 2017, Vol. 37
2017, Vol. 37文章信息
- 宋伦, 刘卫东, 吴景, 宋广军, 宋永刚, 孙明, 王年斌
- SONG Lun, LIU Weidong, WU Jing, SONG Guangjun, SONG Yonggang, SUN Ming, WANG Nianbin.
- 有害甲藻Stoeckeria algicida在辽东湾的时空分布
- Distribution of the toxic dinoflagellate Stoeckeria algicida in Liaodong Bay
- 生态学报. 2017, 37(4): 1339-1345
- Acta Ecologica Sinica. 2017, 37(4): 1339-1345
- http://dx.doi.org/10.5846/stxb201605241000
-
文章历史
- 收稿日期: 2016-05-24
- 修订日期: 2016-09-05

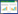




外来海洋生物入侵对生态系统稳定性的影响以及造成的经济损失, 已经引起科学界和社会公众的广泛关注。20世纪及21世纪初运输船舶压载水排放等导致外来海洋浮游生物广泛传播, 曾多次引发有毒有害赤潮, 对沿海养殖业、渔业资源、水产品质量、生态环境及人类健康都造成了巨大危害[1-2]。船舶压载水是海洋生物入侵的最重要途径, 其对海洋环境的侵害已被全球环境基金组织确认为与“陆源对海洋的污染”、“海洋生物资源掠夺性开发利用”、“海洋栖息环境的改变(破坏)”等共同构成危害海洋的四大威胁之一。因此, 世界海事组织、联合国发展计划署和全球环境基金组织联合提出了全球压载水控制与管理公约, 也开展了压载水的处置技术研究, 基于检测技术的限制, 主要是针对部分微型(粒径为2—20 μm)和小型浮游植物(粒径为20—200 μm)的检测。微微型浮游植物(粒径为0.22—2 μm)在海洋中多样性相当丰富, 也是褐潮暴发的主要致灾种, 由于微型藻类个体微小、形态学鉴定困难一度研究较为迟缓。
随着分子生物学的快速发展, 有关微型藻类多样性的研究得到了快速发展。目前, 已有学者对太平洋[3]、地中海[4]和印度洋[5]等各大海域的微型藻类分子多样性进行了研究, 相关的基因文库也在不断更新和扩充。我国在南海[6]、渤海[7]和北黄海[8]等海域对微型藻类的分子多样性研究也在逐渐开展。近年来, 新型海洋生态灾害频发, 由微型藻类引发的褐潮在我国渤海海域频繁暴发, 对贝类养殖危害较大[9]。我国目前报道的褐潮致灾种由抑食金球藻(Aureococcus anophagefferens)引发, 但最新研究发现, 在渤海可引发褐潮的微型藻类并非这一种[7]。近几年高通量测序技术的发展极大地推动了微型藻类的高效检测研究[10-12], 该技术具有简单快速、测序通量高、错误率低和成本低等特点, 为微型藻类的高效检测提供了新思路。
Stoeckeria algicida为甲藻纲胸甲球藻科, 有侵噬鱼类细胞杀鱼的能力, 可以导致鱼类成群死亡, 同时也会杀死其他海洋微藻, 2004年首次在韩国庆南马山海域被发现, 我国目前尚未发现有该种的报道[13]。辽东湾为我国最北端半封闭内海, 航运较为频繁, 加之富营养化严重, 发生外来有害微藻入侵的风险较高, 危害较大。本研究采用高通量测序平台, 结合生物信息学方法, 以18SrDNA的V4区为目标基因对辽东湾海域的微型和微微型浮游植物进行了多样性检测, 已发现外来有害微藻Stoeckeria algicida的扩散迹象, 因此亟需了解其生态分布状况。
1 材料与方法 1.1 样品采集为了解外来有害微藻Stoeckeria algicida的分布及扩散情况, 在辽东湾海域网格化设置16个站位(图 1) , 分别于2014年5月(春)、8月(夏)、10月(秋)、12月(冬)采集表层海水1L, 首先经20μm微孔滤膜过滤去除小型及大型浮游生物, 然后经0.22μm微孔滤膜收集微型和微微型浮游生物, 最后将滤膜转移至1.5mL无菌离心管中, 置于-20℃或-80℃冷冻保存、运输。

|
| 图 1 采样站位示意 Fig. 1 Sampling station locations |
