿山最少, D指数则是塔河最多, 其次为帽儿山和带岭 (表 3)。
3.3 不同纬度的大型土壤动物的垂直分布特征大型土壤动物个体密度和丰富度在不同纬度的垂直方向上均无显著差异。通过分析, 多样性指数也均无显著性差异 (表 3)。个体密度在土层分布上呈现显著的表聚性, 但不同纬度的同土层变化情况不同 (图 2)。在0—5cm层, 大型土壤动物的个体密度呈现带岭样地最高, 帽儿山样地最低, 塔河样地居中, 丰富度大小顺序依次为带岭>帽儿山>塔河;5—10cm层, 大型土壤动物的个体密度带岭样地最多、塔河和帽儿山样地次之, 丰富度为帽儿山>带岭>塔河;10—15cm层大型土壤动物的个体密度和丰富度均为带岭样地最多, 塔河样地最低, 帽儿山样地居中。各土层的大型土壤动物均未随纬度的变化而呈现有规律性的变化。

|
| 图 2 不同纬度农田生境大型土壤动物个体密度和类群数的垂直分布 Fig. 2 Vertical distribution of individual density and species number of soil macro-fauna in farmland habitats from different latitudes |
不同纬度样地大型土壤动物在同土层的类群组成及分布存在差异, 有些类群在各个土层均为优势物种、常见物种或稀有物种, 有些土壤动物类群只分布在特定土层, 有些土壤动物不呈表聚性等。如线蚓科 (Enchytraeidae) 为3个纬度样地各层的优势种, 毛蚊科 (Bibionidae)、叶甲科 (Ghrysomelidae)、锹甲科 (Lucanidae)、拟步甲科 (Tenebrionidae)、剑虻科 (Therevidae) 和蚤蝇科 (Phoridae) 在3个纬度样地中各土层均为稀有物种。0—5cm层, 石蜈蚣目 (Lithobiomorpha) 在塔河和带岭样地为常见物种, 在帽儿山样地为优势物种, 正蚓科 (Lumbricidae) 在塔河和帽儿山样地为常见物种, 在带岭样地为优势物种, 金龟甲科 (Scarabaeoidae) 在塔河样地为稀有物种, 在带岭和帽儿山样地为常见物种。5—10cm层, 蜘蛛目 (Araneida) 在3个纬度样地均为常见物种, 石蜈蚣目 (Lithobiomorpha) 在塔河和带岭样地为常见物种, 在帽儿山样地为优势物种, 正蚓科 (Lumbricidae) 在塔河和帽儿山样地为常见物种, 在带岭样地为优势物种。10—15cm层, 步甲科 (Carabidae) 和蜘蛛目 (Araneida) 在3个纬度样地均为常见物种, 隐翅虫科 (Staphylinidae) 在塔河样地中为优势种, 在带岭和帽儿山样地为常见物种等等。一些类群只出现在特定土层, 塔河样地的象甲科 (Curculionidae)、鹬虻科 (Rhagionidae) 等, 帽儿山样地的蝼蛄科 (Gryllotalpidae) 等只在0—5cm土层有分布;塔河样地的叩甲科 (Elateridae) 和蜢总科 (Eumastacoidea) 等, 带岭样地的鹬虻科 (Rhagionidae) 和毛蚊科 (Bibionidae), 帽儿山样地的长足虻科 (Dolichopodadae) 和尺蛾科 (Geometridae) 只出现在5—10cm土层。不同类群土壤动物个体密度在垂直方向上变化规律是不同的, 如步甲科 (Carabidae) 等物种是随土层深度的增加而减少的, 带岭样地中的葬甲科 (Silphidae) 在各土层分布较均匀, 带岭和帽儿山样地的叩甲科 (Elateridae) 等的个体密度呈递增趋势。塔河样地的舞虻科 (Empidadae)、金龟甲科 (Scarabaeoidae) 在0—5cm土层是稀有物种, 在10—15cm土层却是常见物种, 带岭样地的叶甲科 (Ghrysomelidae) 和球螋科 (Forficulidae) 出现在10—15cm土层, 帽儿山样地的马陆目 (Juliformia) 在5—10cm土层和10—15cm土层为常见物种, 在0—5cm土层却是稀有类群, 这些类群不呈现表聚性 (表 2)。
3.4 大型土壤动物群落指标与土壤环境因子的相关分析通过双变量相关分析得出结果显示:塔河样地的丰富度和D指数与土壤环境因子呈正相关;个体密度除与速效磷和全氮呈负相关、H′指数和C指数除与pH值呈负相关外与其他土壤环境因子均呈正相关;E指数与pH值呈正相关, 与其他土壤环境因子均呈负相关。带岭样地的丰富度与速效钾、速效磷、有机质呈负相关, 与全氮、全磷和pH值呈正相关;个体密度与pH值呈正相关, 与其他土壤环境因子均呈负相关;H′指数、E指数与全氮和有机质呈正相关, C指数与全氮和有机质呈负相关;D指数与土壤环境因子均呈正相关。帽儿山样地的丰富度与速效钾呈正相关、个体密度与速效钾呈负相关, 而丰富度和个体密度与全氮相关结果与上相反;H′指数和D指数除与全氮呈负相关外, 与其他土壤环境因子均呈正相关;E指数与全磷和全氮呈负相关, 与其他土壤环境因子呈正相关;C指数除与全氮呈正相关外, 与其他土壤环境因子均呈负相关。但是, 3个样地大型土壤动物群落指标与土壤环境因子均无显著的相关性。
