 2017, Vol. 37
2017, Vol. 37文章信息
- 孙明, 柴雨, 董婧, 付志璐, 刘治更, 林建齐, 王佳晗
- SUN Ming, CHAI Yu, DONG Jing, FU Zhilu, LIU Zhigeng, LIN Jianqi, WANG Jiahan.
- 环境因子对海月水母螅状体存活与繁殖的影响
- Effects of environmental factors on polyp survival and reproduction of Aurelia sp. 1
- 生态学报. 2017, 37(4): 1309-1317
- Acta Ecologica Sinica. 2017, 37(4): 1309-1317
- http://dx.doi.org/10.5846/stxb201606301323
-
文章历史
- 收稿日期: 2016-06-30
- 修订日期: 2016-11-04





2. 广西民族大学海洋与生物技术学院, 南宁 530008;
3. 大连海洋大学海洋科技与环境学院, 大连 116023
2. Guangxi University for Nationalities, School of Marine Science and Biotechnology, Nanning 530008, China;
3. Dalian Ocean University, School of Marine Sea-Technology and Environment, Dalian 116023, China
全球变化背景下, 海洋生态系统的结构与功能也发生了巨大变化, 水母灾害已成为全球范围的一种新型海洋生态灾害[1-2]。海月水母(Aurelia spp.)作为主要致灾水母种类之一, 自20世纪末起, 在全球许多海域出现了异常的高频度暴发现象, 诸多关于海月水母暴发性增殖给海洋生态和渔业经济带来负面影响的研究被报道[3-5]。2007年, 海月水母在我国烟台、威海海域暴发, 影响了沿岸旅游业, 2009年, 海月水母大量发生几乎导致青岛发电厂机组停机[6]。近10年间, 海月水母在辽东湾的地理分布范围越来越广, 数量显著增加[7], 2016年7月, 绥中沿岸止锚湾渔港码头海月水母暴发(图 1) 。此外, 海月水母频繁入侵沿海池塘养殖区, 与养殖品种争食、争空间, 破坏水质, 影响养殖渔业生物的生长。海月水母暴发已给我国沿岸渔业、养殖业、工业和旅游业造成严重危害[8]。

|
| 图 1 2016年7月止锚湾渔港码头海月水母暴发 Fig. 1 Mass occurrence of Aurelia sp.1 in Zhimao Bay in July, 2016 |
海月水母对环境的适应能力较强, 广泛分布于全球各沿海水域。Dawson等报道了世界各地分布的海月水母存在多个亚种[9-10]。基因型不同等原因使海月水母的众多报道中出现了种群生态特性差异, 不同海域的海月水母有着截然不同的生态习性, 例如:许多研究表明, 盐度对海月水母螅状体种群数量的影响不显著, 但芬兰海域的海月水母暴发却被归咎于盐度上升产生了更适合于螅状体繁殖的环境条件[11-12]。Dong等和李玉龙等均报道了我国黄渤海分布的海月水母为Aurelia sp.1, 但目前针对我国这一亚种的室内生态学研究较少[13-14], 仅有的研究均为针对两种环境条件下的少量梯度展开的实验, 且多侧重研究的是温度和饵料对横裂生殖的影响, 例如王彦涛等、Wang等和石岩等均研究了温度和饵料条件对其个体繁殖和横裂生殖的影响[15-17], Wang等研究了3种温度条件下食物种类对其无性繁殖的影响[18], 而有关各种单一环境因子对这一亚种螅状体存活与繁殖影响的系统研究未见报道。因为螅状体阶段的死亡率和繁殖量是决定水母体种群数量的关键因素, 详细研究底栖螅状体阶段的生态习性对于准确预测水母暴发是十分重要的[19]。本文以我国黄渤海分布的海月水母螅状体为实验对象, 结合黄渤海自然水域各环境条件的实际范围设置实验梯度, 详细研究了温度、盐度、光照强度和饵料量对其存活与繁殖的影响, 旨在为进一步研究海月水母暴发机理、预测潜在分布范围以及预警预报模型的建立提供重要的基础数据。
1 材料与方法 1.1 材料试验用螅状体由辽宁省海洋生物资源与生态学重点实验室人工培育, 为同一批次。亲体采自大连沿岸黄海北部水域。常规培养螅状体的海水取自大连市黑石礁海岸, 经黑暗沉淀并沙滤后使用, 盐度 31±1, pH 8.2±0.1, 溶解氧大于5 mg/L, 水温随室温自然波动(5—25 ℃)。暂养期间, 投喂刚孵化出的优质卤虫无节幼体(Artemia nauplius)。