采用CTAB法提取微型/微微型浮游植物宏基因组, 将0.22 μm滤膜剪碎于1.5 mL离心管中, 加入500 μL CTAB裂解液(2%CTAB;100 mmol/L Tris-Cl pH 8.0;1.4 mmol/L NaCl;10 mmol/L EDTA)和1 μL β-巯基乙醇, 5—10 μL蛋白酶K, 55℃裂解 1—1.5 h。短暂离心, 取出液体于新的离心管中, 用酚氯仿抽提2 次后, 取上清, 加入两倍体积预冷的无水乙醇, 沉淀2—3 h, 保留沉淀, 使用75%乙醇清洗沉淀, 得到微藻宏基因组DNA, 1%琼脂糖凝胶电泳检测并利用紫外分光光度计测定DNA浓度及纯度, -20℃冰箱保存备用。
1.2.2 18S rDNA可变区V4的PCR扩增本研究应用的引物为自行开发的微型/微微型浮游植物18S rDNA的V4区基因扩增引物对V4(F/R)。上游引物为V4-F序列5′-GCGGTAATTCCAGCTCCAATA-3′, 下游引物V4-R序列为5′-GATCCCCHWACTTTCGTTCTTGA-3′。将引物连接适当的接头送往上海生工生物公司进行合成。PCR反应体系为50 μL, 包括PCR Buffer 5 μL、dNTP Mixture 8μL、上下游引物(10 μmol/L)各2 μL、模板DNA 2 μL、Taq DNA聚合酶2.5 U, 加适量灭菌水。扩增反应均在PE 9700型PCR仪(美国PE公司)上完成, 反应条件:94℃预变性3 min;94℃变性30 s, 58 ℃退火45 s, 72℃延伸45 s, 共33个循环;72℃延伸5 min。1%琼脂糖凝胶电泳检测PCR产物, 将检测合格产物交由诺和致源生物信息科技有限公司, 使用NEB Next UltraTM DNA Library Prep Kit for Illumina(New England Biolabs)建库试剂盒进行文库的构建, 构建好的文库经过Qubit定量和文库检测, 合格后使用Hiseq2500 PE250进行上机测序。
1.2.3 数据分析测序得到的原始数据使用FLASH软件进行拼接, 参照Qiime软件质量控制流程将拼接后的序列经过截取、过滤得到有效数据。利用Uparse软件对有效数据进行OTUs(Operational Taxonomic Units)聚类和物种分类, 采用RDP Classifier方法与Silva数据库对OUTs代表序列进行物种注释, 同时对物种注释在各个分类水平上进行群落结构的统计分析。镜检同期水样中出现的所有浮游植物, 筛选各站位出现频度最高的种类角毛藻属(Chaetoceros sp.)作为基准密度[14], 通过OTUs比例, 计算Stoeckeria algicida密度, 相关公式如下:
(1) 利用OTUs比例测算Stoeckeria algicida密度:
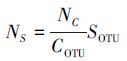
|
式中,NS为该站位Stoeckeria algicida密度(个/L), NC为同一站位水样中所有角毛藻密度(个/L, 镜检获得), COTU为该站位水样中所有角毛藻的OTUs数, SOTU为该站位Stoeckeria algicida的OTUs数。
(2) 相对多度(RA)计算Stoeckeria algicida的季节比例:
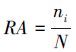
|
式中, ni为i种的密度(个/L), N为统计单元的总密度(个/L)。
(3) 物种优势度(Y)表示微型/微微型浮游植物群落中某一物种在其中所占的优势程度:
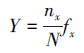
|
式中, nx为第x种微型/微微型浮游植物种类的OTUs数, N为OTUs总数, fx为第x种微型/微微型浮游植物种类在各样品中出现的频率。
2 结果 2.1 分子鉴定引物优化为增加微型/微微型浮游植物检测的高效性, 对引物的进行了筛选优化。本文针对目前已发现的微型/微微型浮游植物并结合浮游植物历史调查结果, 设计了比较适合我国黄渤海海域的微型浮游植物分子鉴定特异引物。针对浮游植物18S rDNA V4区基因设计了引物对V4(F/R)(5′-GCGGTAATTCCAGCTCCAATA-3′, 5′-GATCCCCHWACTTTCGTTCTTGA-3′), 针对V9区基因选择通用引物对V9(F/R)(5′-CCCTGCCHTTTGT ACACAC-3′, 5′-CCTTCYGCAGGTTCACCTAC-3′), 并选择Stoeck[15]等人设计的真核生物18S rDNA V4区扩增引物对C4(F/R)(5′-CCAGCASCYGCGGTAATTCC-3′, 5′-ACTTTCGTTCTTGATYRA-3′)进行了比较分析。
