 2017, Vol. 37
2017, Vol. 37文章信息
- 康勇, 熊梦辉, 黄瑾, 龙文兴, 杨小波, 臧润国, 王茜茜, 林灯
- KANG Yong, XIONG Menghui, HUANG Jin, LONG Wenxing, YANG Xiaobo, ZANG Runguo, WANG Xixi, LIN Deng.
- 海南岛霸王岭热带云雾林木本植物功能性状的分异规律
- Variation in woody plant functional traits of the tropical cloud forests in Bawangling, Hainan Island
- 生态学报. 2017, 37(5): 1572-1582
- Acta Ecologica Sinica. 2017, 37(5): 1572-1582
- http://dx.doi.org/10.5846/stxb201510082023
-
文章历史
- 收稿日期: 2015-10-08
- 网络出版日期: 2016-07-13





2. 海南大学园艺园林学院, 海口 570228;
3. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091;
4. 海南大学环境与植物保护学院, 海口 570228
2. College of horticulture and landscape architecture, Hainan University, Haikou 570228, China;
3. Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Key Laboratory of Forest Ecology and Environment of State Forestry Administration, Beijing 100091, China;
4. College of Environment and Plant Protection, Hainan University, Haikou 570228, China
植物功能性状通常指影响植物存活、生长、繁殖速率和最终适合度的生物特征[1], 它能反映个体、种群、群落和生态系统水平上的生物之间、生物与环境之间的相互作用, 揭示生物对生态系统功能的影响[2-5], 因而提高了群落生态学的研究水平, 使群落生态学研究从定性描述及复杂模型向定量及简约转化[6]。
植物功能性状与环境密切相关[7], 不同功能性状反映植物的生态策略不同[3], 功能性状的可塑性大小也因物种种类而异[8], 因而环境因子可以作为“筛子”, 影响群落中物种的组配过程[9]。环境筛对功能性状的作用既与空间尺度有关[10], 也与群落组织尺度有关[11-12]。群落组织尺度包括个体、种内、种间及群落水平, 比较这些尺度上功能性状变化, 有助于研究者解释生态格局和生态过程[13]。例如, 多数学者认为种间水平的功能性状变化是功能性状研究的主要途径[14-15], 但近来发现种内功能性状变化也非常重要[16], 甚至认为它比种间功能性状变化对环境更敏感, 可能比种间功能性状对环境筛具有更强的指示作用[17], 因而在群落生态学中扮演着重要角色[18]。还有一些学者认为, 群落水平上研究能将环境因子和功能性状置于同一系统中, 根据功能性状变化解释植物的适应性机制[19-20]。虽然目前很多研究在寻找各种群落组织水平上功能性状分异的潜在来源[12], 但对于各个水平上功能性状分异的相对大小, 仍存在争议[16]。
热带云雾林指潮湿热带地区经常被云雾掩盖的森林[21]。其山风强烈、气温低、云雾多, 土壤含水量常处于饱和状态[21-22]。与典型的低海拔热带林相比, 热带云雾林的树木高度和径级明显较小, 树干常弯曲, 小叶革质, 单叶比例高[23];特有物种和濒危物种丰富[21, 24]。海南岛的热带云雾林主要分布在全岛林区海拔1200 m以上的山顶或山脊[22, 25]。受空气低温和土壤低磷等环境筛影响, 热带云雾林植物的比叶面积和高度都比低海拔森林植物小[8-10], 因而植物功能性状的分异规律可能与热带地区低海拔植被不同。以海南霸王岭热带云雾林为对象, 设置21个20 m×20 m样方, 测定每个样方内所有胸径≥5 cm的个体的叶干重、叶面积、比叶重、叶片叶绿素含量、叶片厚度和木材密度, 以及土壤有机质、全磷、有效磷、全氮和有效氮含量;分析植物功能性状在个体、种内、种间及群落水平的分异大小, 探究不同群落组织尺度上, 土壤因子与功能性状的关系。本文假设如下:(1) 热带云雾林植物功能性状在不同群落组织尺度上分异大小不同, 种内水平分异最大;(2) 热带云雾林土壤因子对群落组织尺度上功能性状分异有显著影响。
1 材料与方法 1.1 研究样地概况研究样地位于霸王岭自然保护区 (18°50′—19°05′ N, 109°05′—109°25′ E), 总面积约50000 hm2, 以山地为主, 海拔范围为100—1654 m。属热带季风气候, 干湿季明显, 5—10月为雨季, 11月至次年4月为旱季。海拔100 m位置年平均气温23.6 ℃, 年降雨量1, 677.1 mm[26]。土壤以砖红壤为代表, 随海拔增加逐渐过渡为山地红壤、山地黄壤和山地草甸土, 沿海拔梯度依次分布有热带低地雨林、热带山地雨林、热带山地常绿林和热带山顶矮林[27]。霸王岭的热带云雾林主要呈岛屿状分布在霸王峰的雅加松顶和松林顶[22]。优势种为海南五针松 (Pinus fenzeliana)、蚊母树 (Distylium racemosum)、碎叶蒲桃 (Syzygium buxifolium)、黄杞 (Engelhardia roxburghiana) 和毛棉杜鹃 (Rhododendron moulmainense) 等。树木平均高度约 (4.79±2.80) m, 密度约9633株/hm2(表 1)。
| 样地 Plot |
地理位置 Location |
样方面积/m2 Plot area |
物种数量 Number of species |
纬度 Latitude |
经度 Longitude |
高度/m Height |
| 样方1 Plot1 | 雅加松顶 (YJSM) | 400 | 45 | 19°04′57.93″ | 109°12′44.14″ | 1329.48 |
| 样方2 Plot2 | 雅加松顶 (YJSM) | 400 | 53 | 19°04′58.78″ | 109°12′44.01″ | 1343.62 |
| 样方3 Plot3 | 雅加松顶 (YJSM) | 400 | 56 | 19°05′00.01″ | 109°12′43.53″ | 1341.76 |
| 样方4 Plot4 | 雅加松顶 (YJSM) | 400 | 63 | 19°05′00.35″ | 109°12′43.69″ | 1351.33 |
| 样方5 Plot5 | 雅加松顶 (YJSM) | 400 | 43 | 19°05′01.76″ | 109°12′44.23″ | 1344.21 |