1.2 试验方法各环境因素试验均为每个梯度分别设3个平行, 每个平行20个螅状体, 每组试验螅状体培养于200 mL玻璃皿中。
1.2.1 温度试验根据黄渤海区海水全年温度范围将水温设11个梯度, 即0、2.5、5、7.5、10、12.5、15、17.5、20、22.5、25 ℃。螅状体起始培养温度均为15 ℃, 试验前每隔1 d升温或降温2 ℃直至达到各试验组所需温度。试验在黑暗条件下进行60 d, 培养海水盐度为28±0.5, 用智能光照培养箱(型号:GXZ-280B;GXZ-280C)控制试验水温。各试验组螅状体每隔2 d充分投喂1次, 投饵2 h后换水。为减小横裂生殖对个体繁殖速度的影响, 温度试验于春季多数螅状体(约80%)度过横裂生殖期后进行。
1.2.2 盐度试验共设14个梯度, 即7.5、10、12.5、15、17.5、20、22.5、25、27.5、30、32.5、35、37.5、40。试验在室内自然温度条件下进行(15—25 ℃), 光照为室内自然光(0—1050 lx), 共进行30 d试验。梯度海水用自然海水、粗盐和经24 h曝气后的自来水配制而成, 用PAL-06S型盐度计标定海水盐度(范围0—100, 精度0.1%), 并经24 h充氧, 稳定48 h后使用。各试验组螅状体每隔2 d充分投喂1次, 投饵2 h后换水。
1.2.3 光照强度试验共设5组, 即0、200、400、600、800 lx。试验在智能光照培养箱(型号:GXZ-280B;GXZ-280C)中进行60 d, 光周期10 L:14 D, 黑暗组用遮光布实现光照度为0的条件。海水盐度范围30—31, 水温随室温(20—25 ℃)变化。各试验组螅状体每隔3 d充分投喂一次, 投饵2 h后换水。
1.2.4 饵料量试验共设5组, 即1次/1 d、1次/2 d、1次/8 d、1次/16 d、无投喂组。试验用新孵化出的卤虫无节幼体作饵料, 每次投喂时用吸有卤虫的吸管瞄准螅状体投喂, 用特制的拨针将未被螅状体触手捉住的卤虫拨至螅状体口柄处, 保证每个螅状体摄食10个卤虫, 按每个卤虫约0.83 μg C 计算[20], 每天的饵料投喂量分别为8.3、4.2、1.0、0.5 μg C d-1和0 μg C d-1。试验在黑暗条件、室内自然温度(15—25 ℃)下进行60 d, 培养海水盐度为28±0.5。各试验组螅状体每次充分投饵2 h后换水。
1.3 指标测定与数据统计分析海月水母螅状体柄径增长值作为生长指标。螅状体柄径长指口面观圆周最大水平面的直径长, 当螅状体不为圆形时取直径最大值与最小值的平均值为螅状体柄径长。
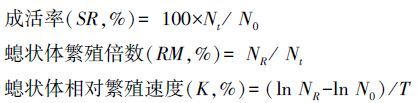
|
式中, N0为试验开始时的亲代螅状体个数, Nt为存活的亲代螅状体个数, NR为试验结束时螅状体的总个数, T为试验时间(d)。试验数据用平均数±标准差(Mean±SD)表示, 用Excel 2003对数据进行处理, 用SPSS 19.0软件对数据进行单因子方差分析。
2 结果与分析 2.1 温度对海月水母螅状体存活与种群数量繁殖的影响温度试验期间, 各试验组螅状体成活率均为100%, 但0 ℃和2.5 ℃组中的螅状体一直触手卷缩, 不摄食, 处于“冬眠”状态, 没有无性繁殖行为。各试验组60 d内未出现横裂生殖现象, 但12.5—17.5 ℃组中的螅状体个体形状纵向拉长, 有横裂前期的表征。5—25 ℃范围, 螅状体繁殖倍数和相对繁殖速度随温度的升高呈现上升趋势(图 2, 图 3) , 温度25 ℃组螅状体的平均相对繁殖速度最高, 亲代螅状体经60 d时间繁殖出的子代螅状体个数范围为7—46个, 平均为22.3个。由图 1和图 2可看出, 5—12.5 ℃范围, 海月水母螅状体繁殖倍数和相对繁殖速度的上升趋势不明显(P=0.056) , 12.5—25 ℃范围, 海月水母螅状体繁殖倍数和相对繁殖速度的上升趋势较明显(P<0.01) 。螅状体繁殖倍数和相对繁殖速度与温度呈线性相关, 螅状体繁殖倍数与温度的关系为y=0.9905x-5.3747 (n=9, r2=0.8528, P<0.01) , 螅状体相对繁殖速度与温度的关系为y=0.1797x+0.5558 (n=9, r2=0.918, P<0.01) 。方差分析结果表明, 温度对螅状体繁殖倍数和相对繁殖速度的影响极显著(P<0.01) 。综上, 本试验认为, 海月水母螅状体在0—25 ℃范围内均可存活, 在5—25 ℃范围内可正常生长并进行无性繁殖, 温度越高, 海月水母螅状体经无性繁殖出的新螅状体数量越多, 速度越快。

|
