 2017, Vol. 37
2017, Vol. 37文章信息
- 解欢欢, 马文瑛, 赵传燕, 高云飞, 王清涛, 葛红元, 林梅, 黄晖云
- XIE Huanhuan, MA Wenying, ZHAO Chuanyan, GAO Yunfei, WANG Qingtao, GE Hongyuan, LIN Mei, HUANG Huiyun.
- 苔藓和凋落物对祁连山青海云杉林土壤呼吸的影响
- Effects of litter and moss on soil respiration in a spruce forest in the Qilian Mountains, Qinghai
- 生态学报. 2017, 37(5): 1379-1390
- Acta Ecologica Sinica. 2017, 37(5): 1379-1390
- http://dx.doi.org/10.5846/stxb201510132067
-
文章历史
- 收稿日期: 2015-10-13
- 网络出版日期: 2016-07-13





2. 兰州大学草地农业科学技术学院, 草地农业生态系统国家重点实验室, 兰州 730000;
3. 甘肃省张掖市林业局寺大隆林场, 张掖 734000
2. College of Pastoral Agriculture Science and Technology, State Key Laboratory of Grassland and Agro-Ecosystem, Lanzhou University, Lanzhou 730000, China;
3. Zhangye forest bureau, Sidalong station, Zhangye 734000, China
土壤是全球陆地系统中最大的碳库载体, 其碳储存量1300—2000PgC, 是全球植被碳库的2—3倍, 是大气碳库的2倍多[1]。据研究, 每年因土壤呼吸向大气中排放的碳约占全球总排放量的5%—25%[2], 超过全球陆地生态系统净初级生产力, 也超过由化石燃料等燃烧向大气中排放的CO2量, 因此土壤呼吸的微小变化就可能引起大气CO2浓度的较大改变, 进而影响气候变化。
苔藓在森林生态系统中发挥着重要的生态服务功能, 如储存碳、涵养水源等[3], 目前对苔藓的研究主要集中在生物指示监测、养分循环 (主要是N、P K、Ca、Mg等) 涵养水源和水土保持等方面[3-5], 而有关苔藓影响全球碳循环方面的研究较少。苔藓生物虽然个体小, 但其具有庞大的净初级生产力和生产量, 有研究发现北方针叶林下羽藓的净初级生产力 (NPP) 能达到20—80 g cm-2 a-1 [6], 亚北极地区夏初和夏末苔藓植物的NPP分别为0.37 g m-2 d-1和0.46 g m-2 d-1 [7]。与此形成鲜明对比的是, 苔藓凋落物的分解速率缓慢[3], 积累了巨大的生物量, 因此苔藓在碳汇方面发挥着重要作用。Goulden等[8]的研究表明, 苔藓光合所固定的碳占森林吸收CO2量的10%—50%, 而呼吸占森林生态系统呼吸的50%—90%。苔藓在全球碳循环中究竟是碳汇还是碳源?苔藓呼吸对森林生态系统呼吸的贡献率有多少?苔藓是通过改变环境因子来改变土壤呼吸吗?对这些问题的回答有助于我们对森林生态系统土壤呼吸的机理理解。祁连山区的青海云杉林 (Picea crassifolia) 占水源涵养林总面积的21.7%, 占乔木林总面积66.4%[9], 林下苔藓覆盖面积达99.3%, 生物量达2418 g/m2, 占总生物量的9.95%[10]。因此研究青海云杉林下的苔藓对土壤呼吸影响具有重要理论意义。
凋落物作为森林地表的另一种覆盖物, 是生态系统物质循环和能量转换的主要途径。凋落物数量、组成及其分解速率在很大程度上影响着土壤有机质的形成和对植物养分的供应, 凋落物覆盖还可通过影响土壤水热因子, 土壤微环境以及微生物种类和数量, 直接或间接地影响土壤CO2通量[11-12], 因此凋落物是影响生态系统碳汇功能的重要因素。目前国内外不乏对凋落物呼吸的研究[13-16], 但却鲜有青海云杉林下凋落物对土壤呼吸速率影响的报道。青海云杉平均年凋落量为2.166 t/hm2[17]。本文针对青海云杉林下苔藓和凋落物对土壤呼吸的影响进行研究, 并分析不同覆盖条件下土壤呼吸的日变化和月变化动态以及不同覆盖物对微环境因子的影响, 以期理解青海云杉林生态系统土壤呼吸及其对森林生态系统碳循环的贡献。
1 材料和方法 1.1 研究区概况试验区位于祁连山黑河上游的天涝池流域 (38°20′—38°30 ′N, 99°44′—99°59 ′E)。研究区域面积12.8 km2, 海拔2600—4450 m, 森林覆盖率65%。气候特点是冬季长而寒冷干燥, 夏季短而温凉湿润, 为典型的大陆性高寒半湿润山地气候[18]。年均气温-0.6—2.0℃, 年降水量400—500 mm, 降水主要集中在6—9月份, 占全年降水量的89.2%, 潜在蒸发量1051.7 mm, 年日照时数1892.6 h, 年平均相对湿度为60%。土壤水分有两大主要输入:在5月和6月初为融化的雪水, 在6月到9月为降雨。青海云杉林主要分布在流域阴坡。本研究区的青海云杉林主要为藓类青海云杉林[19]。
本研究选取藓类青海云杉林 (海拔3083 m, 99°56′E, 38°26′N) 作为试验样地, 群落结构简单, 上层为青海云杉, 下层为苔藓, 其中山羽藓为优势苔藓物种。藓类云杉林样地具体信息为:土壤为粉沙壤土, 云杉密度600株/hm2, 平均树高10.83 m, 平均胸径51.96 cm2, 苔藓盖度90%, 厚度为6.5 cm, 现存量为1935 g/m2(干重), 凋落物以云杉小枝和叶为主, 厚度为0.83 cm, 现存量为369 g/m2(干重)。