V4(F/R)引物对的设计是从NCBI数据库下载179(76条微型/微微型)条藻类18S rDNA序列(包含11个门、23个纲、179种), 利用软件MEGA4进行多重序列比对, 找到对应的V4区, 同时结合测序技术要求进行引物设计, 片段长度为300—450bp, 并使用DNAMAN、Oligo Calc等软件对引物的基本参数进行检测和评估。统计结果显示, 引物V4(F/R)对真菌、后生动物、领鞭毛虫目类群的扩增特异性要低于V9(F/R)和C4(F/R), 而对真核藻类的扩增特异性均高于V9(F/R), 扩增红藻门、定鞭藻纲的特异性与C4(F/R)虽有差异, 但在其他真核藻类类群中较有优势。
为了比较3对引物在实际样品中对真核藻类扩增特异性的差异, 对2014年5—8月采集的8、11、12号环境样品分别进行了引物扩增, 采用Illumina HiSeq2500测序平台PE250测序, 解析出在种的水平下每对引物对应的平均微型/微微型浮游植物OTUs数。结果显示, 3对引物V4(F/R)、 V9(F/R)、C4(F/R)3个站位鉴定的微型/微微型浮游植物OTUs数分别为68、73、92;37、46、44;33、59、81, 平均为78、42、58。结果显示, 引物对V4(F/R)在微型/微微型浮游植物种类数鉴定方面优于其他两对引物, 故采用引物对V4(F/R)研究辽东湾海域微型/微微型浮游植物的群落多样性。
2.2 Stoeckeria algicida的种属确定高通量测序获得Stoeckeria algicida 部分18S rDNA序列为375bp, 通过NCBI Blast比对, 其在Stoeckeria algicida18S rDNA(AJ841809.1) 基因序列中对应的位置为595—969bp, 与基因库中Stoeckeria algicida的18S rDNA序列相似度为100%, E值为0。采用邻接法, 选择甲藻门中部分藻类的18S rDNA序列构建系统进化树(图 2, 节点上的数值表示邻接法的可靠性)。建树结果显示, 高通量测序获得的Stoeckeria algicida 18S V4区序列与Stoeckeria algicida(HG005133) 聚在一个分支, 其支持率为100%, 确定该种为Stoeckeria algicida。

|
| 图 2 Stoeckeria algicida系统进化树 Fig. 2 Stoeckeria algicida phylogenetic tree |
辽东湾春季所有站位均未检出Stoeckeria algicida, 夏季密度分布在(0.023—2.753) ×103个/L, 平均为1.388×103个/L, 最高值出现在1号站, 最低值出现在14号站, 高值区主要分布在辽东湾的东西两侧(1、4、7、9、10、12、13、16号站位密度均在103个/L以上), 辽东湾中部海域密度相对较少;秋季密度分布在(0.011—2.488) ×103个/L, 平均为1.249×103个/L, 最高值出现在9号站, 最低值出现在12号站, 高值区主要分布在辽东湾西北侧的1号(1.250×103个/L)和中东侧9号(2.488×103个/L)站位, 其他区域密度相对较少;冬季密度分布在(0.002—0.073) ×103个/L, 平均为0.037×103个/L, 最高值出现在3号站, 最低值出现在14号站, 密度量相对较少, 各站位差异不大, 分布较为均匀(图 3) 。

|
| 图 3 不同季节Stoeckeria algicida的相对密度 Fig. 3 Relative Stoeckeria algicida density in different seasons |
夏季Stoeckeria algicida优势度在(0.03—0.41) %, 平均为0.22%, 优势度最高在9号站位, 最低在16号站位(6、8、9、15号站位优势度超过0.2%);秋季优势度在(0.03—0.54) %, 平均为0.28%, 优势度最高在16号站位, 最低在4号站位(6、9、11—16号站位优势度超过0.2%);冬季优势度在(0.02—0.34) %, 平均为0.18%, 优势度最高在16号站位, 最低在4号站位(7、9、11—16号站位优势度超过0.2%)(图 4) 。