| 图 2 温度对海月水母螅状体繁殖倍数的影响 Fig. 2 Effect of temperature on reproductioin multiple of Aurelia sp.1 polyps |

|
| 图 3 温度对海月水母螅状体相对繁殖速度的影响 Fig. 3 Effect of temperature on relative reproduction velocity of Aurelia sp.1 polyps |
所有试验组螅状体均为一次性放入, 盐度7.5组中的螅状体1 h内全部死亡, 盐度10组中的螅状体1 d内全部死亡, 盐度12.5组中的螅状体3 d内全部死亡。其余盐度组(15—40) 中的螅状体成活率均为100%。试验期间, 各盐度试验组的螅状体均有无性繁殖现象, 由图 4和图 5可知, 盐度22.5组的螅状体相对繁殖速度最高, 随着盐度的递增或递减, 螅状体相对繁殖速度呈现下降趋势。盐度17.5—30范围内螅状体相对繁殖速度较高, 但方差分析结果表明, 盐度对螅状体繁殖率和相对繁殖速度的影响不显著(P>0.05) 。试验结果表明, 海月水母螅状体的适盐范围较广, 在盐度15—40范围均可存活并进行无性繁殖。

|
| 图 4 盐度对海月水母螅状体繁殖倍数的影响 Fig. 4 Effect of salinity on reproductioin multiple of Aurelia sp.1 polyps |

|
| 图 5 盐度对海月水母螅状体相对繁殖速度的影响 Fig. 5 Effect of salinity on relative reproduction velocity of Aurelia sp.1 polyps |
试验结束时, 800 lx组中的螅状体成活率为90%, 其余各组的成活率均为100%。螅状体繁殖倍数和相对繁殖速度随光照强度的增加而减少(图 6、图 7) , 0 lx试验组的螅状体繁殖倍数和相对繁殖速度最高, 分别为11.8%和3.98%。螅状体繁殖倍数和相对繁殖速度与光照强度呈线性相关, 螅状体繁殖倍数与光照强度的关系为y=-0.0151x+12.434 (n=5, r2=0.9299, P<0.01) , 相对繁殖速度与光照强度的关系为y=-0.0053x+4.398 (n=5, r2=0.8563, P<0.01) 。方差分析结果表明, 光照强度对螅状体繁殖倍数和相对繁殖速度的影响极显著(P<0.01) 。试验结果表明, 黑暗条件有利于海月水母经无性繁殖新螅状体, 增加光照强度可抑制海月水母螅状体种群数量繁殖。

|
| 图 6 光照强度对海月水母螅状体繁殖倍数的影响 Fig. 6 Effect of light intensity on reproductioin multiple of Aurelia sp.1 polyps |

|
| 图 7 光照强度对海月水母螅状体相对繁殖速度的影响 Fig. 7 Effect of light intensity on relative reproduction velocity of Aurelia sp.1 polyps |
试验期间, 各试验组的螅状体成活率均为100%, 且均有无性繁殖现象。由图 8和图 9可知, 螅状体繁殖倍数和相对繁殖速度随饵料量的增多呈现明显的上升趋势, 组间差异显著(P<0.05) 。亲代螅状体经60 d时间繁殖出的子代螅状体个数范围也随饵料量的减少而降低, 8.3 μg C/d组最高, 平均为22.8个, 范围为9—59个;4.2 μg C/d组平均为15.3个, 范围为5—23个, 1.0 μg C/d组平均为4.4个, 范围为2—10个, 0.5 μg C/d组平均为1.7个, 范围为1—3个。

|
| 图 8 饵料量对海月水母螅状体繁殖倍数的影响 Fig. 8 Effect of feeding on reproductioin multiple of Aurelia sp.1 polyps |

|
| 图 9 饵料量对海月水母螅状体相对繁殖速度的影响 Fig. 9 Effect of feeding on relative reproduction velocity of Aurelia sp.1 polyps |
海月水母因广泛的地理分布和较强的繁殖能力而使人们易于获得实验材料, 是目前全球被研究报道最多最详细的钵水母种类。20世纪初期, Mayer最先提出了海月水母具有12个亚种[21], 之后, Dawson等陆续对海月水母的亚种鉴定及地理分布进行了详细的研究, 认为每一亚种都有其特定的生态习性, 致使海月水母可跨越广泛的纬度和温度范围, 在全球不同地理生态环境水域中生存[9-10]。不同亚种不同地理种群的海月水母可能由于饵料和盐度的规律变化导致水母对某些基因片段的选择, 致使它们对自己特殊的环境条件有了适应性。