| 样方6 Plot6 | 雅加松顶 (YJSM) | 400 | 49 | 19°05′03.07″ | 109°12′45.26″ | 1340.57 |
| 样方7 Plot7 | 雅加松顶 (YJSM) | 400 | 45 | 19°05′04.09″ | 109°12′45.39″ | 1331.92 |
| 样方8 Plot8 | 雅加松顶 (YJSM) | 400 | 45 | 19°05′05.15″ | 109°12′45.68″ | 1314.12 |
| 样方9 Plot9 | 雅加松顶 (YJSM) | 400 | 49 | 19°05′06.20″ | 109°12′45.26″ | 1313.24 |
| 样方10 Plot10 | 雅加松顶 (YJSM) | 400 | 47 | 19°05′07.09″ | 109°12′45.61″ | 1318.25 |
| 样方11 Plot11 | 松林顶 (SLDM) | 400 | 57 | 19°05′09.76″ | 109°12′35.76″ | 1368.71 |
| 样方12 Plot12 | 松林顶 (SLDM) | 400 | 49 | 19°05′10.15″ | 109°12′34.93″ | 1389.67 |
| 样方13 Plot13 | 松林顶 (SLDM) | 400 | 47 | 19°05′11.49″ | 109°12′35.69″ | 1395.24 |
| 样方14 Plot14 | 松林顶 (SLDM) | 400 | 58 | 19°05′10.54″ | 109°12′36.26″ | 1380.15 |
| 样方15 Plot15 | 松林顶 (SLDM) | 400 | 56 | 19°05′10.93″ | 109°12′38.91″ | 1372.05 |
| 样方16 Plot16 | 松林顶 (SLDM) | 400 | 39 | 19°05′11.35″ | 109°12′40.40″ | 1373.9 |
| 样方17 Plot17 | 松林顶 (SLDM) | 400 | 40 | 19°05′12.31″ | 109°12′40.76″ | 1369.37 |
| 样方18 Plot18 | 松林顶 (SLDM) | 400 | 46 | 19°05′13.09″ | 109°12′41.07″ | 1366.67 |
| 样方19 Plot19 | 松林顶 (SLDM) | 400 | 43 | 19°05′15.77″ | 109°12′41.77″ | 1343.78 |
| 样方20 Plot20 | 松林顶 (SLDM) | 400 | 45 | 19°05′16.90″ | 109°12′42.43″ | 1338.82 |
| 样方21 Plot21 | 松林顶 (SLDM) | 400 | 44 | 19°05′17.96″ | 109°12′43.02″ | 1318.1 |
2013年7—8月, 用全站仪 (Leica TSP1200+, Heerbrugg, Switzerland) 在霸王岭雅加松顶和松林顶热带云雾林中分别设置10、11个20 m×20 m的样方 (表 1, 图 1), 样方间距离50 m以上, 总面积0.84 hm2。用邻格法将每个400 m2样方划分为16个5 m×5 m小样方, 共得到336个5 m×5 m小样方。调查每个5 m×5 m样方内所有胸径 (DBH)≥5 cm的乔灌木植株, 测定其胸径, 记录每个个体的物种名, 不能现场确定物种, 采集标本带回室内请专家鉴定。共记录2348株植物个体, 67个物种。

|
| 图 1 研究地点的地理位置 Fig. 1 The distribution of the sites |
选取反映植物碳积累、光合作用速率及植物对环境的抗逆能力的功能性状, 如叶干重 (LDW)、木材密度 (WD)、叶面积 (LA)、比叶重 (LMA)、叶片叶绿素含量 (Chl) 和叶片厚度 (LTh)。其中, 植株高度能反映热带云雾林植物对低温及土层薄环境的适应能力及对光、空间等资源的竞争能力。木材密度反映热带云雾林植物的生长速率和抵御病原体和外界的物理性质损害等的能力。叶面积 (LA)、比叶重 (LMA) 以及叶干重 (LDW) 与植物体内光合速率及碳积累有关, 反映植物对水胁迫的反应能力。叶绿素含量与单位质量的叶片氮素浓度密切相关, 所以与光合速率存在着一定的联系[28], 能够提供植物光合能力的信息[29]。叶片厚度反映了热带云雾植物叶片机械强度, 与植物对热带云雾林低温和风力大环境的反应有关[30]。
测定样地内所有的DBH≥5 cm乔灌木个体植株的功能性状 (不含裸子植物)。每个植物个体选取树冠顶端2—3片完全展开的当年成熟的受太阳光照的叶片, 用数显游标卡尺 (SF2000, Guilin, China) 测量叶片边缘与主脉间非叶脉处厚度即为叶片厚度 (mm);用便携式叶绿素仪 (SPAD-502 Plus, Konica Minolta, Japan) 测定叶绿素含量 (Chl, SPAD);用叶面积仪 (LI-COR 3100C Area Meter, LI-COR, USA) 测定叶面积。然后将测量后的叶片装入信封后放入烘箱80 ℃烘72 h。用电子天平 (AR2140, Ohaus, USA) 称量叶干重。根据叶干重与叶面积比值计算比叶重 (g/cm2)。
从每个植株上截取3根10 cm左右2—3年生的枝条, 用小刀削去枝干表面的树皮, 用量筒排水法测量枝干的体积。然后将枝条装入信封后放入烘箱80 ℃烘72 h。用电子天平 (AR2140, Ohaus, USA) 称量枝干干重, 根据枝条干重与体积的比值计算WD (g/cm3)。霸王岭热带森林树木枝条WD与茎干WD有很强相关性[31], 因而能用植物枝条WD表示茎干WD, 同时也避免了年轮钻对树木茎干取样时对树木的伤害。
1.4 土壤取样和养分的测定在每个5 m×5 m样方的中心位置取土样。去掉土壤表层的枯枝落叶, 挖0.2 m深的土壤剖面, 自上而下取1 kg混合土样。共取336个土样, 标记后带到实验室处理。
将土样自然风干、过筛, 测量土壤有机质、全磷、有效磷、全氮及有效氮含量。有机质用高温外热重铬酸钾氧化-容量法测定;全磷测定是用HClO4-H2SO4消化法分解, 然后用钼锑抗比色法测定;有效磷用酸性氟化铵浸提, 然后用抗坏血酸还原比色测定。全氮用凯氏定氮法, 有效氮用碱解扩散法测定[32]。
1.5 数据分析方法(1) 功能性状在个体、种内、种间及群落尺度的分异
为了使数据正态化, 在数据分析前对功能性状值进行log10转换。运用R 3.13软件“lme”包中限制最大似然法 (Restricted maximum likelihood, REML), 用广义线性模型和“varcomp”函数对个体、种内、种间、群落水平上植物功能性状的分异大小进行方差分解。以LMA为例, 其余的功能性状只要将LMA替换即可:
varcomp.LMA < -varcomp (lme (log (LMA)—1, random=—1|Plot/Species/Tree/leaf or stem, data=d, na.action=na.omit), 1)