1.2 研究方法于2012—2014年生长季 (5—9月), 在样地内随机布设9个PVC环 (内径20 cm, 高13 cm), 其中3个为苔藓处理, 即保持苔藓的原始状态, 所测结果为苔藓覆盖土壤呼吸;3个为凋落物处理, 即保留地表的云杉凋落物, 所测结果为凋落物覆盖土壤呼吸;3个为裸土处理, 即将PVC管里的苔藓和凋落物自土壤表层清除, 所测结果为裸土土壤呼吸。为避免PVC环的安装对土壤造成太大的扰动, 在测定前一周将PVC环埋入土壤, 露出地面2—3 cm左右, 整个观测过程保持PVC环位置不变。使用LI8100土壤碳通量自动测量系统 (LI-COR, Lincoln, NE, USA) 对土壤呼吸速率 (Rs) 进行测定, 同时可以获得地表温度 (Ta) 和地表相对湿度数据 (RH), 利用LI8100系统自带的温度探头和湿度探头测定10 cm处土壤温度 (Ts) 和5cm处土壤体积含水量 (VWC)。仪器每1 s记录一次数据, 每2 min求取1个平均值。白天测定时间为7:00到19:00, 在2012年7月和8月各选择一天进行裸土土壤呼吸的昼夜测定, 时间为7:00到第二天7:00, 尽量选取晴朗天气, 测定时间步长均为2 h。
1.3 数据处理与分析 1.3.1 数据处理将平行样求取平均, 获得不同处理条件下的土壤呼吸速率 (Rs) 和环境要素 (Ts, Ta, RH), 用于分析呼吸速率及环境因子的日变化动态。将小时数据进行平均, 获得日平均值, 将该月内的日平均值再次平均获得月平均值, 用于分析呼吸速率及环境因子的月变化动态。
1.3.2 呼吸速率与温度之间的关系
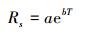
|
(1) |
式中, Rs为呼吸速率 (μmol m-2 s-1), T为土壤温度或地表温度 (℃), a为温度0℃时的呼吸速率 (μmol m-2 s-1), b为温度敏感系数, a和b可以通过观测的温度与土壤呼吸速率获得。Q10代表呼吸速率对温度的敏感性, 是指温度每升高10℃时呼吸速率变化的倍数, 是定量描述土壤呼吸与温度关系的指数, 可通过下式确定:

|
(2) |
采用如下公式计算苔藓和凋落物对土壤呼吸的贡献[22]:

|
(5) |
式中, LCD代表地表凋落物或苔藓的贡献率;CKD为测定的苔藓组或凋落物组的Rs平均值, NLD为裸土组的Rs平均值。
1.3.4 统计分析采用One-Way ANOVA来分析裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤之间的呼吸速率以及环境因子之间的显著性差异 (SPSS 18, Chicago, IL, USA), 并分析呼吸速率和环境因子的季节性差异。采用Sigmaplot 12.5软件进行作图。
2 结果和分析 2.1 土壤呼吸和环境因子的时间变化动态 2.1.1 裸土土壤呼吸速率及环境因子的昼夜动态变化土壤呼吸和环境因子在日变化尺度上具有显著差异性 (P < 0.03)。土壤呼吸日变化动态表现为单峰曲线 (图 1)。Rs从7:00逐渐增加, 15:00达到最大值 ((3.34±0.06) μmol m-2 s-1), 随后逐渐下降, 直到第二天7:00。地表温度 (Ta) 与土壤温度 (Ts) 的变化趋势与Rs一致, 并都在15:00达到最大值。地表相对湿度 (RH) 从观测开始随着时间推移逐渐下降, 15:00下降到最低值 (52.25%), 然后逐渐上升, 直到第二天7:00。

|
| 图 1 裸土土壤呼吸和环境因子的昼夜变化 Fig. 1 Temporal variation of soil respiration and environmental factors in bare soil Rs为裸土土壤呼吸速率soil respiration in bare soil, , Ts为10 cm土壤温度soil temperature at 10 cm, Ta为地表温度soil surface temperature, RH为地表相对湿度soil surface relative humidity |
苔藓覆盖土壤、凋落物覆盖土壤的呼吸和环境因子的日变化动态与裸土土壤相似, 均表现为单峰曲线, 但由于覆盖物的不同, 各个处理组的呼吸速率达到最大值的时间不同。以2012年6月22日和6月24日的观测数据为例 (图 2), 裸土土壤和凋落物覆盖土壤的Rs在13:00达到最大值, 而苔藓覆盖土壤的Rs在15:00达到最大值。Ta和Ts分别在13:00和15:00达到最大值, Ta和Ts之间存在滞后。

|
| 图 2 2012年6月土壤呼吸及环境因子的日间变化 Fig. 2 Diurnal variation of soil respiration rate and environmental factors in June 2012 Rs, Ts, Ta和RH的含义同图 1, 其中“裸”、“苔”和“凋”分别代表裸土处理, 苔藓覆盖处理和凋落物覆盖处理The meaning of Rs, Ts, Taand RH have been showed in fig 1, In addition, “裸”, “苔” and “凋” represent bare soil treatment, moss treatment and litter treatment, respectively |