3.1 水温本试验结合黄渤海自然水域的水温范围, 将水温实验范围设置为0—25℃, 试验结果表明黄渤海海月水母螅状体在温度0—25℃下均可存活。Kroiher等观察到海月水母(Aurelia aurita)螅状体在4 ℃时解体死亡[22], 本试验中, 0 ℃组中的螅状体尽管不摄食, 但试验60d无死亡现象, 试验结束后当水温升高到5 ℃以上时, 螅状体恢复正常状态, 说明其耐低温的存活能力较强, 可以在黄渤海水域成功越冬。
海月水母的生活史十分复杂, 螅状体可通过多种无性繁殖方式进行种群数量的补充[23-25]。已有大量研究结果表明, 水温对海月水母螅状体无性繁殖有重要影响[26-27], 升温可加速螅状体繁殖速度[11, 27-29], Wang等的研究表明黄海海月水母(Aurelia sp.1) 螅状体的数量随水温的升高而增加[16, 18], 与本研究结果一致。相反, 美国华盛顿胡德运河的海月水母(Aurelia labiate)螅状体的出芽量随水温(7、10、15 ℃)的升高而减少[30];Liu等对台湾热带地区分布的海月水母(Aurelia aurita L.)的研究表明, 低温组(20 ℃)螅状体的出芽量比高温组(25 ℃和30℃)的多[31]。本温度试验中, 海月水母螅状体繁殖倍数和相对繁殖速度在5—12.5 ℃范围内的增加不明显, 当水温高于12.5 ℃后, 出现明显的上升趋势(图 2、图 3) , 这可能与螅状体的横裂生殖适宜温度有关, 已有研究结果表明海月水母螅状体发生横裂生殖的最适水温为13 ℃[16], 而春季横裂生殖期后的夏秋季高温期将是螅状体进行种群补充的高速繁殖期。黄渤海水域夏秋季的水温范围正处在15—25 ℃之间, 全球变暖导致的水温升高增加了螅状体高速繁殖期的时间, 致使水母暴发事件越来越频繁, 这也与许多全球变暖导致的温度升高可能使温带地区的水母种群数量增加的观点相印证[32-33]。本试验结果说明黄渤海海月水母螅状体与其他温带水域海月水母的生态习性一致, 属于典型温水性水母种类。
3.2 盐度本试验的盐度范围设置考虑到海湾河口盐度多变的情况, 可达到自然海域环境可能遭遇到的所有盐度范围, 而盐度15—40范围内40 d的存活率均为100%的试验结果说明了海月水母螅状体有广泛的适盐性, 完全可以适应盐度多变的河口等水域环境, 这应该也是海月水母可以在海湾、近岸和河口附近存活的主要原因之一。与众多研究结果一致, 盐度对螅状体繁殖的影响并不显著[11, 27], Willcox等研究表明塔斯马尼亚海月水母螅状体在盐度35中的出芽量最多, 认为高盐会加快螅状体种群生长速度[11], 本试验中, 盐度22.5组的螅状体相对繁殖速度最高, 低盐条件下的螅状体繁殖速度略快。
3.3 光照度海月水母螅状体常被发现于船舶、养殖浮阀底部和桥墩的侧立面等避光处[30, 34], 螅状体有向背阴处移动的行为[35]。相比温度和饵料量, 光照对螅状体无性繁殖的影响并不十分显著, 往往需要与其他因素发生交互作用[31, 36], 但光照也是调控水母螅状体种群数量不可缺少的环境因子之一[37-38]。不同地理生态习性的螅状体所适宜的光照区间截然不同, 台湾热带地区分布的海月水母(Aurelia aurita L.)螅状体的出芽量随光照度(375、56、0 lux)的减少而增加[31]。美国华盛顿胡德运河的海月水母(Aurelia labiata)螅状体的出芽量随每天光照时间(12、8、4 h)的减少而减少, 光照有利于加速螅状体发生横裂生殖、增加横裂生殖的次数并延长横裂生殖的时间[30]。以往有关光照条件对海月水母横裂生殖的影响研究较多[39], 多数研究认为强光抑制横裂生殖, 黑暗或弱光条件下横裂率较高[31, 40]。作者曾初步研究了光照强度对海月水母螅状体存活与生长的影响[35], 结果表明, 螅状体成活率随光照度的增加而减少, 本试验中各组螅状体的成活率较以往试验结果高, 原因可能与饵料供应量的多少有关, 螅状体的存活状态与多种环境因子有关, 在相同的光照强度条件下, 尤其是强光条件下, 饵料供应充足的螅状体其成活率可能要高于饵料欠缺的螅状体。