利用CCA分析不同纬度大型土壤动物类群的数量与土壤环境因子之间的关系 (图 3)。图中塔河样地的Axis1与Axis2的Eigenvalues值分别为0.2274和0.1137, 带岭样地的Axis1与Axis2的Eigenvalues值分别为0.2048和0.1087, 帽儿山样地的Axis1与Axis2的Eigenvalues值分别为0.2788和0.1317。塔河样地中的蜘蛛目 (Araneida)、隐翅虫科 (Staphylinidae) 等, 带岭样地中的线蚓科 (Enchytraeidae)、地蜈蚣目 (Geophilomorpha) 和石蜈蚣目 (Lithobiomorpha) 等, 帽儿山样地中线蚓科 (Enchytraeidae)、地蜈蚣目 (Geophilomorpha) 和蚁科 (Formicidae) 等分布广泛, 也是各样地中的优势类群和常见类群, 集中在图的中心位置;塔河样地的舞虻科 (Empidadae) 和带岭样地的毛蚊科 (Bibionidae) 与全氮相关性较大, 带岭样地的蜘蛛目 (Araneida) 和叩甲科 (Elateridae)、帽儿山样地的尺蛾科 (Geometridae) 与土壤有机质相关性较大。

|
| 图 3 不同纬度农田生境大型土壤动物对应分析二维排序图 Fig. 3 Correspondence analysis ordination bassed on soil macro-fauna in famland habitats from different latitudes AK:速效钾;TP:全磷;AP:速效磷;TN:全氮;SOM有机质; 1:线蚓科Enchytraeidae; 2:正蚓科Lumbricidae; 3:蜘蛛目Araneida; 4:地蜈蚣目Geophilomorpha; 5:石蜈蚣目Lithobiomorpha; 6:马陆目Juliformia; 7:鼠妇科Porcellio spinicorni; 8:步甲科Carabidae; 9:隐翅虫科Staphylinidae; 10:象甲科Curculionidae; 11:叩甲科Elateridae; 12:锹甲科Lucanidae; 13:金龟甲科Scarabaeoidae; 14:叶甲科Ghrysomelidae; 15:虎甲科Cicindelidae; 16:拟步甲科Tenebrionidae; 17:葬甲科Silphidae; 18:冬大蚊科Trichoceridae; 19:毛蚊科Bibionidae; 20:蚤蝇科Phoridae; 21:大蚊科Tipulidae; 22:舞虻科; Empidadae; 23:鹬虻科Rhagionidae; 24:长足虻科Dolichopodadae; 25:剑虻科Therevidae; 26:食木虻科Xylophagiae; 27:蚁科Formicidae; 28:土蝽科Cydnidae; 29:椿象若虫科Hemiptera; 30:蝙蝠蛾科Hepialidae; 31:尺蛾科Geometridae; 32:天蛾科Sphingidae; 33:球螋科Forficulidae; 34:蝼蛄科Gryllotalpidae; 35:蜢总科Eumastacoidea |
通过对黑龙江省3个不同纬度的大型土壤动物的调查表明, 线蚓科 (Enchytraeidae) 为3个样地的共同优势类群, 表现出对不同纬度的普适性。张雪萍等[20]在大兴安岭北部森林生态系统大型土壤动物群落特征中, 线蚓科 (Enchytraeidae) 为优势类群, 与本研究塔河样地结果一致。其研究表明蜘蛛目 (Araneida)、隐翅虫科 (Staphylinidae) 和步甲科 (Carabidae) 是常见类群, 但在本研究中为优势类群, 结论不完全一致, 说明在同一地点不同生态系统中物种分布有差异。在调查中, 天蛾科 (Sphingidae) 只在带岭样地中出现, 而张雪萍、林英华等[20-22]对塔河、带岭和帽儿山土壤动物的研究中均未调查出天蛾科 (Sphingidae), 这可能是受不同环境因素或者样地位置选择的影响的缘故。本研究中, 帽儿山样地中的稀有丰富度最小, 与以往学者[21-22, 31]对帽儿山土壤动物群落结构研究相一致, 且类群种类差异也较小。由于不同地区环境的异质性有所差异, 因此3个纬度的群落以及不同土层深度均有各自的特有类群。通过单因素方差分析法可知3个样地的丰富度均无显著差异, 说明丰富度受纬度影响不大。大型土壤动物对环境变化具有一定的指示作用, 由此可见, 对于土壤动物的研究应该更加注重类群的不同纬度的分析, 基于物种不同纬度的水平土壤动物类群与环境关系的研究, 能更好的对大环境变化及发展趋势进行评价和预测。
