 2017, Vol. 37
2017, Vol. 37文章信息
- 曹永昌, 杨瑞, 刘帅, 王紫泉, 和文祥, 耿增超
- CAO Yongchang, YANG Rui, LIU Shuai, WANG Ziquan, HE Wenxiang, GENG Zengchao.
- 秦岭典型林分夏秋两季根际与非根际土壤微生物群落结构
- Characteristics of microbial community in forest soil between rhizosphere and non-rhizosphere in summer and autumn in Qinling Mountains, China
- 生态学报. 2017, 37(5): 1667-1676
- Acta Ecologica Sinica. 2017, 37(5): 1667-1676
- http://dx.doi.org/10.5846/stxb201509151898
-
文章历史
- 收稿日期: 2015-09-15
- 网络出版日期: 2016-07-13

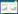




2. 金正大生态工程集团股份有限公司, 养分资源高效开发与综合利用国家重点实验室, 临沭 276700
2. Kingenta Ecological Engineering Group Co. Ltd, State Key Laboratory of Nutrition Resources Integrated Utilization, Linshu 276700, China
秦岭作为我国南北气候分界线的主要林区, 在调节气候、固碳以及保护珍稀物种等方面发挥重要作用, 且其独特的地理位置和复杂的自然环境孕育了丰富的植物资源, 在不同的海拔范围生长着典型的植物类型, 如锐齿栎、华山松、油松、云杉等[1]。根际是植物能量和物质代谢最活跃的部位之一, 其中包括的微生物数量远高于非根际土壤, 且根际微生物能对根际释放的营养物质做出响应[2-3], 其生理活动会明显影响土壤性状、植物养分吸收、生长状况[4-6], 最终导致植物群落和陆地生态系统碳循环及格局的变化, 对提高土壤肥力、改善土壤生态环境具有重要意义[7]。国内外学者从分子生物学方法研究显示林分类型会对土壤微生物群落产生影响[8], 且具有明显的季节变化特征[9];同一林分微生物对碳源优先利用种类和程度及树种间的菌根根际微生物多样性差异显著[10-11];天然次生林比人工林和荒地的土壤微生物群落多样性更高, 揭示自然恢复更有利于土壤微生物结构和功能的改善[12];秦岭地区落叶松林根际微生物数量大于松栎混交林[14], 土壤中细菌>放线菌>真菌[15]。可见一方面森林生态系统的恢复与保护不仅要考虑植物多样性, 更要关注土壤微生物多样性的特征, 其一直是土壤生态学研究难点和热点[13], 另一方面对秦岭地区根际与非根际土壤微生物特征受典型林分影响缺乏系统研究[16]。因此有必要对秦岭山地土壤微生物特征进行探讨, 揭示该地区森林生态系统属性过程的季节变化规律并进行科学预测, 最终为秦岭地区针叶林和阔叶林森林生态系统的保护和合理综合开发利用提供科学依据。
本文以秦岭4种典型林分为对象, 采用BIOLOG方法研究夏、秋两季根际与非根际土壤微生物功能多样性变化特征, 以期探明:(1) 典型林分下根际与非根际土壤微生物主要碳源类型及其代谢特征;(2) 夏秋两季对4种林分土壤微生物碳源类型及其代谢特征的影响;(3) 分析土壤微生物与环境因子之间的内在联系。最终为定向调控秦岭森林的发育和森林生态系统的保护提供依据。
1 材料与方法 1.1 试验材料在秦岭山脉中选取马头滩林场的锐齿栎 (Quercus alienavar.acutidentata)、油松Pinus tabuliformis)、华山松 (Pinus armandii) 以及辛家山林场的云杉 (Picea asperata)4个典型林地 (表 1)。每个林地中选取20 m×20 m三个标准样方, 分别于夏 (S) 秋 (A) 两季无菌操作方法采集根际 (R) 与非根际 (N) 土样。其中根际土:在每个样方内选取3株树龄相近的树木, “品字形”采集0—10 cm剖面且距根表小于5 mm的土壤样品;非根际土:在每个样方先去除表层枯枝落叶, 然后5个采样点采集0—10 cm土样。将每个样方采集土样混匀后, 部分置于4℃冰箱保存测定土壤微生物多样性;其余土壤风干, 过筛, 常规方法分析土壤pH、有机质、全氮、碱解氮、速效磷、速效钾、缓效钾等基本理化性质[17]。