3.4 饵料量饵料对水母种群起着至关重要的调控作用[27, 41]。在水温适宜的环境条件下, 饵料充足的海月水母螅状体可迅速繁殖出大量螅状体, 以增加其种群数量。与其他大型水母种类相比, 海蜇、沙蜇和白色霞水母的螅状体进行足囊繁殖时一次通常仅能生成一个足囊, 适宜条件下最多同时生成2个或3个足囊, 与这些种类的螅状体繁殖速度相比, 海月水母螅状体的繁殖速度最快, 1个螅状体可同时生成多个螅状体, 一般为2—5个, 试验中观察到最多为8个, 子螅状体再繁殖的数量也多, 食物是螅状体重复进行无性繁殖的能量来源。Han等观察发现海月水母螅状体在26 ℃与饵料最多的交互组中的出芽量最多[28], 石岩等认为食物水平越高, 海月水母螅状体繁殖子体的能力越强[17], 众多研究均海月水母的螅状体繁殖速度随饵料量的增加而增加[15, 42], 本试验结果与之相符。本试验中, 无投喂组螅状体60 d的成活率为100%, 说明螅状体的饥饿耐受能力较强, 但螅状体在无饵料条件下可存活的时间与水温有直接关系, 高温加速新陈代谢会减少螅状体的存活时间。
4 小结实验结果表明, 海月水母螅状体在0—25 ℃范围内的存活率均为100%, 温度越高, 螅状体的个体繁殖速度越快;尽管盐度对螅状体繁殖速度的影响不显著, 但螅状体可适宜存活的盐度范围较广, 15—40范围内的成活率均为100%S, 其中低盐22.5条件下相对繁殖速度最高;黑暗条件有利于海月水母螅状体个体数量增加;螅状体相对繁殖速度随饵料量的增加而增加。研究结果说明海月水母螅状体对环境具有较强的适应性, 水温升高、饵料量增加可导致海月水母螅状体种群数量快速增加。近些年, 海月水母在黄渤海近岸水域爆发的事件越来越频繁, 未来仍有继续蔓延和加重的可能, 建议尽早采取有效措施加以控制和防范。
| [1] | Mills C E. Jellyfish blooms:are populations increasing globally in response to changing ocean conditions?. Hydrobiologia, 2001, 451(1): 55–68. |
| [2] | 孙松. 对黄、东海水母暴发机理的新认知. 海洋与湖沼, 2012, 43(3): 406–410. |
| [3] | Ki J S, Hwang D S, Shin K, Yoon W D, Lim D, Kang Y S, Lee Y, Lee J S. Recent moon jelly (Aurelia sp.1) blooms in Korean coastal waters suggest global expansion:examples inferred from mitochondrial COI and nuclear ITS 5.8S rDNA sequences. ICES Journal of Marine Science, 2008, 65: 443–452. DOI:10.1093/icesjms/fsn018 |
| [4] | Purcell J E, Uye S I, Lo W T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans:a review. Marine Ecology Progress Series, 2007, 350: 153–174. DOI:10.3354/meps07093 |
| [5] | Uye S I, Fujii N, Takeoka H. Unusual aggregations of the scyphomedusa Aurelia aurita in coastal waters along western Shikoku, Japan. Plankton Biology and Ecology, 2003, 50(1): 17–21. |
| [6] | Dong Z J, Liu D Y, Keesing J K. Jellyfish blooms in China:Dominant species, causes and consequences. Marine Pollution Bulletin, 2010, 60(7): 954–963. DOI:10.1016/j.marpolbul.2010.04.022 |
| [7] | 王彬, 董婧, 王文波, 李玉龙, 李轶平, 刘修泽, 付杰. 辽东湾北部近海大型水母数量分布和温度盐度特征. 海洋与湖沼, 2012, 43(3): 569–578. |
| [8] | 郑凤英, 陈四清, 倪佳. 海月水母的生物学特征及其爆发. 海洋科学进展, 2010, 28(1): 126–132. |
| [9] | Dawson M N, Hamner W M. Geographic variation and behavioral evolution in marine plankton:the case of Mastigias (Scyphozoa, Rhizostomeae). Marine Biology, 2003, 143(6): 1161–1174. DOI:10.1007/s00227-003-1155-z |