4.2 不同纬度大型土壤动物空间分布特征不同样地微生境的变化也会引起土壤动物群落结构的细微差异, 土壤环境因子是引起土壤动物个体密度和丰富度变化的主要原因[31]。个体密度和丰富度随着纬度的升高呈先上涨后下降趋势。由于塔河的年平均气温低, 降水量少, 纬度高, 所以大型土壤动物的个体密度和丰富度均最少, 而带岭大型土壤动物的个体密度和丰富度高于帽儿山的原因有以下两点:(1) 带岭的土壤有机质、速效钾、速效磷、全磷等土壤环境因子均高于帽儿山, 且带岭的优势度指数也高于帽儿山, 综合各因素或者某些因子对其影响导致带岭的大型土壤动物个体密度和丰富度高于帽儿山。(2) 本研究于8月在野外取样, 带岭的降水集中在6—8月, 土壤动物较多;帽儿山在采样期间受环境胁迫影响较大, 如施肥量及时间的不同等原因导致帽儿山的土壤动物个体密度和丰富度减少[32-33]。不同纬度大型土壤动物丰富度、个体密度差异不显著, 主要是因为土壤动物优势类群和常见类群多具有较强的环境适应性 (图 3), 由此使不同纬度农田耕地生态系统大型土壤动物群落存在较大的稳定性, 群落结构不随纬度和环境差异而显著变化。
凋落物及其形成的腐殖质是土壤动物重要食物来源, 且具有一定保温和保湿作用, 有利于土壤动物群落的数量增长, 随深度增加, 土壤有机质养分下降 (表 1), 且通气透水性及温度随之降低, 因此, 在垂直方向上, 不同纬度的大型土壤动物个体密度及丰富度有明显表聚性特征。此次调查研究结果表明, 不同层次大型土壤动物的个体密度和丰富度下降幅度有所差异, 导致不同纬度样地大型土壤动物在土壤剖面中的分布模式存在差异。其中, 除5—10cm土层的帽儿山样地的丰富度外, 3个不同纬度样地的个体密度和丰富度均随土层深度的增加而下降, 且中、低纬度样地表现为中层和下层土壤相近, 从中层开始明显下降, 高纬度样地则表现为表层土壤和中层土壤相近, 下层土壤明显较低。类群和个体最小密度为高纬度的最深层, 类群和个体最大密度为带岭样地的表层[10]。而帽儿山5—10cm土层的丰富度最大, 可能由于8月降水丰富, 土壤表层受雨水的冲刷导致土壤动物种类减少, 且5—10cm的H′指数、E指数和D指数均高于0—5cm层, 土壤动物多样性反映土壤中食物资源和生境的多样性, 多样性指数比类群和密度更能体现群落的结构, 在理论和实践中都具有重要意义, 所以在帽儿山大型土壤动物丰富度5—10cm土层最丰富, 土壤动物垂直分布特征说明, 维持土壤表层的质量在保护土壤动物种群方面具有重要意义[34]。
4.3 不同纬度大型土壤动物与土壤理化性质相关分析3个不同纬度样地土壤动物的个体密度和丰富度的多少受土壤环境因子的影响, 通过双变量相关分析结果表明:速效钾和有机质含量多少对塔河、带岭和帽儿山样地的土壤动物的个体密度有重要的影响, 同时, 全磷、速效磷、全氮和含水量的变化对各纬度样地的个体密度和丰富度多少也起到决定性的作用。
塔河样地的丰富度与土壤环境因子呈正相关;个体密度除与速效磷和全氮呈负相关外与其他土壤环境因子呈正相关。带岭样地的丰富度与速效钾、速效磷、有机质呈负相关, 与全氮、全磷和pH值呈正相关;个体密度与pH值呈正相关, 与其他土壤环境因子呈负相关。帽儿山样地的丰富度与速效钾呈正相关、个体密度与速效钾呈负相关, 而丰富度和个体密度与全氮相关结果与上相反。但是, 3个不同纬度样地大型土壤动物的个体与土壤环境因子均无显著的相关性, 这与刘继亮等[35]在左家自然保护区大型土壤动物与土壤因子关系研究中的结果不同。CCA结果表明, 不同纬度不同类群土壤动物与土壤理化环境的相关性不同:塔河样地的舞虻科 (Empidadae) 和带岭样地的毛蚊科 (Bibionidae) 与全氮相关性较大, 带岭样地的蜘蛛目 (Araneida) 和叩甲科 (Elateridae)、帽儿山样地的尺蛾科 (Geometridae) 与土壤有机质相关性较大。不同纬度不同类群的土壤动物具有选择性吸收和富集某些元素的功能, 因此研究不同纬度土壤动物分布与生态环境的关系, 对促进土壤动物的有益发展和抑制其有害活动, 维持土壤生态系统平衡, 具有重要意义。