在生长季期间, 呼吸速率和土壤温度表现出显著的月变化 (P < 0.02)。图 3显示:3组处理的呼吸速率均随着温度的上升而上升。将所有数据求取月平均值得到:3组处理的Rs最大值均出现在8月, 最小值均出现在5月, 且月均呼吸值大小表现为裸土土壤呼吸 ((2.24±0.22) μmol m-2 s-1) < 凋落物覆盖土壤呼吸 ((3.06±0.41)μmol m-2 s-1) < 苔藓覆盖土壤呼吸 ((3.48±0.53)μmol m-2 s-1);裸土土壤呼吸、苔藓覆盖土壤呼吸和凋落物覆盖土壤呼吸月变动范围分别在1.67—2.85、1.73—4.63 μmol m-2 s-1和1.77—4.15 μmol m-2 s-1之间。

|
| 图 3 三组处理的Rs、Ta、Ts和RH的月变化 Fig. 3 Monthly variation of Rs, Ta, Ts, and RHin three treatments |
裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤间的呼吸速率和环境因子的差异性如图 4显示:裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤两两之间的Rs均达到显著差异 (P=0.001)。3组处理之间的RH没有显著性差异, 但裸土土壤和苔藓覆盖土壤之间的RH接近显著 (P=0.059)。3组处理之间的Ta、Ts和土壤水分 (VWC) 均没有显著差异。裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤的年均Rs分别为 (2.28±0.31), (3.88±0.26) μmol m-2 s-1和 (3.31±0.19) μmol m-2 s-1, 年均Ta分别为 (11.06±0.68)℃, (11.20±0.76)℃和 (11.15±0.74)℃, 年均Ts分别为 (6.64±0.35)℃, (7.18±0.51)℃和 (7.32±0.52)℃, 年均VWC分别为 (38.64±3.38)%, (39.25±3.07)%和 (32.00±2.98)%。苔藓和凋落物组Rs分别高于裸土土壤组Rs的70.18%和45.18%;苔藓和凋落物组Ta分别高于裸土组Ta的1.27%和0.81%;苔藓和凋落物组Ts分别高于裸土组Ts的8.13%和10.24%。

|
| 图 4 3组处理的Rs、Ta、Ts、RH和VWC在3a之间的显著性差异 Fig. 4 Significant difference of Rs, Ta, Ts, and RH among three treatments in three years |
苔藓和凋落物对裸土土壤呼吸的贡献具有明显的月变化差异。图 5显示:苔藓和凋落物对土壤呼吸的贡献率在5月份最低, 分别只有3.93%和5.63%, 但在6月份急剧上升, 苔藓贡献率在9月份达到最大值67.96%, 凋落物贡献率在8月份达到最大值63.96%。将所有数据求取月贡献率, 得到苔藓对裸土土壤呼吸的月贡献率在3.93%—39.01%之间, 凋落物对裸土土壤呼吸的月贡献率在5.63%—30.92%之间;苔藓和凋落物对裸土土壤呼吸的月均贡献率分别为29.33%和24.06%。

|
| 图 5 苔藓和凋落物对土壤呼吸的贡献率 Fig. 5 Contribution rates of moss and litter to total soil respiration rate |
在裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤中, 呼吸速率与温度的相关性均达到极显著, 且呼吸速率与低温的拟合效果要高于高温 (图 6)。地表温度可以分别解释裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤呼吸的68.80%, 52.91%, 60.93%;土壤温度可以分别解释裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤呼吸的79.56%, 91.11%, 84.66%。相比于地表温度, 呼吸速率对土壤温度的响应要高于地表温度。在3组处理中, Q10值的排序为:苔藓覆盖土壤 (Q10=5.47)>凋落物覆盖土壤 (Q10=3.67)>裸土土壤 (Q10=2.23)。

|
| 图 6 呼吸速率与地表温度和土壤温度之间的关系 Fig. 6 The correlation relationship between soil respiration rate with surface temperature or soil temperature |
呼吸速率与土壤含水量在裸土土壤中呈二次多项式关系 (图 7a), 当VWC < 34%时, 裸土土壤呼吸速率随土壤含水量的上升而增加, 当VWC>34%时, 裸土土壤呼吸速率反而随着土壤含水量的上升而下降;在苔藓覆盖土壤和凋落物覆盖土壤中, 呼吸速率与土壤含水量均呈线性负相关关系 (图 7b和图 7c)。土壤含水量可以解释呼吸速率的43%—65%。

|
| 图 7 呼吸速率与土壤含水量之间的关系 Fig. 7 The relationship between soil respiration and soil water content |