| 编号 No. | 植被类型 Vegetation type | 林地密度 Forest density/ (株/m2) | 灌木密度 Shrub density/ (株/m2) | 草本密度 Herb density/ (株/m2) | 凋落物量 Litter fall/ (t/hm2) | 经度 Latitude | 纬度 Longitude |
| TO | 锐齿栎 | 0.065 | 0.383 | 0.258 | 2.761 | E 106°59′ 30.7″ | N 34°13′ 34.5 ″ |
| CP | 油松 | 0.055 | 0.535 | 0.675 | 2.190 | E 106°59′ 12.3″ | N 34°13′ 26.5 ″ |
| AP | 华山松 | 0.151 | 0.528 | 0.96 | 6.068 | E 106°59′ 3.7″ | N 34°13′ 14.0″ |
| SP | 云杉 | 0.053 | 0.470 | 1.673 | 9.960 | E 106°31′ 38.0″ | N 34°16′ 51.4″ |
取过2mm筛相当5g风干土的新鲜土样, 加入45 mL0.85%灭菌盐水, 振荡 (200 r/min)30 min, 制备10-1土壤悬液, 然后系列稀释至10-3, 吸取150μL, 接种至ECO生态测试板, 加盖, 在 (25±1) ℃下培养120 h, 每隔12 h在590 nm下测定读数。其中31种单一碳源分为6大类[18]:氨基酸类 (6种)、羧酸类 (5种)、糖类 (12种)、胺类 (2种)、多聚物类 (4种) 和芳香化合物类 (2种)[19]。
1.3 数据处理 1.3.1 平均颜色变化率 (AWCD)
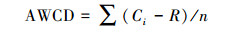
|
式中, Ci为各反应孔的光密度值;R为ECO板对照孔的光密度值, Ci-R小于零的孔, 计算中记为零, 即Ci-R≥0, n=31。
1.3.2 Gompertz方程曲线[20]。
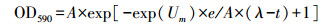
|
式中, A为最大吸光度, λ到达指数增长期所需时间 (时滞相), Um为最大吸光度变化率, t为培养时间。
1.3.3 群落丰富度指数 (S)用碳源代谢孔的数目 (AWCD> 0.2, 则代表该孔碳源被利用, 该孔即为反应孔) 表示群落丰富度指数。
1.3.4 Shannon多样性指数 (H′) 及McIntosh多样性指数 (U):
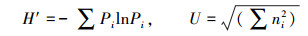
|
式中, Pi为每一孔吸光度除以所有孔吸光度的和。
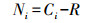
|
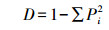
|
数据分析采用Microsoft Excel 2003进行基础数据处理, 采用SPSS 12.0对平均颜色变化率和培养时间进行方程拟合, 对培养72 h时土壤微生物群落多样性指数和对碳源的利用进行单因素方差分析, 对培养72 h时31种碳源的平均吸光度进行主成分分析 (PCA), 采用CANOCO 4.5对土壤基本理化性质和31种碳源的平均吸光度进行冗余分析 (RDA)。
2 结果 2.1 土壤微生物平均颜色变化率的变化平均颜色变化率 (AWCD) 表征微生物群落碳源利用率, 是土壤微生物群落利用单一碳源能力的一个重要指标, 反映了土壤微生物活性、微生物群落生理功能多样性。供试土壤AWCD值见图 1, 可看出:随培养时间延长AWCD增长, 在24 h之前AWCD很小, 低于0.018, 说明此时碳源基本未被利用;24—120 h AWCD快速升高, 反映24 h后碳源被大幅利用。相同培养时间下, 夏、秋季土壤根际与非根际的AWCD值截然不同, 除秋季云杉外, 其余处理均表现为非根际>根际。季节对4种林分的AWCD有明显影响, 其中锐齿栎和华山松为夏季>秋季;油松则相反, 而云杉根际土秋季>夏季, 非根际土夏季>秋季。

|
| 图 1 4种林区平均颜色变化率 Fig. 1 AWCD (average well color development) of four treatments TO、CP、AP、SP表示锐齿栎、油松、华山松和云杉, S表示夏季, A表示秋季, N表示非根际土壤, R表示根际土壤 |