| [10] | Dawson M N, Jacobs D K. Molecular evidence for cryptic species of Aurelia aurita (Cnidaria, Scyphozoa). The Biological Bulletin, 2001, 200(1): 92–96. DOI:10.2307/1543089 |
| [11] | Willcox S, Moltschaniwskyj N A, Crawford C. Asexual reproduction in scyphistomae of Aurelia sp.:Effects of temperature and salinity in an experimental study. Journal of Experimental Marine Biology and Ecology, 2007, 353(1): 107–114. DOI:10.1016/j.jembe.2007.09.006 |
| [12] | Palmen E. Seasonal occurrence of ephyrae and subsequent instars of Aurelia aurita (L.) in the shallow waters of Tvärminne, S Finland. Archivum Societatis Zoologicae Botanicae Fennicae, 1953, 8:122-131. |
| [13] | Dong Z J, Liu Z Y, Liu D Y. Genetic characterization of the scyphozoan jellyfish Aurelia spp. in Chinese coastal waters using mitochondrial markers. Biochemical Systematics and Ecology, 2015, 60:15-23. |
| [14] | 李玉龙, 董婧, 孙明, 王彬, 王文波. 4种大型水母类ITS-5.8S rDNA序列分析及其在钵水母类系统分析中的应用. 中国水产科学, 2015, 22(6): 1278–1288. |
| [15] | 王彦涛, 孙松, 李超伦, 张芳. 温度、投饵频次对海月水母(Aurelia sp.1)水螅体生长和繁殖的影响. 海洋与湖沼, 2012, 43(5): 900–904. |
| [16] | Wang N, Li C L, Liang Y, Shi Y Q, Lu J L. Prey concentration and temperature effect on budding and strobilation of Aurelia sp. 1 polyps. Hydrobiologia, 2015, 754(1): 125–134. DOI:10.1007/s10750-014-1978-2 |
| [17] | 石岩, 米铁柱, 王建艳, 陈丽红, 王国善, 甄毓, 于志刚. 温度和食物水平对海月水母螅状体无性繁殖的影响. 生态学报, 2016, 36(3): 786–794. |
| [18] | Wang Y T, Zheng S, Sun S, Zhang F. Effect of temperature and food type on asexual reproduction in Aurelia sp. 1 polyps. Hydrobiologia, 2015, 754(1): 169–178. DOI:10.1007/s10750-014-2020-4 |
| [19] | Lucas C H. Reproduction and life history strategies of the common jellyfish, Aurelia aurita, in relation to its ambient environment. Hydrobiologia, 2001, 451(1): 229–246. |
| [20] | Uye S, Kayano Y. Predatory feeding of the planktonic copepod tortanus forcipatus on three different prey. Bulletin of the Plankton Society of Japan, 1994, 40(2): 173–176. |