|
| 图 4 不同季节Stoeckeria algicida的优势度 Fig. 4 Stoeckeria algicida dominance in different seasons |
目前有关Stoeckeria algicida的研究报道较少, 成果主要来自韩国首尔国立大学海洋研究所赤潮研究中心[16-17], 2004年首次在韩国庆南马山海域发现胸甲球藻科一个新种S. algicida, 该种曾引发两次大规模赤潮, 给渔民造成了1亿多韩元的经济损失, 暴发密度可达2×107 个/L, 有侵噬鱼类细胞杀鱼的能力, 可以导致鱼类成群死亡, 同时也会杀死其他海洋微藻。研究组以世界海洋生态学家、美国马里兰大学教授戴安·斯迪克(音译)的名字结合“杀死其他赤潮生物”之意的拉丁语“algicida”将此生物的名字命名为斯迪克里亚·阿尔兹西达(音译)。
Jeong等对异养型甲藻S. algicida的形态学特征进行了描述, 并从人工培养的藻细胞中获得核糖体小亚基rDNA序列。该藻细胞呈椭圆形, 长(7.3—15.9 μm, 平均11.6 μm)大于宽(2.7—12.2 μm, 平均7.3 μm)。核糖体小亚基rDNA序列的GenBank登录号为AJ841809。序列比对结果显示, S. algicida的核糖体小亚基rDNA序列与牧羊杖地区的一种甲藻差异3%, 与Cryptoperidiniopsoid sp.、brodyi、Pfiesteria sp.(有害费氏藻)差异4%。采用最大似然法构建S. algicida的核糖体小亚基rDNA序列的系统进化树, S. algicida与有害费氏藻及近缘种、以及牧羊杖地区的一种甲藻亲缘关系最为接近, 却又处在不同的分支上。基于形态学观察和谱系分析, 认为S. algicida是一个新种[15]。
为研究S. algicida对其他赤潮生物的影响, Jeong等在2004年5—6月赤潮期间监测了韩国马上海湾赤潮异弯藻(Heterosigma akashiwo)和S. algicida的密度变化。同时在实验室研究S. algicida摄食赤潮异弯藻情况, 检测其生长率和捕食率。结果发现赤潮异弯藻最大密度出现在第2次赤潮期间(58400—99200 cells/mL), S. algicida为1130—17400 cells/mL, 其密度的高峰期相对赤潮异弯藻要推迟1—2 d。S. algicida捕食赤潮异弯藻是先通过一个柄状组织将其固定, 然后通过细丝拖拽, 在赤潮异弯藻的密度达到约3500 cells/mL饱和浓度之前, S. algicida的比生长速率随着赤潮异弯藻密度的增加而快速升高, 其最大比生长速率为1.63/d, 当赤潮异弯藻的阈值浓度为1.9 ng·C/mL(19 cells/mL)时, 净生长率为0, 对赤潮异弯藻的最大摄食率和清除率分别为0.75 ng C/grazer·mL(7.5 cells /grazer·mL)和3.7 μL/h。S. algicida对赤潮异弯藻的捕食系数达到0.142 cells /min(即每分钟清除赤潮异弯藻0.13%), 目前的研究结果显示, S. algicida的摄食对赤潮异弯藻的密度影响较大[16]。
Stoeckeria algicida为甲藻纲胸甲球藻科, 有侵噬鱼类细胞杀鱼的能力, 可以导致鱼类成群死亡, 同时也会杀死其他海洋微藻, 2004年首次在韩国庆南马山海域被发现。由于该种研究较少, 致灾机理尚不清楚, 但其与费氏藻属(Pfiesteria sp.)同属一科, 费氏藻产生的毒素能使半咸水鱼类很快死亡, 表现为皮肤溃烂, 失去抵抗力, 有研究表明其毒素分脂溶性、水溶性两种, 脂溶性表现为皮肤伤害作用, 水溶性表现为神经伤害作用, 脂溶性为一种塑料增塑剂, 水溶性成分是含铜和铁元素的糖苷类物质[18]。2013—2014年在大连长兴岛海域曾发现大量死鱼, 检测水样中Stoeckeria algicida的密度在106 个/L左右, 接近赤潮警戒标准(107 个/L)[19]。
通过对辽东湾四季Stoeckeria algicida的检测, 春季未检出, 冬季密度急剧下降, 说明温度是影响该藻繁殖的主要因素。夏、秋季整个海域都有检出, 虽然Stoeckeria algicida在整个环境样品中优势度不太明显, 但其密度较高, 最高达2.753×103个/L, 夏季主要分布在辽东湾东西两岸, 致灾风险较高, 应引起足够重视。