将AWCD值与时间按Gompertz方程曲线进行拟合, 决定系数R2均大于0.974(表 2), 反映可较好表征二者间关系。最大吸光度变化率 (Um) 揭示微生物对碳源的最快利用速率, 表 2可看出绝大多数处理表现为非根际土对碳源利用率快 (云杉秋季除外);季节影响为锐齿栎、华山松和云杉的非根际土夏季快, 其余处理相反。最大吸光度A反映微生物对碳源利用能力, 油松、锐齿栎 (夏) 和云杉 (夏) 表现为非根际土壤利用能力强, 其余处理根际土壤利用能力强;季节变化为油松、锐齿栎 (非根际)、云杉 (非根际) 以及华山松 (根际) 秋季强, 其余处理则相反。时滞相 (λ) 为到达指数增长期所需时间, 即微生物快速利用碳源的能力, 绝大多数处理均表现为非根际所需时间短 (除夏季油松外);季节变化为锐齿栎与油松的非根际、华山松的根际秋季时间短, 其余处理夏季时间短。
| 植被 Vegetation | 编号 Numbering | OD590=A×exp[-exp (Um×e×1/A×(λ-t)+1)] | |||
| A | Um(×10-2) | λ | R2 | ||
| 锐齿栎 | TO-S-N | 1.20 | 2.10 | 35.43 | 0.999 |
| TO-S-R | 0.55 | 0.80 | 41.70 | 1.000 | |
| TO-A-N | 1.06 | 1.20 | 32.46 | 0.993 | |
| TO-A-R | 1.18 | 0.60 | 50.28 | 0.974 | |
| 油松 | CP-S-N | 1.18 | 1.50 | 39.60 | 0.999 |
| CP-S-R | 0.71 | 1.00 | 35.32 | 0.995 | |
| CP-A-N | 1.83 | 2.10 | 29.03 | 0.989 | |
| CP-A-R | 1.06 | 1.20 | 41.01 | 0.997 | |
| 华山松 | AP-S-N | 1.23 | 2.80 | 31.30 | 0.998 |
| AP-S-R | 1.10 | 1.40 | 36.71 | 0.997 | |
| AP-A-N | 1.37 | 2.00 | 31.43 | 0.998 | |
| AP-A-R | 1.03 | 1.30 | 35.24 | 0.995 | |
| 云杉 | SP-S-N | 1.23 | 1.80 | 33.89 | 0.996 |
| SP-S-R | 0.95 | 1.20 | 36.23 | 0.999 | |
| SP-A-N | 0.83 | 0.80 | 31.49 | 0.986 | |
| SP-A-R | 1.28 | 1.90 | 35.11 | 0.999 | |
| TO、CP、AP、SP表示锐齿栎、油松、华山松和云杉, S表示夏季, A表示秋季, N表示非根际土壤, R表示根际土壤 | |||||
土壤微生物群落功能多样性是土壤微生物群落状态与功能的指标, 反映土壤中微生物的生态特征。不同林分的微生物群落Shannon指数 (H)、丰富度指数 (S)、McIntosh指数 (U)、Gini指数 (D) 见表 3。不同林分表现出不同的多样性指数, 其中锐齿栎均为非根际>根际土, 夏季>秋季;油松、华山松和云杉均表现为夏季非根际>根际, 秋季则非根际 < 根际, 非根际秋季降低, 根际秋季升高。
| 植被 | 编号 | 平均颜色变化率 | 丰富度指数 (S) | Shannon指数 (H) | McIntosh指数 (U) | Gini指数 (D) |
| Vegetation | Numbering | AWCD | Richness index | Shannon index | McIntosh index | Gini index |
| 锐齿栎 | TO-S-N | 0.76±0.009a | 19.33±0.333a | 2.92±0.008a | 5.83±0.045a | 0.939±0.001a |
| TO-S-R | 0.23±0.009c | 11.66±0.333c | 2.53±0.038c | 2.26±0.062c | 0.905±0.003b | |
| TO-A-N | 0.48±0.014b | 17.00±0.000b | 2.75±0.029b | 3.81±0.030b | 0.941±0.001a | |
| TO-A-R | 0.17±0.009d | 11.00±0.000c | 2.67±0.021b | 1.57±0.070d | 0.915±0.006b | |
| 油松 | CP-S-N | 0.46±0.003b | 16.00±0.000b | 2.84±0.029b | 3.79±0.026b | 0.932±0.002b |
| CP-S-R | 0.36±0.007c | 15.66±0.333b | 2.83±0.022b | 2.96±0.035c | 0.932±0.001b | |