群落之间的功能性状的差异, 可能由于环境或物种组成不同所致。热带云雾林物种多样性较高, 物种组成对功能性状分异的影响可能会大于环境。为了分析物种组成差异, 计算不同样方间植物物种Sorenson相似性系数。计算公式为: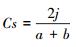
为了分析不同群落中功能性状的变化特征, 利用每个植物个体的功能性状值, 比较每个样方及所有样方中功能性状的频度分布及平均值大小。物种间的竞争、共生以及寄生等关系会影响个体、种内和群落尺度水平上功能性状的变化。为了排除种间关系的影响, 通过移除种间尺度来分析方差分异模型, 来探究个体、种内、群落尺度上植物功能性状的分异规律。
(2) 功能性状与土壤养分的关系分析
以土壤有机质、全磷、有效磷、全氮及有效氮含量为自变量, 以个体、种内、种间以及群落尺度的叶面积、叶干重、比叶重、叶绿素含量、叶厚度、木材密度为因变量, 进行逐步线性回归分析。根据AIC值、模型决定系数及P值大小选择最优模型, 选择影响植物功能性状的关键土壤因子。数据分析及作图用R Development Core Team 2009[33]。
2 研究结果 2.1 功能性状在个体、种内、种间、群落尺度上的分异21个20 m×20 m群落中, 平均物种丰富度为25.80±6.00, 最大值为34, 最小值为13;群落间的相似性系数为0.58±0.10。
总体上6种不同的功能性状值在群落尺度的解释方差最小 (图 2)。叶面积、叶干重、比叶重的分异规律较为相似, 解释方差大小皆表现为种间 (0.57—0.72)>种内 (0.21—0.30)>个体 (0.06—0.09)>群落 (0.0—0.01);叶厚度的解释方差为种间 (0.58)>种内 (0.26)>个体 (0.06)≈群落 (0.06);叶绿素解释方差为种间 (0.34)≈种内 (0.35)>个体 (0.23)>群落 (0.03);茎干密度的解释方差为个体 (0.47)>种间 (0.43)>种内 (0.09)>群落 (0.00)。6种功能性状在群落组织尺度上的分异大小表现出一致规律, 即种间>种内>群落。

|
| 图 2 功能性状在个体、种内、种间及群落水平的方差分解 Fig. 2 Variance partitioning for the six functional traits across the individual, within-species, among-species and community levels LA:叶面积leaf area;LDW:叶干重leaf dry weight;LMA:比叶重leaf mass per area;Chl:叶绿素含量leaf chlorophyll content;LTh:叶片厚度Leaf thickness;WD:木材密度wood density |
各个样方的功能性状频度分布特征及平均值大小与所有样方一致 (图 3)。移除种间尺度后, 来源于种间尺度的方差分量大部分被增加到了种内尺度, 小部分增加到了群落尺度, 但功能性状分异大小仍是种内>群落 (图 4)。

|
| 图 3 功能性状频度分布图 Fig. 3 Density distribution of functional traits 实线表示所有样地的功能性状值频度分布曲线, 虚线表示各个样地的功能性状频度分布曲线;▏:样地功能性状平均值, ●:所有样地的功能性状平均值 |

|
| 图 4 功能性状在个体、种内及群落水平的方差分解 Fig. 4 Variance partitioning for the six functional traits across the individual, within-species and community levels |
总体上, 功能性状分异与有机质、氮磷都有密切关系 (表 2)。LA在种内尺度上与土壤有机质含量相关, 在种间、群落尺度不相关;LDW在种内尺度与土壤有机质和全氮含量相关, 而在种间与有效磷、全氮相关, 群落尺度与土壤有机质含量相关;LMA在种内尺度与有机质、全磷、全氮、有效氮相关, 而在种间尺度不相关, 群落尺度与全氮相关;Chl在种内尺度与土壤中的五个因子都相关, 而种间尺度与有机质含量、全磷、有效磷相关, 群落尺度与有机质含量、全磷、有效磷、全氮相关;LTh在种内尺度有机质、全氮、有效氮相关, 在种间尺度与有机质、有效磷、有效氮相关, 在群落尺度与有效磷、全氮相关;WD在种内尺度与有效磷、全氮、有效氮相关, 而在种间尺度与有机质、全磷、有效磷相关。
| 功能性状 Functional traits |
尺度 Scale |
回归方程 Regression equation |
决定系数 R2 |
F (自由度) |
P |
| 叶面积Leaf area | 种内 | Y=2.64+0.57X1 | < 0.01 | 2.9 (1, 2186) | 0.09 |
| 种间 | — | — | — | — | |
| 群落 | — | — | — | — | |
| 叶干重 | 种内 | Y=-1.8+2.73X1-0.18X4 | 0.02 | 16.24 (2, 2096) | < 0.001 |
| Leaf dry weight | 种间 | Y=-1.6+0.04X3-0.34X4 | < 0.01 | 2.1 (2, 481) | 0.12 |
| 群落 | Y=-1.71+1.7X1 | 0.31 | 8.49 (1, 19) | 0.01 | |
| 比叶重 | 种内 | Y=-4.38+1.92X1-0.69X2-0.08X4+0.03X5 | 0.03 | 18.07 (4, 2094) | < 0.001 |
| Leaf mass per area | 种间 | — | — | — | — |
| 群落 | Y=-5.36+0.65X4 | 0.59 | 27.81 (1, 19) | < 0.001 | |
| 叶绿素含量 | 种内 | Y=4.11+0.56X1-0.15X2-0.01X3-0.03X4+0.02X5 | 0.02 | 9.84 (5, 2182) | < 0.001 |
| Leaf chlorophyll | 种间 | Y=4.14+1.15X1-0.47X2-0.01X3 | 0.03 | 4.8 (3, 496) | 0 |
| content | 群落 | Y=3.99+0.87X1-0.74X2-0.01X3+0.23X4 | 0.51 | 4.23 (4, 16) | 0.02 |
| 叶片厚度 | 种内 | Y=-1.42+1.21X1-0.11X4+0.02X5 | 0.04 | 29.8 (3, 2184) | < 0.001 |
| Leaf thickness | 种间 | Y=-1.57+0.98X1+0.02X3-0.05X5 | 0.02 | 3.51 (3, 496) | 0.02 |
| 群落 | Y=-2.22+0.02X3+0.44X4 | 0.54 | 10.39 (2, 18) | 0 | |
| 木材密度 | 种内 | Y=-0.44-0.01X3-0.04X4+0.01X5 | 0.01 | 6.99 (3, 1975) | < 0.001 |