| [21] | Mayer A G. Medusae of the World. III. The Scyphomedusae. Washington, DC:Carnegie Institute of Washington, 1910: 499–735. |
| [22] | Kroiher M, Siefker B, Berking S. Induction of segmentation in polyps of Aurelia aurita (Scyphozoa, Cnidaria) into medusae and formation of mirror-image medusa anlagen. The International Journal of Development Biology, 2000, 44(5): 485–490. |
| [23] | Berrill N J. Developmental analysis of Scyphomedusae. Biological Reviews, 1949, 24(4): 393–409. DOI:10.1111/brv.1949.24.issue-4 |
| [24] | 孙松, 于志刚, 李超伦, 黄邦钦, 庄志猛, 魏皓, 孙晓霞. 黄、东海水母暴发机理及其生态环境效应研究进展. 海洋与湖沼, 2012, 43(3): 401–405. |
| [25] | Vagelli A A. New observations on the asexual reproduction of Aurelia aurita (Cnidaria, Scyphozoa) with comments on its life cycle and adaptive significance. Invertebrate Zoology, 2007, 4: 111–127. |
| [26] | Holst S. Effects of climate warming on strobilation and ephyra production of North Sea scyphozoan jellyfish. Hydrobiologia, 2012, 690(1): 127–140. DOI:10.1007/s10750-012-1043-y |
| [27] | Thein H, Ikeda H, Uye S I. The potential role of podocysts in perpetuation of the common jellyfish Aurelia aurita s.l. (Cnidaria:Scyphozoa) in anthropogenically perturbed coastal waters. Hydrobiologia, 2012, 690(1):157-167. |
| [28] | Han C H, Uye S I. Combined effects of food supply and temperature on asexual reproduction and somatic growth of polyps of the common jellyfish Aurelia aurita s.l. Plankton and Benthos Research, 2010, 5(3): 98–105. DOI:10.3800/pbr.5.98 |
| [29] | Purcell J E, Atienza D, Fuentes V, Olariaga A, Tilves U, Colahan C, Gili J M. Temperature effects on asexual reproduction rates of scyphozoan species from the northwest Mediterranean Sea. Hydrobiologia, 2012, 690(1): 169–180. DOI:10.1007/s10750-012-1047-7 |
| [30] | Purcell J E. Environmental effects on asexual reproduction rates of the scyphozoan Aurelia labiata. Marine Ecology Progress Series, 2007, 348: 183–196. DOI:10.3354/meps07056 |