| CP-A-N | 0.33±0.011d | 14.00±0.000c | 2.73±0.089b | 3.03±0.033c | 0.915±0.006c | |
| CP-A-R | 0.95±0.015a | 21.00±0.000a | 3.01±0.008a | 7.02±0.090a | 0.943±0.000a | |
| 华山松 | AP-S-N | 0.96±0.006a | 20.66±0.333a | 2.97±0.003a | 7.25±0.073a | 0.941±0.001a |
| AP-S-R | 0.49±0.003c | 17.00±0.000b | 2.77±0.018b | 4.36±0.005c | 0.920±0.001c | |
| AP-A-N | 0.49±0.003c | 15.00±0.000c | 2.73±0.022b | 4.18±0.015d | 0.925±0.001b | |
| AP-A-R | 0.78±0.004b | 21.00±0.000a | 2.99±0.003a | 5.94±0.035b | 0.940±0.000a | |
| 云杉 | SP-S-N | 0.67±0.002a | 19.00±0.000a | 2.80±0.012b | 5.78±0.073a | 0.923±0.002b |
| SP-S-R | 0.35±0.010d | 16.33±0.333b | 2.66±0.024c | 3.44±0.100c | 0.901±0.004c | |
| SP-A-N | 0.41±0.007c | 17.00±0.000b | 2.80±0.009b | 3.30±0.035c | 0.933±0.001a | |
| SP-A-R | 0.60±0.014b | 18.66±0.333a | 2.96±0.021a | 4.73±0.109b | 0.937±0.002a | |
| 同列不同字母表示差异显著 (P<0.05) | ||||||
土壤微生物对不同碳源的利用反映了微生物的代谢功能类群。对培养72h时6类碳源利用情况进行分析发现, 4种林分对6类碳源的利用规律与多样性指数基本一致。但各林分随季节的优势和弱势碳源截然不同。4种林分优势碳源依次为氨基酸类、多聚物类和胺类、糖类和多聚物类、糖类及氨基酸类;弱势碳源依次为羧酸类和芳香类、芳香类和羧酸类、芳香类与羧酸类、芳香类。
2.4 微生物群落功能主成分分析为探讨不同典型林分下对土壤微生物特性起主要作用的因子, 对培养72h的吸光度值进行主成分分析 (PCA), 结果表明获得的4种林分前两个主成分累积方差贡献率为76.97%—95.58%, 因此以前2个主成分PC1、PC2来分析细菌群落功能多样性[8]。结果 (图 2) 可见, 不同林分4个处理在主成分坐标体系中分布差异十分明显;锐齿栎在PC1上可将根际与非根际区分开来;在PC2上将非根际 (秋) 和根际 (夏) 予以区分。油松、华山松及云杉规律相似, 通过PC1将根际 (秋) 与其它3个处理区别开来, 通过PC2把非根际 (夏)、根际 (夏)、秋季进行了区分。

|
| 图 2 不同林木土壤细菌群落主成分分析 Fig. 2 Principal component analysis of soil bacteria communities for different tree species |
对主成分得分进一步分析, 4种林分优势利用碳源不同, 其中PC1轴上表现为:锐齿栎是腐胺和D-苹果酸;油松是D-甘露醇和N-乙酰-D葡萄糖氨;华山松是L-丝氨酸和D-苹果酸;云杉是L-丝氨酸和D-葡糖胺酸。4种林分PC2轴上优势利用碳源依次是D-半乳糖酸γ-内酯和丙酮酸甲酯、α-环式糊精和D-葡糖胺酸、吐温80和腐胺、L-苯丙氨酸和苯乙胺。
计算4种林分主成分的综合得分发现不同林分季节间的主成分综合得分差异明显 (表 4)。锐齿栎为非根际大于根际, 夏季>秋季;油松、华山松和云杉规律相同, 为夏季非根际较大, 秋季非根际较小;季节对综合得分的影响为:根际夏季大, 秋季小。对综合得分与多样性指数进行相关性分析发现, 综合得分与多样性指数间的相关系数为0.596*—0.993* *, 均呈现显著或极显著正相关。
| 植被 | 编号 | 综合得分 | 植被 | 编号 | 综合得分 |
| Vegetation | Numbering | comprehensive scores | Vegetation | Numbering | Comprehensive scores |
| 锐齿栎 | TO-S-N | 16.04 | 华山松 | AP-S-N | 9.67 |
| TO-S-R | -9.07 | AP-S-R | -7.39 | ||