5 结论通过对3块不同纬度玉米农田样地的大型土壤动物研究可知, 不同纬度大型土壤动物的个体密度、丰富度和多样性指数的垂直分布基本呈现表聚性, 及随土层深度的增加而呈递减变化;大型土壤动物的个体密度和丰富度未随纬度的升高而呈现规律性的变化趋势;大型土壤动物的群落结构和多样性受纬度和土壤理化性质的影响, 在局地尺度环境因子对土壤动物的影响不容忽视, 但不同样地间的大型土壤动物的群落结构指标均无显著性差异, 说明多数类群在黑龙江省不同纬度的大环境中具有一定的适应性。
致谢: 感谢塔河、带岭、帽儿山当地农民在实验过程中给予的帮助。| [1] | 尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998. |
| [2] | 朱永恒, 赵春雨, 王宗英, 濮励杰. 我国土壤动物群落生态学研究综述. 生态学杂志, 2005, 24(12): 1477–1481. |
| [3] | 高立杰, 侯建华, 安哲, 高宝嘉. 内蒙古高原东南缘森林草原交错带土壤动物群落特征. 草业学报, 2013, 22(4): 27–34. |
| [4] | 潘林, 王文峰, 郭继勋, 孙玉坤. 扎龙自然保护区大型土壤动物的群落结构. 土壤通报, 2012, 43(4): 781–786. |
| [5] | 林恭华, 赵芳, 陈桂琛, 陈生云, 苏建平, 张同作. 青海湖北岸不同土地利用方式对大型土壤动物群落的影响. 草业学报, 2012, 21(2): 180–186. |
| [6] | 林英华, 孙家宝, 张夫道. 我国重要森林群落凋落物层土壤动物群落生态特征. 生态学报, 2009, 29(6): 2938–2944. |
| [7] | Gartner T B, Cardon Z G. Decomposition dynamics in mixed-species leaf litter. Oikos, 2004, 104(2): 230–246. DOI:10.1111/oik.2004.104.issue-2 |
| [8] | Begon M, Harper J L, Townsend C R. Ecology:individuals, populations and communities. Sunderland, Massachusettes: Sinauer Associates, Inc., 1986: 386–415. |
| [9] | Zhao H L, Li J, Liu R T, Zhou R L, Qu H, Pan C C. Effects of desertification on temporal and spatial distribution of soil macro-arthropods in Horqin sandy grassland, Inner Mongolia. Geoderma, 2014, 223-225: 62–67. DOI:10.1016/j.geoderma.2014.01.026 |
| [10] | 赵哈林, 刘任涛, 赵学勇, 张铜会. 沙漠化对科尔沁沙质草地大型土壤节肢动物群落时空变化的影响. 草地学报, 2013, 21(2): 394–400. |
| [11] | 刘任涛. 科尔沁沙地土壤动物群落分布特征. 北京: 科学出版社, 2015. |
| [12] | 赵哈林, 刘任涛, 周瑞莲, 曲浩, 潘成臣, 王燕, 李瑾. 沙漠化对科尔沁沙质草地大型土壤动物群落的影响及其成因分析. 草业学报, 2013, 22(3): 70–77. |
| [13] | 刘任涛, 赵哈林. 沙质草地土壤动物的研究进展及建议. 中国沙漠, 2009, 29(4): 656–662. |
| [14] | 宋理洪, 武海涛, 吴东辉. 我国农田生态系统土壤动物生态学研究进展. 生态学杂志, 2011, 30(12): 2898–2906. |
| [15] | Rivadeneira M M, Fernández M, Navarrete S A. Latitudinal trends of species diversity in rocky intertidal herbivore assemblages:spatial scale and the relationship between local and regional species richness. Marine Ecology Progress Series, 2002, 245: 123–131. DOI:10.3354/meps245123 |