Stoeckeria algicida被韩国定为外来物种, 在我国海域也是首次报道, 其危害后果严峻, 必须加强监测监管。从目前辽东湾海域的Stoeckeria algicida密度分布来看, 该种已成为归化种, 虽未发生赤潮记录, 但防患于未然, 未来应从以下3方面入手:①加快研究该藻的产毒机制、生态危害、暴发风险、防御措施;②重点监测各港口海域有毒微型/微微型藻类的种群和分布, 补充外来入侵微藻数据库;③加强对养殖区产毒微藻的监测和管理, 建立贝毒快速预警技术, 减少对渔业生态系统的危害[20-23]。
| [1] | 宋伦, 毕相东. 渤海海洋生态灾害及应急处置. 沈阳: 辽宁科学技术出版社, 2015: 175–210. |
| [2] | 梁玉波, 王斌. 中国外来海洋生物及其影响. 生物多样性, 2001, 9(4): 458–465. |
| [3] | Worden A Z. Picoeukaryote diversity in coastal waters of the Pacific Ocean. Aquatic Microbial Ecology, 2006, 43(2): 165–175. |
| [4] | Marie D, Zhu F, Balagué V, Ras J, Vaulot D. Eukaryotic picoplankton communities of the Mediterranean Sea in summer assessed by molecular approaches (DGGE, TTGE, QPCR). FEMS Microbiology Ecology, 2006, 55(3): 403–415. DOI:10.1111/fem.2006.55.issue-3 |
| [5] | Not F, Latasa M, Scharek R, Viprey M, Karleskind P, Balagué V, Ontoria-Oviedo I, Cumino A, Goetze E, Vaulot D, Massana R. Protistan assemblages across the Indian Ocean, with a specific emphasis on the picoeukaryotes. Deep Sea Research Part I:Oceanographic Research Papers, 2008, 55(11): 1456–1473. DOI:10.1016/j.dsr.2008.06.007 |
| [6] | 李绿砚. 中国南海微型与超微型真核浮游生物的分子生态学研究[D]. 广州:中山大学, 2008. |
| [7] | 于杰. 浮游生物多样性高效检测技术的建立及其在渤海褐潮研究中的应用[D]. 青岛:中国海洋大学, 2014. |
| [8] | 江雪娇. 北黄海微微型浮游植物的丰度及微微型真核浮游生物分子多样性研究[D]. 青岛:中国海洋大学, 2009. |
| [9] | Gobler C J, Lonsdale D J, Boyer G L. A review of the causes, effects, and potential management of harmful brown tide blooms caused by Aureococcus anophagefferens (Hargraves et sieburth). Estuaries, 2005, 28(5): 726–749. DOI:10.1007/BF02732911 |
| [10] | 王丽平, 南炳旭, 扈培龙. 秦皇岛褐潮暴发敏感海域细菌种群特征. 环境科学研究, 2015, 28(6): 899–906. |
| [11] | Amaral-Zettler L, Artigas L F, Baross J, Loka Bharathi P.A., Boetius A, Chandramohan D, Herndl G, Kogure K, Neal P, Pedrós-Alió C, Ramette A, Schouten S, Stal L, Thessen A, de Leeuw J, Sogin M. A global census of marine microbes. In:McLntyre A D, ed. Life in the World's Oceans:Diversity, Distribution and Abundance. Oxford:Blackwell Publishing Ltd, 2010:223-245. |