裸土土壤、苔藓覆盖土壤和凋落物覆盖土壤的呼吸速率日变化均表现为单峰曲线, 与土壤温度和地表温度的变化趋势基本一致, 说明呼吸速率的日变化主要受温度影响。3组处理的Rs达到最大值的时间不尽相同, 从整个生长季来看, 裸土土壤和凋落物土壤Rs最大值出现在13:00, 苔藓覆盖土壤的最大Rs通常滞后于裸土土壤2 h左右, 可能的原因是苔藓作为屏障阻碍了热量对土壤的直接作用[23]。从月变化看, 本研究得出:裸土处理Rs变动范围在1.67—-2.85 μmol m-2 s-1之间, 马文瑛[24]的观测结果为1.25—3.19 μmol m-2 s-1, 郑祥霖[25]的观测结果为1.41—4.09 μmol m-2 s-1, 彭家中[26]的观测结果为0.50—3.88 μmol m-2 s-1。虽然植被类型相同, 但前人的研究结果与本研究结果有所差异, 这可能是土壤的空间异质性所导致, 上层的林冠结构差异也可能是原因之一。从月变化看, 本研究中, 苔藓覆盖处理下Rs变动范围在1.73—4.63 μmol m-2 s-1之间, 郑祥霖[25]的观测结果为0.74—4.03 μmol m-2 s-1, Botting[27]的观测结果为0.4—1.4 μmol m-2 s-1, 均小于本研究的观测结果, 这可能是苔藓的厚度、种类或海拔等因子的不同所造成的。从不同的群落上看, 本研究中, 凋落物组Rs变动范围在1.77—4.15 μmol m-2 s-1之间, 熊莉[28]对粗枝云杉的观测结果为0.40—5.15 μmol m-2 s-1, 张超[29]对马尾松的观测结果为0.69—2.83 μmol m-2 s-1, 均小于本研究结果, 这可能与不同树种产生的凋落物量和凋落物组成有关, 凋落物越少, 凋落物分解率越低, 土壤呼吸速率就相对较低。
3.2 呼吸速率与温度和土壤含水量的关系研究者普遍认为, 温度和水分是影响土壤呼吸的主要因素[30], 尤其在干旱和半干旱地区, 土壤温度和水分对土壤呼吸的影响显得更为重要[31], 所以这两大因素常被用来解释土壤呼吸速率的大部分变化[32]。大多数研究表明指数模型能够较好地模拟土壤呼吸速率与温度之间的关系[26, 28, 33-34]。本研究中指数模型同样也获得了较高的相关系数, 且呼吸速率与土壤温度的相关性要大于地表温度。苔藓和凋落物覆盖分别增加了土壤温度的8.13%和10.24%, 这可能是其增加土壤呼吸速率的原因之一。温度越高, 苔藓枯落物和云杉凋落物分解速率越快, 土壤微生物代谢更加活跃, 固对呼吸速率有所提高。王光军[35]等研究指出凋落物去除降低了杉木人工林土壤温度, 与本文研究结果一致, 而熊莉[28]研究得出凋落物去除使土壤温度升高了1.2%。
本研究中呼吸速率与土壤含水量表现出极显著相关性, 在裸土土壤呼吸中, 呼吸速率与土壤含水量呈高斯函数关系, 当VWC < 34%时, 呼吸速率随含水量的上升而增加, 当VWC > 34%时, 呼吸速率随含水量的上升而下降;在苔藓覆盖土壤和凋落物覆盖土壤中, 呼吸速率与土壤含水量均呈线性负相关关系。由此看来, 呼吸速率与土壤水分的关系受地表覆盖物的影响。大多数研究表明:土壤湿度对土壤呼吸有双向调节作用, 最适模型为二次曲线模型[36-38], 常宗强在祁连山高山草甸[39]和亚高山灌丛[40]研究中分别得到呼吸速率与土壤含水量的临界值是30.2%和28%。Robert[41]对黑云杉林下的苔藓呼吸结果显示:无论是裸土处理还是苔藓处理, 呼吸速率和土壤含水量均呈弱的负相关性。王春燕[42]认为土壤呼吸速率和土壤水分可能没有直接关系, Bolstad[43]对5种森林类型土壤呼吸的研究结果是土壤呼吸随立地水分含量的增加而减小, 熊莉认为土壤水分的对数值均与土壤呼吸速率呈显著线性正相关。呼吸速率与土壤含水量的关系比较复杂, 其受地上植被类型、土壤含水量、降雨等因素的影响, 需要作进一步研究。
3.3 苔藓和凋落物对土壤呼吸的影响本研究中苔藓提高了裸土土壤呼吸速率的70.18%, 对土壤呼吸的贡献为29.33%。Ekaterina等人[44]也表明苔藓覆盖土壤较裸土有较高的呼吸速率。Robert等人[41]对黑云杉林下的苔藓呼吸的研究表明, 在春季和秋季, 苔藓光合作用所固定的CO2和呼吸释放的CO2相抵消, 在夏季, 苔藓的呼吸作用高于光合作用, 表现为碳损失, 苔藓覆盖土壤的呼吸速率最高可达到7 μmol m-2 s-1。Goulden等人[8]对黑云杉林的地表苔藓呼吸的研究得出, 羽状苔藓和泥炭藓在正午的光合速率分别为0.5—1.0 μmol m-2 s-1和0.5—2.5 μmol m-2 s-1, 在夜间羽状苔藓覆盖土壤和泥炭藓覆盖土壤的呼吸速率分别为1—2.5 μmol m-2 s-1和0—1.5 μmol m-2 s-1。O′Neill等人[45]对森林火灾后的苔藓建植土壤的观测表明, 苔藓在最适水分条件下对土壤呼吸的贡献率在10%—55%之间。综述文献, 苔藓覆盖土壤呼吸速率高于裸土土壤的原因主要有5点:1) 苔藓生物量巨大, 本身呼吸量大;2) 苔藓的长期覆盖使得土壤表层有大量的腐殖质积累;3) 苔藓上寄居着能够分解有机碳的微生物[46];4) 苔藓可以通过聚集有机碳、生产氨盐基提高土壤养分含量[41];5) 苔藓覆盖可以提高土壤水分含量[47]。但郑祥霖等人[48]发现苔藓会降低土壤呼吸速率的8.83%, 他们认为苔藓的覆盖阻碍了土壤与大气间的气体交换。O′Neill[49]报道苔藓会降低土壤温度, 抑制有机质分解, 从而降低土壤呼吸速率。苔藓也会通过改变土壤颗粒、气孔导度、pH[50]、渗滤液[49]来影响土壤呼吸。目前有关苔藓对全球碳循环贡献的研究较少, 苔藓对土壤呼吸的正负作用仍有待于长期观测研究。
本研究得出凋落物提高了裸土土壤呼吸速率的45.18%, 对土壤呼吸的贡献为24.06%。凋落物是土壤呼吸的重要介质[51]。凋落物的存在会抑制土壤降温, 提高土壤湿度, 从而增加土壤呼吸[34], 凋落物也会通过影响土壤的淋溶而影响土壤呼吸[12], 与裸土处理相比, 凋落物的输入会增加矿质土壤微生物量[52-53], 并且促进微生物呼吸[54]。Vasconcelos[55]在巴西的热带再生林移除所有的叶片和枝条凋落物得到凋落物对土壤呼吸的贡献有28%。Sayer[56]在巴拿马热带雨林的研究为凋落物对土壤呼吸的贡献率高达20%。