| [31] | Liu W C, Lo W T, Purcell J E, Chang H H. Effects of temperature and light intensity on asexual reproduction of the scyphozoan, Aurelia aurita (L.) in Taiwan. Hydrobiologia, 2009, 616(1):247-258. |
| [32] | Purcell J E. Climate effects on formation of jellyfish and ctenophore blooms:A review. Journal of the Marine Biological Association of the United Kingdom, 2005, 85(3): 461–476. DOI:10.1017/S0025315405011409 |
| [33] | 张芳, 孙松, 李超伦. 海洋水母类生态学研究进展. 自然科学进展, 2009, 19(2): 121–130. |
| [34] | Brewer R H. Larval settlement behaviour in the jellyfish Aurelia aurita (Linnaeus) (Scyphozoa:Semaeostomae). Estuaries, 1978, 1(2): 120–122. DOI:10.2307/1351601 |
| [35] | 孙明, 董婧, 付志璐, 李玉龙. 光照度对海月水母螅状体存活和生长的影响. 水产科学, 2012, 31(4): 212–215. |
| [36] | Kakinuma Y. An experimental study of the life cycle and organ differentiation of Aurelia aurita Lamarck. The Bulletin of the Marine Biological Station of Asamushi, 1975, 15: 101–113. |
| [37] | Dong J, Sun M, Purcell J E, Chai Y, Zhao Y, Wang A Y. Effect of salinity and light intensity on somatic growth and podocyst production in polyps of the giant jellyfish Nemopilema nomurai (Scyphozoa:Rhizostomeae). Hydrobiologia, 2015, 754(1): 75–83. DOI:10.1007/s10750-014-2087-y |
| [38] | 王建艳, 于志刚, 甄毓, 米铁柱, 姚庆祯, 王国善. 环境因子对海月水母生长发育影响的研究进展. 生态应用学报, 2012, 23(11): 3207–3217. |
| [39] | Custance D R N. Light as an inhibitor of strobilation in Aurelia aurita. Nature, 1964, 204(4964): 1219–1220. DOI:10.1038/2041219a0 |
| [40] | Ishii H, Shioi H. The effects of environmental light condition on strobilation in Aurelia aurita polyps. Sessile Organisms, 2003, 20: 51–54. DOI:10.4282/sosj.20.51 |
| [41] | Hamed A E, Khaled A A. Reproductive strategy of the jellyfish Aurelia aurita (Cnidaria Scyphomedusae) in the Suez Canal and its migration between the Red Sea and Mediterranean. Aquatic Ecosystem Health & Management, 2011, 14(3): 269–275. |
| [42] | Ishii H, Watanabe T. Experimental study of growth and asexual reproduction in Aurelia aurita polyps. Sessile Organisms, 2003, 20: 69–73. DOI:10.4282/sosj.20.69 |