| [12] | Howad E C, Henriksen J R, Buchan A, Reisch C R, Bürgmann H, Welsh R, Ye W Y, González J M, Mace K, Joye S B, Kiene R P, Whitman W B, Moran M A. Bacterial taxa that limit sulfur flux from the ocean. Science, 2006, 314(5799): 649–652. DOI:10.1126/science.1130657 |
| [13] | 刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 2008: 301–870. |
| [14] | 辽宁省质量技术监督局. DB21/T 2427-2015海洋褐潮监测技术规程. 辽宁省地方标准, 2015. |
| [15] | Stoeck T, Bass D, Nebel M, Christen R, Jones M D M, Breiner H W, Richards T A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Molecular Ecology, 2010, 19(S1): 21–31. |
| [16] | Jeong H J, Kim J S, Park J Y, Kim J H, Kim S, Lee I, Lee S H, Ha J H, Yih W H. Stoeckeria algicida n. gen., n. sp. (Dinophyceae) from the Coastal Waters off Southern Korea:Morphology and Small Subunit Ribosomal DNA Gene Sequence. The Journal of Eukaryotic Microbiology, 2005, 52(4):382-390. |
| [17] | Jeong H J, Kim J S, Kim J H, Kim S T, Seong K A, Kim T H, Song J Y, Kim S K. Feeding and grazing impact of the newly described heterotrophic dinoflagellate Stoeckeria algicida on the harmful alga Heterosigma akashiwo. Marine Ecology Progress Series, 2005, 295: 69–78. DOI:10.3354/meps295069 |
| [18] | 林旭吟, 杨军霞, 李扬, 吴云辉. 有害费氏藻(Pfiesteria)的研究进展. 福建水产, 2007(4): 54–60. |
| [19] | 中华人民共和国国家海洋局. HY/T 069-2005赤潮监测技术规程. 北京: 中国标准出版社, 2005. |
| [20] | 孙军, 刘东艳, 徐俊, 陈凯彪. 1999年春季渤海中部及其邻近海域的网采浮游植物群落. 生态学报, 2004, 24(9): 2003–2016. |
| [21] | 蔡琨, 秦春燕, 李继影, 张咏, 牛志春, 李旭文. 基于浮游植物生物完整性指数的湖泊生态系统评价——以2012年冬季太湖为例. 生态学报, 2016, 36(5): 1431–1441. |
| [22] | 吕永龙, 苑晶晶, 李奇锋, 张悦清, 吕笑天, 苏超. 陆源人类活动对近海生态系统的影响. 生态学报, 2016, 36(5): 1183–1191. |
| [23] | 姜冰, 宋伦, 时明明, 宋广军, 王影. 辽宁近海外来赤潮海洋微藻入侵现状. 水产科学, 2015, 34(12): 795–800. |