4 结论(1) 土壤表层的覆盖物情况会影响最大呼吸速率的出现时间。凋落物覆盖土壤和裸土土壤的最大呼吸速率出现时间一致, 在13:00或15:00达到最大值, 而苔藓覆盖土壤滞后于这两组2 h, 在15:00和17:00达到最大值。苔藓覆盖物将土壤呼吸的峰值推后。
(2) 覆盖物本身的呼吸对总呼吸的贡献会影响土壤呼吸, 本研究中苔藓和凋落物对裸土土壤呼吸速率的贡献率分别为29.33%和24.06%;覆盖物也会通过改变土壤的微环境间接影响土壤呼吸。但本研究中前者的作用比较明显。因为3种处理间的呼吸速率达到显著性差异, 地表相对湿度、地表温度、土壤温度和土壤含水量均没有显著差异。
(3) 覆盖物能提高土壤呼吸对温度的敏感性, 苔藓和凋落物处理均提高了Q10值, 3组处理Q10大小为:苔藓覆盖土壤 (Q10=5.47) > 凋落物覆盖土壤 (Q10=3.67) > 裸土土壤 (Q10=2.23)。
(4)3组处理的呼吸速率均与温度呈显著指数相关, 且与土壤温度的相关性高于地表温度0.06—0.23;土壤水分对土壤呼吸的影响有正负作用, 无覆盖条件下, 正负作用存在阈值, 本研究得出VWC=34%, 有覆盖物条件下, 土壤呼吸速率与土壤含水量均呈线性负相关关系。
| [1] | 陈泮勤, 黄耀, 于贵瑞. 地球系统碳循环. 北京: 科学出版社, 2004. |
| [2] | Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemistry Cycles, 1995, 9(1): 23–36. DOI:10.1029/94GB02723 |
| [3] | 叶吉, 郝占庆, 于德永, 闫海冰, 封德全. 苔藓植物生态功能的研究进展. 应用生态学报, 2004, 15(10): 1939–1942. |
| [4] | 吴玉环, 黄国宏, 高谦, 曹同. 苔藓植物对环境变化的响应及适应性研究进展. 应用生态学报, 2001, 12(6): 943–946. |
| [5] | 郝占庆, 叶吉, 姜萍, 蔺菲. 长白山暗针叶林苔藓植物在养分循环中的作用. 应用生态学报, 2005, 16(12): 2263–2266. |
| [6] | Bisbee K E, Gower S T, Norman J M, Nordheim E V. Environmental controls on ground cover species composition and productivity in a boreal black spruce forest. Oecologia, 2001, 129(2): 261–270. DOI:10.1007/s004420100719 |
| [7] | Campioli M, Samson R, Michelsen A, Jonasson S, Baxter R, Lemeur R. Nonvascular contribution to ecosystem NPP in a subarctic heath during early and late growing season. Plant Ecology, 2009, 202(1): 41–53. DOI:10.1007/s11258-008-9527-6 |
| [8] | Goulden M L, Crill P M. Automated measurements of CO2 exchange at the moss surface of a black spruce forest. Tree Physiology, 1997, 17(8/9): 537–542. |
| [9] | 王金叶, 张学龙, 张虎, 王艺林, 金铭. 祁连山水源涵养林组成结构及生长状况. 西北林学院学报, 2001, 16(S): 4–7. |
| [10] | 王金叶, 车克均, 傅辉恩, 常学向, 宋采福, 贺红元. 祁连山水源涵养林生物量的研究. 福建林学院学报, 1998, 18(4): 319–323. |
| [11] | Wan S Q, Norby R J, Ledford J, Weltzin J F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Global Change Biology, 2007, 13(11): 2411–2424. DOI:10.1111/gcb.2007.13.issue-11 |
| [12] | Sulzman E W, Brant J B, Bowden R D, Lajtha K. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry, 2005, 73(1): 231–256. DOI:10.1007/s10533-004-7314-6 |
| [13] | Zhou X H, Talley M, Luo Y Q. Biomass, litter, and soil respiration along a precipitation gradient in Southern Great Plains, USA. Ecosystems, 2009, 12(8): 1369–1380. DOI:10.1007/s10021-009-9296-7 |
