 2017, Vol. 37
2017, Vol. 37文章信息
- 葛晓改, 曾立雄, 肖文发, 黄志霖, 周本智
- GE Xiaogai, ZENG Lixiong, XIAO Wenfa, HUANG Zhilin, ZHOU Benzhi.
- 模拟N沉降下不同林龄马尾松林凋落叶分解-土壤C、N化学计量特征
- Relationship between leaf litter decomposition and soil C, N stoichiometry in different-aged Pinus massioniana stands exposed to simulated nitrogen deposition
- 生态学报. 2017, 37(4): 1147-1158
- Acta Ecologica Sinica. 2017, 37(4): 1147-1158
- http://dx.doi.org/10.5846/stxb201604280808
-
文章历史
- 收稿日期: 2016-04-28
- 修订日期: 2016-07-11





2. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091;
3. 南京林业大学南方现代林业协同创新中心, 南京 210037;
4. 国家林业局秭归森林生态系统定位观测研究站, 秭归 443000
2. State Forestry Administration Key Laboratory of Forest Ecology and Environment; Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China;
3. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China;
4. Zigui Forest Ecosystem Research Station, State Forestry Administration, Zigui 443000, China
近20年来,由于我国经济的高速发展,尤其东部地区存在着高N沉降问题[1]。我国近年Nr排放量激增,并由此导致N沉降量的显著增加,尤其是我国东部和南部的N沉降量普遍高于3 g m-2 a-1,与欧美地区的最高值相当[1-2]。人类活动已显著增加了活性N的生成和沉降速率,增加的N对陆地生态系统C循环过程产生了一系列影响。有研究表明中国1981-2010年期间N沉降增加了0.058 g N m-2 a-1,N沉降通过增加地上生产力或改变凋落物质量 (如降低凋落物C/N比) 诱发中国森林土壤总碳库增加4.78%,大部分积累在植被中[3],且增加的树木生长通过增加植物凋落物和根系输入促进土壤有机质积累[4];较低的N素沉降能促进森林生长,但当N沉降超过生态系统可以利用的限度时,就会发生N饱和,进而导致土壤酸化、森林生产力下降、生物多样性降低、延缓凋落物分解等[5],从而影响森林生态系统各种营养元素的循环。森林凋落物分解是陆地生态系统土壤有机质、有机养分矿化和碳平衡形成的重要环节[6],在促进森林生态系统正常物质循环、维持土壤肥力[7]、保证植物再生长养分可利用中起着重要作用[8]。因此,研究N沉降下森林凋落物分解过程中C-N相互作用对确定N输入对森林生态系统C循环的影响很重要[3]。
N沉降主要通过两方面影响凋落物分解。一方面,大气N沉降改变了森林群落的组成与结构,改变凋落物的外部环境,变化后的森林凋落物化学组成间接影响森林凋落物分解;另一方面,N沉降增加了凋落物分解过程中外源N含量,影响凋落物分解的化学过程,改变了凋落物分解速率[9]。我国模拟大气N沉降对凋落物分解的相关研究主要是在鼎湖山[10-11]、长白山[12]、四川雅安[13-14]、南昌[15]等地区针对季风常绿阔叶林 (monsoon evergreen broad-leaved forest)、针阔混交林 (conifer-broadleaf forest)、华西雨屏区慈竹 (Neosinocalamnus affinis)、亮叶桦 (Betula luminifera)、湿地松 (Pinus elliottii) 等,主要侧重凋落叶分解过程、养分释放、森林生物多样性[16]、土壤酶活性[17]、土壤有机碳和养分[18]等方面。目前,有关N沉降对不同树种凋落叶分解的研究较多,对不同林龄凋落叶分解中C、N相互作用关注较少[19],且凋落叶分解对N沉降的响应尚存争议,外加N源对针叶凋落物的分解有促进作用[10-11]、抑制[14]或影响不明显,因此,N沉降对凋落叶分解的影响仍需长期深入研究。
三峡库区地处中亚热带北缘,气候和植被处于过渡状态,是我国生态环境保护的重点地区;该地区复杂的地形变化,使区域内的生态系统十分敏感和脆弱。马尾松 (Pinus massoniana) 是我国亚热带地区主要针叶树种,因其适应性强、耐干旱与瘠薄,被广泛用于荒山绿化,是三峡库区典型的造林树种之一[20]。研究表明林木龄级影响马尾松林凋落物产量、质量、林地凋落物养分积累量等[21],且树木的不同生长发育阶段对土壤养分的需求不同,如年幼的树木凋落物产量一般随着土壤N的有效性提高而增加;马尾松林凋落叶C/N比较大 (50<凋落物C/N比<65),是凋落叶分解初期的主要限制因子之一,开展不同林龄马尾松林在外源N输入变化下凋落叶分解-土壤C、N及其化学计量响应研究对土壤养分供应力和生态系统养分循环有重要意义。本研究以三峡库区不同林龄马尾松林为研究对象,通过18个月的模拟N沉降和原位分解试验,探讨N沉降对不同林龄马尾松林凋落叶分解速率、凋落叶分解-土壤C、N及其化学计量的影响,对全面评价N沉降对森林生态系统结构和功能的影响具有重要意义,为理解N沉降对三峡库区森林经营和管理提供参考。