| Wood density | 种间 | Y=-0.28+0.51X1-0.42X2-0.02X3 | 0.04 | 6.38 (3, 454) | < 0.001 |
| 群落 | — | — | — | — | |
| X1、X2、X3、X4、X5分别表示土壤有机质含量、土壤全磷含量、土壤有效磷含量、土壤全氮含量、土壤有效氮含量; Y分别表示种内、种间、群落尺度下LA, LDW, LMA, Chl, LTh, WD | |||||
6种植物功能性状在群落组织尺度上分异大小不同, 种间尺度上分异最大, 而群落尺度分异最小 (图 2—图 4)。说明种间水平的功能性状变化对植物功能性状的影响最大。种间功能性状分异来源可能与遗传及环境因素有关[8, 34], 有学者研究发现LMA、LTh和Chl有较高的系统发育保守性[35-36], 并且很多植物的功能性状都能检测到显著的系统发育信号, 如, Ackerly[37]通过调查加州树丛叶子的进化规律时发现叶子的比叶面积具有很强的系统发育信号;曹科等[38]对古田山24hm2样地中木本植物功能性状的调查研究发现, 植物样品中叶氮含量、叶磷含量、叶面积、比叶面积、木质密度和种子重量都能检测到较为显著的系统发育信号。说明植物功能性状在形成和发展过程中受到历史进化的显著影响。此外, 物种性状之间的关联也离不开系统发育的影响。例如, Ackerly和Reich[39]通过研究108种被子植物叶片性状间关联时发现, 在不同的系统发育阶段, 物种的叶片大小与叶片寿命、叶片氮含量及单位面积叶片光合能力之间的相关性有很大差异。系统发育对性状关联性的影响反映了物种性状变异与遗传相关, 这有利于植物在多变环境中通过功能协调而容易生存。另外, 6个功能性状的种间分异也可能受环境条件的强烈影响。热带云雾林分布在海拔1200m以上范围, 环境因子变化较为剧烈, 植物受空气低温和土壤低磷胁迫的影响[9], 土壤养分对种间功能性状有显著影响 (表 2)。与多数学者的观点一致, 即植物功能性状的分异主要表现在种间水平[40-41]。因而, 应用植物功能性状分析植物对环境的适应策略及物种共存机制研究中, 种间尺度的功能性状分析是研究的重点。
研究发现, 群落尺度的植物功能性状分异最小。群落尺度的功能性状分异与群落间物种周转关系密切。不同群落的物种相似性为0.58±0.10, 群落间物种较为相似, 因而对功能性状变化的影响较小。与Messier等[12]的研究一致, LMA和LDMC的含量在群落水平的解释方差最小。但是, 研究证明了群落尺度的功能性状分异是功能性状变化的重要来源。群落水平的功能性状变化能直接将生态系统中的环境因子与功能性状联系起来[20, 40], 因而是分析植物生态策略变化及植物对环境适应性的重要途径。
虽然群落尺度上功能性状分异最小, 且不同样方中物种组成和数量都有差异, 6种植物功能性状在每个样方的频度分布曲线及平均值与所有样方基本一致 (图 3)。说明在群落水平上, 环境筛不是作用于物种组成, 而可能通过作用于功能性状的总体分布来影响群落物种的组配过程。
与假设相反, 种内尺度的功能性状分异大小比种间尺度稍小, 但比个体及群落尺度上的分异大, 说明种内功能性状分异是功能性状变化的重要途径。与其他学者的研究结果一致[16-18], 种内功能性状变化对热带云雾林植物的适应性及群落物种共存机制有重要作用[9]。种内功能性状变化主要来源于表型可塑性和遗传多样性[42]。热带云雾林环境条件容易变化, 植物通过表型可塑性调整其自身属性大小, 使其适应较复杂的环境, 有助于增加物种沿环境梯度的分布范围。
3.2 土壤因子对植物功能性状的影响与假设一致, 在不同的群落组织尺度上, 虽然土壤因子对不同的植物功能性状影响不尽相同, 但个体、种内、种间及群落尺度上功能性状分异都与土壤有机质、氮、磷含量有密切关系 (表 2)。例如, 叶片功能性状在种内水平上基本都与土壤有机质含量密切相关, 而木材密度在种内水平分异主要与土壤磷和氮有关;叶片和茎干功能性状在种间水平既与土壤有机质有关, 也与氮磷有关;叶片功能性状在群落水平与有机质、氮磷都有关, 但木材密度在群落水平上与土壤养分没有显著关系。不同群落组织尺度上植物叶片和茎干功能性状与土壤养分差异, 说明物种类别、多度等影响植物对土壤养分利用。土壤有机质中含有丰富的氮 (N)、磷 (P)、钾 (K) 等元素, 这些元素在叶片干物质及组成结构形成中有重要作用, 因而叶面积和干物质含量与土壤有机质含量呈正相关 (表 2)。研究发现, 叶片LMA和LTh与土壤养分有密切关系;在土壤养分贫瘠的环境中, 植物叶片较厚, LMA小的植物能很好的适应资源丰富的环境, 而LMA大的植物在贫瘠的环境中适应性更强[8], 且往往具有更长的叶寿命[43-44]。热带云雾林土壤磷含量较低[25], 植物往往采取保守的养分利用策略, 单位面积积累的干物质较高, 因而有较大的LMA[8], 叶片厚度较大[30]。叶绿素含量的变化一方面与植物对光照的适应性有关, 另一方面也受到土壤中矿质元素尤其是氮素的影响[45], 因为缺氮一方面可能会使叶绿素的生物合成途径受阻, 另一方面可能会引起植物体内活性氧的积累, 导致叶绿素分解加快, 两方面都会减少光合色素含量, 导致光合速率降低[46], 这与本研究结果一致。植物的木材密度与土壤有机质和有效氮含量呈正相关, 与土壤磷呈负相关 (表 2)。原因可能是热带云雾林风力强劲, 树木生长速度慢, 木材密度较高;土壤磷元素影响茎干中与干物质含量有关的功能性状[47], 植物能利用有限的土壤磷合成茎干的结构物质;而有效氮能被植物能直接吸收, 用于合成木纤维、木质素等物质, 增强茎干的硬度。
总之, 通过对热带云雾林植物叶干重、叶面积、比叶重、叶片叶绿素含量、叶片厚度和木材密度等功能性状6种功能性状在个体、种内、种间和群落水平上进行方差分解, 发现种间水平上功能性状分异最大, 种内水平次之, 群落水平功能性状分异最小。种间水平和种内水平分异是功能性状变化的重要来源, 在基于功能性状的生态学研究中, 测定和分析这两个水平的功能性状值非常重要;群落水平功能性状分异虽然最小, 但也是功能性状变化的来源之一, 环境因子在群落水平上是作用于功能性状总体分布来影响植物的适应性。热带云雾林中不同群落组织尺度下, 不同水平植物功能性状的分异与土壤有机质、氮和磷含量存在密切关系。
附录
| 物种名录 | |||||
| 科名 | 拉丁科名 | 属名 | 拉丁属名 | 物种名 | 拉丁名 |
| 槭树科 | Aceraceae | 槭树属 | Acer | 十蕊槭 | Acer decandrum |
| 木兰科 | Magnoliaceae | 含笑属 | Michelia | 白花含笑 | Michelia mediocris |
| 桑科 | Moraceae | 榕属 | Ficus | 变叶榕 | Ficus variolosa |
| 樟科 | Lauraceae | 木姜属 | Litsea | 豺皮樟 | Litsea rotundifolia |
| 山矾科 | Symplocaceae | 山矾属 | Symplocos | 丛花山矾 | Symplocos poilanei |