| TO-A-N | 5.89 | AP-A-N | -4.21 | ||
| TO-A-R | -12.86 | AP-A-R | 1.92 | ||
| 油松 | CP-S-N | -2.64 | 云杉 | SP-S-N | 5.13 |
| CP-S-R | -8.25 | SP-S-R | -10.30 | ||
| CP-A-N | -11.81 | SP-A-N | -0.36 | ||
| CP-A-R | 22.69 | SP-A-R | 5.53 |
冗余分析可进一步确定土壤理化性质对样地土壤微生物群落碳源利用模式的影响。结果 (图 3) 表明, 土壤理化性质对土壤微生物群落功能多样性的影响显著。不同林分对土壤中微生物群落碳源利用模式影响的环境因素不同。锐齿栎表现为有机质、碱解氮、速效磷以及碳氮比与微生物对碳源利用呈正相关;油松则表现为pH、有机质、全氮、碱解氮、速效磷、速效钾和碳氮比均与微生物碳源利用呈正相关;华山松基本理化性质对微生物碳源利用促进作用较小, 仅碳氮比和速效磷与微生物碳源利用呈正相关;云杉则表现为有机质、全氮、碱解氮、速效磷和速效钾均与绝大多数碳源呈正相关。

|
| 图 3 31个单一碳源与环境因子的冗余分析 Fig. 3 Redundancy analysis of 31 sole-carbon sources and environmental variables 实线箭头表示31种碳源, 虚线箭头表示环境因子 |
在本研究中, 同一季节, 根际土壤与非根际土壤微生物功能多样性指数有很大差别, 季节变化对根际与非根际土壤微生物功能多样性指数产生不同影响。夏季4种林分根际土壤微生物功能多样性指数均低于非根际, 主要原因是该地区雨季主要集中在夏季, 且温度高, 高温高湿是土壤微生物生长和繁殖的有利条件, 在一定条件下根系分泌物的选择作用也会导致根际土壤微生物功能多样性指数要小于非根际土壤, 即根际的负效应。此外杉木根际分泌物中, 酚类物质和香草醛能引起杉木的自毒作用[21], 栎林根系分泌物对固氮菌有抑制作用, 均使根际微生物的数量低于非根际[22-23]。且夏季林区草本科植物以及灌木密度均远远高于林地密度。秋季锐齿栎根际土壤微生物功能多样性指数低于非根际, 但其他林分秋季的变化相反, 这主要是由于林分类型不同造成的。如锐齿栎在0—10cm的表层土壤中须根系较少, 主根发达, 且锐齿栎林中的灌木旺盛, 对土壤的微生物多样性影响较大;而油松、华山松和云杉在0—10cm的表层土壤中须根系较锐齿栎林发达, 根系旺盛, 这就造成了根系分泌物和脱落物的种类以及数量较大, 且发达须根系也会保护根际土壤, 减少外部环境变化对土壤微生物功能多样性的影响。正如Sugden发现的土壤微生物多样性指标越高, 生态系统抵御外界干扰的能力越强[24]。而且与锐齿栎相比, 这3种针叶林灌木相对较少, 主要是草本科植物, 秋季草本科植物根系对土壤影响较少。
不同林分下土壤微生物对6类碳源的利用差异显著, 锐齿栎的优势群落主要为氨基酸和多聚物代谢群, 油松的主要是氨基酸和胺类代谢群, 华山松和云杉的主要是糖类和氨基酸类代谢群。4种林分的弱势群落主要是羧酸类和芳香类代谢群。4种林分中华山松林地密度最高, 油松灌木密度最高, 云杉草本密度和凋落物量最高, 锐齿栎灌木和草本密度最低。这也佐证了微生物群落结构和功能的差异与不同优势树种的枯枝落叶的量和生物化学组成有关, 同时与根系分泌物密切相关[25-26]。因为根系分泌物可通过改善根际养分环境提高养分的生物有效性, 因此, 林木根系对其周围的土壤微生物的数量和活性具有重要的影响[14, 27]。
冗余分析揭示土壤基本理化性质对土壤微生物碳源利用影响显著。这些环境因子的差异所导致的环境异质性与土壤微生物群落有直接的关系。在4种林分夏、秋两季根际与非根际中, 土壤碱解氮、速效磷与土壤中微生物碳利用关系密切。这是由于森林土壤中碳含量远远高于氮含量, 碱解氮含量较低, 并且4种林分中土壤速效磷含量很低, 碱解氮以及速效磷成为土壤中微生物生长繁殖的限制因素, 与土壤微生物生长息息相关。在4种林分中, pH与绝大多数碳源利用呈负相关, 这可能是由于森林土壤呈酸性, 酸性条件抑制了土壤细菌种群对碳源的利用[28]。
4 结论综上所述, 研究表明夏、秋两季4种林分土壤微生物功能多样性表现为锐齿栎根际小于非根际, 而其它针叶林则为夏季根际和秋季非根际的较小;不同季节下的土壤微生物功能多样性差异显著, 其中锐齿栎为秋季低于夏季, 其他林分为非根际土秋季低于夏季, 而根际土相反;主成分分析显示4种林分夏、秋两季根际与非根际主成分综合得分显著不同, 且计算得到的综合得分与多样性指数达显著或极显著正相关, 非根际土壤综合得分中锐齿栎最高, 其次是华山松和云杉, 油松最低;根际土夏季华山松和油松较高, 云杉最低, 秋季油松和云杉较高, 锐齿栎最低。冗余分析表明土壤理化性质的综合作用对土壤微生物群落功能多样性有显著影响。