| [14] | Wu C S, Zhang Y P, Xu X L, Sha L Q, You G Y, Liu Y H, Xie Y N. Influence of interactions between litter decomposition and rhizosphere activity on soil respiration and on the temperature sensitivity in a subtropical montane forest in SW China. Plant and Soil, 2014, 381(1/2): 215–224. |
| [15] | Wang Y K, Fang S Z, Chang S X, Tian Y. Non-additive effects of litter-mixing on soil carbon dioxide efflux from poplar-based agroforestry systems in the warm temperate region of China. Agroforestry Systems, 2014, 88(2): 193–203. DOI:10.1007/s10457-013-9665-2 |
| [16] | Wang Y D, Wang H M, Ma Z Q, Wen X F, Li Q K, Liu Y F, Sun X M, Yu G R. Contribution of aboveground litter decomposition to soil respiration in a subtropical coniferous plantation in Southern China. Asia-Pacific Journal of Atmospheric Sciences, 2009, 45(2): 137–147. |
| [17] | 王金叶, 车克钧, 蒋志荣. 祁连山青海云杉林碳平衡研究. 西北林学院学报, 2000, 15(1): 9–14. |
| [18] | 刘兴明.祁连山青海云杉林潜在分布及其生物量碳空间特征研究[D].兰州:甘肃农业大学, 2012. |
| [19] | 刘兴聪. 青海云杉. 兰州: 兰州大学出版社, 1992. |
| [20] | Buchmann N. Biotic and abiotic factors controlling soil respiration rates in Picea abies stands. Soil Biology and Biochemistry, 2000, 32(11-12): 1625–1635. DOI:10.1016/S0038-0717(00)00077-8 |
| [21] | Sánchez M L, Ozores M I, López M J, Colle R, De Torre B, García M A, Pérez I. Soil CO2 fluxes beneath barley on the central Spanish plateau. Agricultural and Forest Meteorology, 2003, 118(1/2): 85–95. |
| [22] | 赵昕, 张万军, 沈会涛, 艾治频, 廉诗启, 刘长柏. 针阔树种人工林地表凋落物对土壤呼吸的贡献. 中国生态农业学报, 2014, 22(11): 1318–1325. |
| [23] | Soudzilovskaia N A, van Bodegom P M, Cornelissen J H C. Dominant bryophyte control over high-latitude soil temperature fluctuations predicted by heat transfer traits, field moisture regime and laws of thermal insulation. Functional Ecology, 2013, 27(6): 1442–1454. DOI:10.1111/1365-2435.12127 |
| [24] | 马文瑛, 赵传燕, 彭守璋, 高云飞, 袁亚鹏, 李文娟. 黑河天涝池五种植被类型土壤呼吸速率动态特征及其影响因子. 生态学报, 2015, 35(17): 5654–5665. |
| [25] | 郑祥霖.祁连山中部青海云杉林碳循环研究[D].兰州:兰州大学, 2012. |
| [26] | 彭家中, 常宗强, 冯起. 温度和土壤水分对祁连山青海云杉林土壤呼吸的影响. 干旱区资源与环境, 2008, 22(3): 165–169. |
| [27] | Botting R S, Fredeen A L. Net ecosystem CO2 exchange for moss and lichen dominated forest floors of old-growth sub-boreal spruce forests in central British Columbia, Canada. Forest Ecology and Management, 2006, 235(1/3): 240–251. |
| [28] | 熊莉, 徐振锋, 杨万勤, 殷睿, 唐仕姗, 王滨, 徐李亚, 常晨晖. 川西亚高山粗枝云杉人工林地上凋落物对土壤呼吸的贡献. 生态学报, 2015, 35(4): 4678–4686. |
| [29] | 张超.凋落物对亚热带4种森林土壤呼吸的影响[D].长沙:中南林业科技大学, 2013. |