| 金缕梅科 | Hamamelidaceae | 马蹄荷属 | Exbucklandia | 大果马蹄荷 | Exbucklandia tonkinensis |
| 山茶科 | Theaceae | 大头茶属 | Polyspora | 大头茶 | Polyspora axillaris |
| 山矾科 | Symplocaceae | 山矾属 | Symplocos | 单花山矾 | Symplocos ovatilobata |
| 壳斗科 | Fagaceae | 青冈属 | Cyclobalanopsis | 碟斗青冈 | Cyclobalanopsis disciformis |
| 樟科 | Lauraceae | 润楠属 | Machilus | 短序润楠 | Machilus breviflora |
| 五加科 | Araliaceae | 鹅掌柴属 | Schefflera | 鹅掌柴 | Schefflera heptaphylla |
| 壳斗科 | Fagaceae | 青冈属 | Cyclobalanopsis | 饭甑青冈 | Cyclobalanopsis fleuryi |
| 樟科 | Lauraceae | 润楠属 | Machilus | 芳槁润楠 | Machilus gamblei |
| 芸香科 | Rutaceae | 山油柑属 | Acronychia | 贡甲 | Acronychia oligophlebia |
| 山矾科 | Symplocaceae | 山矾属 | Symplocos | 光叶山矾 | Symplocos lancifolia |
| 冬青科 | Aquifoliaceae | 冬青属 | Ilex | 海南冬青 | Ilex hainanensis |
| 五福花科 | Adoxaceae | 荚蒾属 | Viburnum | 海南荚蒾 | Viburnum hainanense |
| 山茶科 | Theaceae | 柃木属 | Eurya | 海南柃 | Eurya hainanensis |
| 蔷薇科 | Rosaceae | 李属 | Prunus | 海南樱桃 | Prunus hainanensis |
| 山茶科 | Theaceae | 木荷属 | Schima | 荷木 | Schima superba |
| 槭树科 | Aceraceae | 槭属 | Acer | 红翅槭 | Acer lucidum |
| 壳斗科 | Fagaceae | 柯属 | Lithocarpus | 红椆 | Lithocarpus fenzelianus |
| 杜鹃花科 | Ericaceae | 珍珠花属 | Lyonia | 红脉南烛 | Lyonia rubrovenia |
| 樟科 | Lauraceae | 厚壳桂属 | Cryptocarya | 厚壳桂 | Cryptocarya chinensis |
| 山茶科 | Theaceae | 厚皮香属 | Ternstroemia | 厚皮香 | Ternstroemia gymnanthera |
| 八角科 | Illiciaceae | 八角属 | Illicium | 厚皮香八角 | Illicium ternstroemioides |
| 樟科 | Lauraceae | 琼楠属 | Beilschmiedia | 厚叶琼楠 | Beilschmiedia percoriacea |
| 壳斗科 | Fagaceae | 青冈属 | Cyclobalanopsis | 黄背青冈 | Cyclobalanopsis poilanei |
| 胡桃科 | Juglandaceae | 黄杞属 | Engelhardia | 黄杞 | Engelhardia roxburghiana |
| 远志科 | Polygalaceae | 黄叶树属 | Xanthophyllum | 黄叶树 | Xanthophyllum hainanense |
| 冬青科 | Aquifoliaceae | 冬青属 | Ilex | 剑叶冬青 | Ilex lancilimba |
| 樟科 | Lauraceae | 樟属 | Cinnamomum | 平托桂 | Cinnamomum tsoi |
| 茜草科 | Rubiaceae | 九节属 | Psychotria | 九节 | Psychotria asiatica |
| 木兰科 | Magnoliaceae | 拟单性木兰属 | Parakmeria | 乐东拟单性木兰 | Parakmeria lotungensis |
| 壳斗科 | Fagaceae | 柯属 | Lithocarpus | 犁耙柯 | Lithocarpus silvicolarum |
| 卫矛科 | Celastraceae | 假卫矛属 | Microtropis | 灵香假卫矛 | Microtropis submembranacea |
| 紫金牛科 | Myrsinaceae | 铁仔属 | Myrsine | 柳叶密花树 | Myrsine linearis |
| 紫金牛科 | Myrsinaceae | 紫金牛属 | Ardisia | 罗伞树 | Ardisia quinquegona |
| 杜鹃花科 | Ericaceae | 杜鹃花属 | Rhododendron | 毛棉杜鹃 | Rhododendron moulmainense |
| 樟科 | Lauraceae | 新木姜属 | Neolitsea | 美丽新木姜 | Neolitsea pulchella |
| 紫金牛科 | Myrsinaceae | 铁仔属 | Myrsine | 密花树 | Myrsine seguinii |
| 樟科 | Lauraceae | 润楠属 | Machilus | 绒毛润楠 | Machilus velutina |
| 壳斗科 | Fagaceae | 柯属 | Lithocarpus | 梨果柯 | Lithocarpus howii |
| 杜英科 | Elaeocarpaceae | 杜英属 | Elaeocarpus | 山杜英 | Elaeocarpus sylvestris |
| 山矾科 | Symplocaceae | 山矾属 | Symplocos | 山矾 | Symplocos sumuntia |
| 五加科 | Araliaceae | 树参属 | Dendropanax | 树参 | Dendropanax dentiger |
| 木犀科 | Oleaceae | 木犀属 | Osmanthus | 双瓣木犀 | Osmanthus didymopetalus |
| 桃金娘科 | Myrtaceae | 蒲桃属 | Syzygium | 碎叶蒲桃 | Syzygium buxifolium |
| 冬青科 | Aquifoliaceae | 冬青属 | Ilex | 凸脉冬青 | Ilex kobuskiana |
| 樟科 | Lauraceae | 琼楠属 | Beilschmiedia | 网脉琼楠 | Beilschmiedia tsangii |