| [16] | Qiao X J, Jabot F, Tang Z Y, Jiang M X, Fang J Y. A latitudinal gradient in tree community assembly processes evidenced in Chinese forests. Global Ecology and Biogeography, 2015, 24(3): 314–323. DOI:10.1111/geb.2015.24.issue-3 |
| [17] | Gorelick R. Species richness and the analytic geometry of latitudinal and altitudinal gradients. Acta Biotheoretica, 2008, 56(3): 197–203. DOI:10.1007/s10441-008-9048-7 |
| [18] | Fiera C, Ulrich W. Spatial patterns in the distribution of European springtails (Hexapoda:Collembola). Biological Journal of the Linnean Society, 2012, 105(3): 498–506. DOI:10.1111/bij.2012.105.issue-3 |
| [19] | 张雪萍, 黄初龙, 李景科. 赤子爱胜蚓对森林凋落物的分解效率. 生态学报, 2005, 25(9): 2427–2433. |
| [20] | 张雪萍, 黄丽荣, 姜丽秋. 大兴安岭北部森林生态系统大型土壤动物群落特征. 地理研究, 2008, 27(3): 509–518. |
| [21] | 仲伟彦, 殷秀琴, 陈鹏. 帽儿山森林落叶分解消耗与土壤动物关系的研究. 应用生态学报, 1999, 10(4): 511–512. |
| [22] | 林英华, 孙家宝, 刘海良, 张夫道, 孙龙, 金森. 黑龙江帽儿山土壤动物群落组成与多样性分析. 林业科学, 2006, 42(4): 71–77. |
| [23] | 张雪萍, 张毅, 侯威岭, 殷秀琴. 小兴安岭针叶凋落物的分解与土壤动物的作用. 地理科学, 2000, 20(6): 552–556. |
| [24] | 李娜, 张雪萍, 张利敏. 红松人工林与天然次生林大型土壤动物功能类群. 应用与环境生物学报, 2014, 20(1): 22–29. |
| [25] | 黄丽荣, 张雪萍. 大兴安岭北部森林生态系统土壤动物的功能类群及其生态分布. 土壤通报, 2008, 39(5): 1017–1022. |
| [26] | 张志罡, 孙继英, 胡波, 颜亨梅. 土壤动物研究综述. 生命科学研究, 2006, 10(4): 72–75. |
| [27] | 刘继亮, 曹靖, 张晓阳, 李世杰, 潘春林. 秦岭西部日本落叶松林大型土壤动物群落特征. 应用与环境生物学报, 2013, 19(4): 611–617. |
| [28] | 吴东辉, 张柏, 陈鹏. 长春市不同土地利用条件下大型土壤动物群落结构与组成. 动物学报, 2006, 52(2): 279–287. |
| [29] | 董炜华, 王子婷, 李锋瑞, 刘继亮. 黑河中游绿洲化过程中不同开垦年限农田大型土壤动物群落特征. 地理科学, 2014, 34(10): 1254–1261. |
| [30] | 张淑花.农田生态系统土壤动物群落结构对不同管理与利用方式的响应[D].哈尔滨:哈尔滨师范大学, 2014:19-22. |
| [31] | 傅必谦, 陈卫, 董晓晖, 邢忠民, 高武. 北京松山四种大型土壤动物群落组成和结构. 生态学报, 2002, 22(2): 215–223. |
| [32] | 朱新玉, 朱波. 不同施肥方式对紫色土农田土壤动物主要类群的影响. 中国农业科学, 2015, 48(5): 911–920. |
| [33] | 林英华, 朱平, 张夫道, 彭畅, 高洪军, 刘淑环. 吉林黑土区不同施肥处理对农田土壤昆虫的影响. 生态学报, 2006, 26(4): 1122–1130. |
| [34] | 李娜, 张雪萍, 张利敏. 三种温带森林大型土壤动物群落结构的时空动态. 生态学报, 2013, 33(19): 6236–6245. |
| [35] | 刘继亮, 殷秀琴, 邱丽丽. 左家自然保护区大型土壤动物与土壤因子关系研究. 土壤学报, 2008, 45(1): 130–136. |