| [30] | Burton A J, Pregitzer K S. Field measurements of root respiration indicate little to no seasonal temperature acclimation for sugar maple and red pine. Tree Physiology, 2003, 23(4): 273–280. DOI:10.1093/treephys/23.4.273 |
| [31] | 王新源, 李玉霖, 赵学勇, 毛伟, 崔夺, 曲浩, 连杰, 罗永清. 干旱半干旱区不同环境因素对土壤呼吸影响研究进展. 生态学报, 2012, 32(15): 4890–4901. |
| [32] | Lloyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315–323. DOI:10.2307/2389824 |
| [33] | 刘颖, 韩士杰, 胡艳玲, 戴冠华. 土壤温度和湿度对长白松林土壤呼吸速率的影响. 应用生态学报, 2005, 16(9): 1581–1585. |
| [34] | 姜艳, 王兵, 汪玉如, 杨清培. 亚热带林分土壤呼吸及其与土壤温湿度关系的模型模拟. 应用生态学报, 2010, 21(7): 1641–1648. |
| [35] | 王光军, 田大伦, 闫文德, 朱凡, 项文华, 梁小翠. 改变凋落物输入对杉木人工林土壤呼吸的短期影响. 植物生态学报, 2009, 33(4): 739–747. |
| [36] | 张红星, 王效科, 冯宗炜, 宋文质, 刘文兆, 李双江, 朱元骏, 庞军柱, 欧阳志云. 干湿交替格局下黄土高原小麦田土壤呼吸的温湿度模型. 生态学报, 2009, 29(6): 3028–3035. |
| [37] | 王君, 沙丽清, 李检舟, 冯志立. 云南香格里拉地区亚高山草甸不同放牧管理方式下的碳排放. 生态学报, 2008, 28(8): 3574–3583. |
| [38] | Liu Y C, Liu S R, Wang J X, Zhu X L, Zhang Y D, Liu X J. Variation in soil respiration under the tree canopy in a temperate mixed forest, central China, under different soil water conditions. Ecological Research, 2014, 29(2): 133–142. DOI:10.1007/s11284-013-1110-5 |
| [39] | 常宗强, 冯起, 吴雨霞, 苏永红. 祁连山亚高山灌丛林土壤呼吸速率的时空变化及其影响分析. 冰川冻土, 2005, 27(5): 666–672. |
| [40] | 常宗强, 冯起, 司建华, 苏永红, 席海洋, 郭瑞. 祁连山高山草甸土壤CO2通量的时空变化及其影响分析. 环境科学, 2007, 28(10): 2389–2395. |
| [41] | Swanson R V, Flanagan L B. Environmental regulation of carbon dioxide exchange at the forest floor in a boreal black spruce ecosystem. Agricultural and Forest Meteorology, 2001, 108(3): 165–181. DOI:10.1016/S0168-1923(01)00243-X |
| [42] | 王春燕, 杨秋波, 袁坤, 李光明, 杨礼富, 王真辉. 橡胶林土壤呼吸速率及其与土壤温湿度的关系. 土壤学报, 2013, 50(5): 974–982. |
| [43] | Martin J G, Bolstad P V. Annual soil respiration in broadleaf forests of northern Wisconsin:influence of moisture and site biological, chemical, and physical characteristics. Biogeochemistry, 2005, 73(1): 149–182. DOI:10.1007/s10533-004-5166-8 |
| [44] | Sedia E G, Ehrenfeld J G. Differential effects of lichens, mosses and grasses on respiration and nitrogen mineralization in soils of the New Jersey Pinelands. Oecologia, 2005, 144(1): 137–147. DOI:10.1007/s00442-005-0037-0 |
| [45] | O'Neill K P, Richter D D, Kasischke E S. Succession-driven changes in soil respiration following fire in black spruce stands of interior Alaska. Biogeochemistry, 2006, 80(1): 1–20. DOI:10.1007/s10533-005-5964-7 |