| 金缕梅科 | Hamamelidaceae | 蚊母树属 | Distylium | 蚊母树 | Distylium racemosum |
| 五列木科 | Pentaphylacaceae | 五列木属 | Pentaphylax | 五列木 | Pentaphylax euryoides |
| 木犀科 | Oleaceae | 木犀属 | Osmanthus | 显脉木犀 | Osmanthus hainanensis |
| 桃金娘科 | Myrtaceae | 蒲桃属 | Syzygium | 线枝蒲桃 | Syzygium araiocladum |
| 桃金娘科 | Myrtaceae | 蒲桃属 | Syzygium | 香蒲桃 | Syzygium odoratum |
| 山茶科 | Theaceae | 红淡比属 | Cleyera | 肖柃 | Cleyera incornuta |
| 番荔枝科 | Annonaceae | 暗罗属 | Polyalthia | 斜脉暗罗 | Polyalthia plagioneura |
| 杜英科 | Elaeocarpaceae | 杜英属 | Elaeocarpus | 锈毛杜英 | Elaeocarpus howii |
| 樟科 | Lauraceae | 新木姜属 | Neolitsea | 锈叶新木姜 | Neolitsea cambodiana |
| 杨梅科 | Myricaceae | 杨梅属 | Myrica | 杨梅 | Myrica rubra |
| 夹竹桃科 | Apocynaceae | 狗牙花属 | Tabernaemontana | 药用狗牙花 | Tabernaemontana bovina |
| 漆树科 | Anacardiaceae | 漆属 | Toxicodendron | 野漆 | Toxicodendron succedaneum |
| 木犀科 | Oleaceae | 木犀榄属 | Olea | 异株木樨榄 | Olea dioica |
| 樟科 | Lauraceae | 樟属 | Cinnamomum | 银叶樟 | Cinnamomum mairei |
| 壳斗科 | Fagaceae | 锥栗属 | Castanopsis | 公孙锥 | Castanopsis tonkinensis |
| 桃金娘科 | Myrtaceae | 蒲桃属 | Syzygium | 子凌蒲桃 | Syzygium championii |
| 野牡丹科 | Melastomataceae | 野牡丹属 | Melastoma | 紫毛野牡丹 | Melastoma penicillatum |
| [1] | Ackerly D D. Community assembly, niche conservatism, and adaptive evolution in changing environments. International Journal of Plant Sciences, 2003, 164(S3): S165–S185. DOI:10.1086/368401 |
| [2] | 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150–165. |
| [3] | Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, Van Der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335–380. DOI:10.1071/BT02124 |
| [4] | Poorter L, Markesteijn L. Seedling traits determine drought tolerance of tropical tree species. Biotropica, 2008, 40(3): 321–331. |
| [5] | Westoby M, Wright I J. Land-plant ecology on the basis of functional traits. Trends in Ecology & Evolution, 2006, 21(5): 261–268. |
| [6] | McGill B J, Enquist B J, Weiher E, Westoby M. Rebuilding community ecology from functional traits. Trends in Ecology & Evolution, 2006, 21(4): 178–185. |
| [7] | Vile D, Shipley B, Garnier E. Ecosystem productivity can be predicted from potential relative growth rate and species abundance. Ecology Letters, 2006, 9(9): 1061–1067. DOI:10.1111/j.1461-0248.2006.00958.x |
| [8] | Long W X, Zang R G, Ding Y. Air temperature and soil phosphorus availability correlate with trait differences between two types of tropical cloud forests. Flora-Morphology, Distribution, Functional Ecology of Plants, 2011, 206(10): 896–903. DOI:10.1016/j.flora.2011.05.007 |
| [9] | Long W X, Zang R G, Schamp B S, Ding Y. Within-and among-species variation in specific leaf area drive community assembly in a tropical cloud forest. Oecologia, 2011, 167(4): 1103–1113. |
| [10] | Long W X, Schamp B S, Zang R G, Ding Y, Huang Y F, Xiang Y Z. Community assembly in a tropical cloud forest related to specific leaf area and maximum species height. Journal of Vegetation Science, 2015, 26(3): 513–523. |
| [11] | Cavender-Bares J, Keen A, Miles B. Phylogenetic structure of Floridian plant communities depends on taxonomic and spatial scale. Ecology, 2006, 87(S7): S109–S122. |