| [1] | 赵永华, 雷瑞德, 何兴元, 贾夏. 秦岭锐齿栎林种群生态位特征研究. 应用生态学报, 2004, 15(6): 913–918. |
| [2] | Marilley L, Vogt G, Blanc M, Aragno M. Bacterial diversity in the bulk soil and rhizosphere fractions of Lolium perenne and Trifolium repens as revealed by PCR restriction analysis of 16S rDNA. Plant and Soil, 1998, 198(2): 219–224. DOI:10.1023/A:1004309008799 |
| [3] | Jjemba P K, Alexander M. Possible determinants of rhizosphere competence of bacteria. Soil Biology and Biochemistry, 1999, 31(4): 623–632. DOI:10.1016/S0038-0717(98)00168-0 |
| [4] | 纪婧琦, 夏志洁, 司红丽, 戴美学. 不同品种及农作方式对盐碱地甘薯根际微生物的影响. 山东科学, 2013, 26(1): 22–27. |
| [5] | 陈凯, 王智文, 刘训理, 孙海新, 薛东红, 吴凡. 圆孢芽孢杆菌A95的分离与鉴定. 蚕业科学, 2006, 32(2): 211–214. |
| [6] | 王超, 吴凡, 刘训理, 刘兵. 不同肥力条件下烟草根际微生物的初步研究. 中国烟草科学, 2005, 26(2): 12–14. |
| [7] | 徐文静, 靳晓东, 杨秋生. 植物根际微生物的影响因素研究进展. 河南农业科学, 2014, 43(5): 6–12. |
| [8] | Grayston S J, Wang S, Campbell C D, Edwards A C. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biology and Biochemistry, 1998, 30(3): 369–378. DOI:10.1016/S0038-0717(97)00124-7 |
| [9] | 肖育贵, 胡震宇. 不同林型土壤微生物种群数量及养分变化分析. 四川林业科技, 1997, 18(4): 32–35. |
| [10] | 张海涵, 唐明, 陈辉. 黄土高原典型林木根际土壤微生物群落结构与功能特征及其环境指示意义. 环境科学, 2009, 30(8): 2432–2437. |
| [11] | 张海涵, 唐明, 陈辉, 杜小刚. 黄土高原5种造林树种菌根根际土壤微生物群落多样性研究. 北京林业大学学报, 2008, 30(3): 85–90. |
| [12] | 郑华, 欧阳志云, 王效科, 方治国, 赵同谦, 苗鸿. 不同森林恢复类型对土壤微生物群落的影响. 应用生态学报, 2004, 15(11): 2019–2024. |
| [13] | Zhang B, Yang Y S, Zepp H. Effect of vegetation restoration on soil and water erosion and nutrient losses of a severely eroded clayey Plinthudult in southeastern China. Catena, 2004, 57(1): 77–90. DOI:10.1016/j.catena.2003.07.001 |
| [14] | 田呈明, 刘建军, 梁英梅, 刘永华. 秦岭火地塘林区森林根际微生物及其土壤生化特性研究. 水土保持通报, 1999, 19(2): 19–22. |
| [15] | 崔芳芳, 刘增文, 付刚, 段而军, 高文俊. 秦岭山区几种典型森林的土壤微生物特征及其对人为干扰的响应. 西北林学院学报, 2008, 23(2): 129–134. |
| [16] | Cheng F, Peng X B, Zhao P, Yuan J, Zhong C G, Cheng Y L, Cui C, Zhang S X. Soil microbial biomass, basal respiration and enzyme activity of main forest types in the Qinling Mountains. PLoS One, 2013, 8(6): e67353. DOI:10.1371/journal.pone.0067353 |