| [46] | Thormann M N, Currah R S, Bayley S E. The relative ability of fungi from Sphagnum fuscum to decompose selected carbon substrates. Canadian Journal of Microbiology, 2002, 48(3): 204–211. DOI:10.1139/w02-010 |
| [47] | Eckstein R L. Nitrogen retention by Hylocomium splendens in a subarctic birch woodland. Journal of Ecology, 2000, 88(3): 506–515. DOI:10.1046/j.1365-2745.2000.00480.x |
| [48] | Zheng X L, Zhao C Y, Peng S Z, Jian S Q, Liang B, Wang X P, Yang S F, Wang C, Peng H H, Wang Y. Soil CO2 efflux along an elevation gradient in Qinghai Spruce forests in the upper reaches of the Heihe River, northwest China. Environmental Earth Sciences, 2014, 71(5): 2065–2076. DOI:10.1007/s12665-013-2608-4 |
| [49] | O'Neill K. Bryophytes in the Global Carbon Budget. Cambridge: Cambridge University Press, 2000. |
| [50] | Turetsky M R. The role of bryophytes in carbon and nitrogen cycling. The Bryologist, 2003, 106(3): 395–409. DOI:10.1639/05 |
| [51] | DeForest J L, Chen J Q, McNulty S G. Leaf litter is an important mediator of soil respiration in an oak-dominated forest. International Journal of Biometeorology, 2009, 53(2): 127–134. DOI:10.1007/s00484-008-0195-y |
| [52] | Feng W T, Zou X M, Schaefer D. Above-and belowground carbon inputs affect seasonal variations of soil microbial biomass in a subtropical monsoon forest of southwest China. Soil Biology and Biochemistry, 2009, 41(5): 978–983. DOI:10.1016/j.soilbio.2008.10.002 |
| [53] | Li Y Q, Xu M, Sun O J, Cui W C. Effects of root and litter exclusion on soil CO2 efflux and microbial biomass in wet tropical forests. Soil Biology and Biochemistry, 2004, 36(12): 2111–2114. DOI:10.1016/j.soilbio.2004.06.003 |
| [54] | Lajtha K, Crow S E, Yano Y, Kaushal S S, Sulzman E, Sollins P, Spears J D H. Detrital controls on soil solution N and dissolved organic matter in soils:a field experiment. Biogeochemistry, 2005, 76(2): 261–281. DOI:10.1007/s10533-005-5071-9 |
| [55] | Vasconcelos S S, Zarin D J, Capanu M, Littell R, Davidson E A, Ishida F Y, Santos E B, Araújo M M, Aragão D V, Rangel-Vasconcelos L G T, de Assis Oliveira F, McDowell W H, de Carvalho C J R. Moisture and substrate availability constrain soil trace gas fluxes in an eastern Amazonian regrowth forest. Global Biogeochemical Cycles, 2004, 18(2). |
| [56] | Sayer E J, Powers J S, Tanner E V J. Increased litterfall in tropical forests boosts the transfer of soil CO2 to the atmosphere. PLoS One, 2007, 2(12): e1299. DOI:10.1371/journal.pone.0001299 |