| [12] | Messier J, McGill B J, Lechowicz M J. How do traits vary across ecological scales? A case for trait-based ecology. Ecology Letters, 2010, 13(7): 838–848. DOI:10.1111/ele.2010.13.issue-7 |
| [13] | McGill B J. Exploring predictions of abundance from body mass using hierarchical comparative approaches. The American Naturalist, 2008, 172(1): 88–101. DOI:10.1086/588044 |
| [14] | Burns K C. Patterns in specific leaf area and the structure of a temperate heath community. Diversity and Distributions, 2004, 10(2): 105–112. |
| [15] | Cornwell W K, Ackerly D D. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecological Monographs, 2009, 79(1): 109–126. DOI:10.1890/07-1134.1 |
| [16] | Albert C H, Thuiller W, Yoccoz N G, Douzet R, Aubert S, Lavorel S. A multi-trait approach reveals the structure and the relative importance of intra-vs. interspecific variability in plant traits. Functional Ecology, 2010, 24(6): 1192–1201. DOI:10.1111/j.1365-2435.2010.01727.x |
| [17] | Paine C E T, Baraloto C, Chave J, Hérault B. Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos, 2011, 120(5): 720–727. DOI:10.1111/more.2011.120.issue-5 |
| [18] | Bolnick D I, Amarasekare P, Araújo M S, Bürger R, Levine J M, Novak M, Rudolf V H W, Schreiber S J, Urban M C, Vasseur D A. Why intraspecific trait variation matters in community ecology. Trends in Ecology & Evolution, 2011, 26(4): 183–192. |
| [19] | Lebrija-Trejos E, Pérez-García E A, Meave J A, Bongers F, Poorter L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 2010, 91(2): 386–398. |
| [20] | Swenson N G, Weiser M D. Plant geography upon the basis of functional traits:an example from eastern North American trees. Ecology, 2010, 91(8): 2234–2241. |
| [21] | Bubb P, May I, Miles L, Sayer J. Cloud Forest Agenda. Cambridge, UK: UNEP-World Conservation Monitoring Centre, 2004. |
| [22] | 龙文兴, 臧润国, 丁易. 海南岛霸王岭热带山地常绿林和热带山顶矮林群落特征. 生物多样性, 2011, 19(5): 558–566. |
| [23] | Williams-Linera G. Tree species richness complementarity, disturbance and fragmentation in a Mexican tropical montane cloud forest. Biodiversity & Conservation, 2002, 11(10): 1825–1843. |
| [24] | 施济普.云南山顶苔藓矮林群落生态学与生物地理学研究[D].西双版纳:中国科学院研究生院 (西双版纳热带植物园), 2007. |
| [25] | 龙文兴, 丁易, 臧润国, 杨民, 陈少伟. 海南岛霸王岭热带云雾林雨季的环境特征. 植物生态学报, 2011, 35(2): 137–146. |
| [26] | Zang R G, Tao J P, Li C Y. Within community patch dynamics in a tropical montane rain forest of Hainan Island, South China. Acta Oecologica, 2005, 28(1): 39–48. DOI:10.1016/j.actao.2005.02.001 |
| [27] | 胡玉佳, 李玉杏. 海南岛热带雨林. 广东: 高等教育出版社, 1992. |
| [28] | Baraloto C, Paine C E T, Patiño S, Bonal D, Hérault B, Chave J. Functional trait variation and sampling strategies in species-rich plant communities. Functional Ecology, 2010, 24(1): 208–216. |
| [29] | Chaturvedi R K, Raghubanshi A S, Singh J S. Leaf attributes and tree growth in a tropical dry forest. Journal of Vegetation Science, 2011, 22(5): 917–931. DOI:10.1111/jvs.2011.22.issue-5 |
| [30] | Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, De Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, Van Der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167–234. DOI:10.1071/BT12225 |