| [17] | 鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000. |
| [18] | Papatheodorou E M, Efthimiadou E, Stamou G P. Functional diversity of soil bacteria as affected by management practices and phenological stage of Phaseolus vulgaris. European Journal of Soil Biology, 2008, 44(4): 429–536. DOI:10.1016/j.ejsobi.2008.06.002 |
| [19] | Kong W D, Zhu Y G, Fu B J, Marschner P, He J Z. The veterinary antibiotic oxytetracycline and Cu influence functional diversity of the soil microbial community. Environmental Pollution, 2006, 143(1): 129–137. DOI:10.1016/j.envpol.2005.11.003 |
| [20] | Verschuere L, Fievez V, Van Vooren L, Verstraete W. The contribution of individual populations to the Biolog pattern of model microbial communities. FEMS Microbiology Ecology, 1997, 24(4): 353–362. DOI:10.1111/j.1574-6941.1997.tb00452.x |
| [21] | Huang Z Q, Liao L P, Wang S L, Cao G Q. Allelopathy of phenolics from decomposing stump-roots in replant Chinese fir woodland. Journal of Chemical Ecology, 2000, 26(9): 2211–2219. DOI:10.1023/A:1005580718765 |
| [22] | 蒋秋怡, 叶仲节, 钱新标, 姜培坤. 杉木根际土壤微生物和酶活性初探. 土壤, 1993, 25(5): 271–273. |
| [23] | 杨玉盛, 何宗明, 邹双全, 俞新妥. 格氏栲天然林与人工林根际土壤微生物及其生化特性的研究. 生态学报, 1998, 18(2): 198–202. |
| [24] | Sugden A M. Diversity & ecosystem resilience. Science, 2000, 290(5490): 233–235. |
| [25] | Johansson M B. The chemical composition of needle and leaf litter from Scots pine, Norway spruce and white birch in Scandinavian forests. Forestry, 1995, 68(1): 49–62. DOI:10.1093/forestry/68.1.49 |
| [26] | Grayston S J, Campbell C D. Functional biodiversity of microbial communities in the rhizospheres of hybrid larch (Larix eurolepis) and Sitka spruce (Picea sitchensis). Tree Physiology, 1996, 16(11-12): 1031–1038. DOI:10.1093/treephys/16.11-12.1031 |
| [27] | 焦如珍, 杨承栋. 不同代杉木人工林根际及非根际土壤微生物数量及种类的变化. 林业科学研究, 1999, 12(1): 13–18. |
| [28] | 胡婵娟, 傅伯杰, 刘国华, 靳甜甜, 刘宇. 黄土丘陵沟壑区典型人工林下土壤微生物功能多样性. 生态学报, 2009, 29(2): 727–733. |