| [31] | Bu W S, Zang R G, Ding Y. Field observed relationships between biodiversity and ecosystem functioning during secondary succession in a tropical lowland rainforest. Acta Oecologica, 2014, 55: 1–7. DOI:10.1016/j.actao.2013.10.002 |
| [32] | 中国土壤学会农业化学专业委员会. 土壤农业化学常规分析方法. 北京: 科学出版社, 1983. |
| [33] | The R Development Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing, 2009. |
| [34] | Hughes A R, Inouye B D, Johnson M T J, Underwood N, Vellend M. Ecological consequences of genetic diversity. Ecology Letters, 2008, 11(6): 609–623. DOI:10.1111/ele.2008.11.issue-6 |
| [35] | Vasseur F, Violle C, Enquist B J, Granier C, Vile D. A common genetic basis to the origin of the leaf economics spectrum and metabolic scaling allometry. Ecology Letters, 2012, 15(10): 1149–1157. DOI:10.1111/j.1461-0248.2012.01839.x |
| [36] | Grady K C, Laughlin D C, Ferrier S M, Kolb T E, Hart S C, Allan G J, Whitham T G. Conservative leaf economic traits correlate with fast growth of genotypes of a foundation riparian species near the thermal maximum extent of its geographic range. Functional Ecology, 2013, 27(2): 428–438. DOI:10.1111/fec.2013.27.issue-2 |
| [37] | Ackerly D D. Adaptation, niche conservatism, and convergence:comparative studies of leaf evolution in the California chaparral. The American Naturalist, 2004, 163(5): 654–671. DOI:10.1086/383062 |
| [38] | 曹科, 饶米德, 余建中, 刘晓娟, 米湘成, 陈建华. 古田山木本植物功能性状的系统发育信号及其对群落结构的影响. 生物多样性, 2013, 21(5): 564–571. |
| [39] | Ackerly D D, Reich P B. Convergence and correlations among leaf size and function in seed plants:a comparative test using independent contrasts. American Journal of Botany, 1999, 86(9): 1272–1281. |
| [40] | Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets V, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821–827. |
| [41] | Chave J, Coomes D, Jansen S, Lewis S L, Swenson N G, Zanne A E. Towards a worldwide wood economics spectrum. Ecology Letters, 2009, 12(4): 351–366. DOI:10.1111/ele.2009.12.issue-4 |
| [42] | Jung V, Violle C, Mondy C, Hoffmann L, Muller S. Intraspecific variability and trait-based community assembly. Journal of Ecology, 2010, 98(5): 1134–1140. DOI:10.1111/jec.2010.98.issue-5 |
| [43] | Reich P B, Walters M B, Ellsworth D S, Vose J M, Volin J C, Gresham C, Bowman W D. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span:a test across biomes and functional groups. Oecologia, 1998, 114(4): 471–482. |
| [44] | Wright I J, Westoby M, Reich P B. Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. Journal of Ecology, 2002, 90(3): 534–543. |
| [45] | Verhoeven A S, Demmig-Adams B, Adams Ⅲ W W. Enhanced employment of the xanthophyll cycle and thermal energy dissipation in spinach exposed to high light and N stress. Plant Physiology, 1997, 113(3): 817–824. |
| [46] | 郭卫东, 桑丹, 郑建树, 廖海兵, 陈文荣. 缺氮对佛手气体交换、叶绿素荧光及叶绿体超微结构的影响. 浙江大学学报:农业与生命科学版, 2009, 35(3): 307–314. |
| [47] | Wright I J, Reich P B, Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats. Functional Ecology, 2001, 15(4): 423–434. |