1 材料和方法 1.1 研究区概况研究区位于湖北省宜昌市秭归县 (30°38′14″-31°11′31″N,110°00′14″-111°18′41″E),海拔40-2057 m,该地区属于亚热带大陆性季风气候,年均气温17-19 ℃,极端最高温可达44.2 ℃(1998年),极端最低气温-2.5 ℃(1989年)。库区雨量充足,降水集中在4-9月,年降水量1000-1250 mm,空气相对湿度较大,可达60%-80%。秭归县海拔500 m以下以茶园、板栗、柑橘和农田为主,海拔500 m以上主要为灌木、马尾松林、杉木 (Cunninghamia lanceolata) 和松栎混交林等[21]。
本研究以秭归县茅坪镇境内20年生、30年生、46年生马尾松林为研究对象,林下灌木多为油茶 (Camellia oleifera)、宜昌荚蒾 (Viburnum ichangense)、檵木 (Loropetalum chinensis)、毛黄栌 (Cotinus coggygria) 等为优势种,草本多为铁芒萁 (Dicranopteris gigantea)、芒 (Miscanthus sinensis)、稗草 (Echinochloa crusgalli)、腹水草 (Veronicastrum axillare) 等,层间植物有爬藤榕 (Ficus martinii)、菝葜 (Smilax china) 等[20]。马尾松林分均是20世纪60年代初、70年代末飞播林,由于30年生马尾松林坡度、海拔及其土壤环境致使土壤养分含量较低,研究样地基本概况见表 1。
| 林分 Stands | 20年生 20-year-old stand | 30年生 30-year-old stand | 46年生 46-year-old stand |
| 海拔Elevation/m | 964 | 357 | 987 |
| 年龄Age/a | 20 | 30 | 46 |
| 胸径DBH/cm | 16.1±1.15 | 25.3±0.93 | 33.1±1.09 |
| 坡度Slope/(°) | 20 | 25 | 15 |
| 坡向Aspect | 阳坡 | 半阴坡 | 阳坡 |
| 郁闭度Canopy density | 0.60±0.03 | 0.95±0.01 | 0.85±0.02 |
| 土壤厚度Soil depth/cm | 70-100 | 60-80 | 70-100 |
| 凋落物现存量Litter standing crop/(t/hm2) | 9.35±5.14 | 9.26±2.97 | 14.05±6.40 |
| 凋落物年产量Annual litterfall/(t hm-2 a-1) | 3.38±0.72 | 4.69±0.20 | 5.60±0.23 |
| 凋落物层厚度Litter layer depth/cm | 4.03±1.59 | 5.77±0.85 | 6.34±1.99 |
| 凋落物木质素Litter lignin/% | 34.27±1.31 | 34.31±0.42 | 34.24±0.16 |
| 凋落物纤维素Litter cellulose/% | 8.68±0.12 | 8.17±1.65 | 6.7±0.51 |
| 土壤有机质Soil organic matter/(g/kg) | 73.10±16.66 | 20.96±10.89 | 82.77±10.17 |
| 土壤总氮Total nitrogen/(g/kg) | 3.27±0.87 | 1.43±0.40 | 4.22±0.70 |
在不同林龄马尾松林中各设置3个20 m×30 m的样地,每个样地中随机设置4个3 m×3 m的小样方,即每林龄马尾松林分别设置12个小样方,用于N沉降处理。为防止相互干扰,小样方间至少相隔10 m。每个小样方放置凋落叶分解袋前清理凋落物,放置凋落叶分解袋后每个月底进行新近凋落物清理。根据马尾松凋落叶基质质量 (50<基质质量C/N比<65) 并参考国际上同类研究氮处理的强度和频度,试验分4个处理,分别为对照 (CK)、低氮 (LN,5 g m-2 a-1)、中氮 (MN,10 g m-2 a-1) 和高氮 (HN,15 g m-2 a-1),每个马尾松林分每个处理3次重复。2010年8月将拟在每个样方所施的NH4NO3溶解在3 L去离子水中 (全年所增加的水量相当于新增降水1.0 mm) 后,用1.5 L的小喷壶来回均匀喷洒。对照样方则喷洒同样多的水,以尽量减少不同处理间的差异。除了施氮处理外,其它处理措施均保持一致。
凋落叶分解研究采用分解袋法。2010年5-7月,在3种马尾松人工林地中分别收集新近自然凋落、上层未分解的针叶,同一样地收集的凋落叶充分混合,风干后,称取凋落叶15 g,分别装入大小为20 cm×20 cm、孔径为1 mm的分解袋中。2010年7月底将凋落叶分解袋分别置于除去地表凋落物层的3个马尾松林地土壤表层,让凋落叶自然分解。3林龄马尾松林中各放置288袋 (6次×12个样方×4袋/次/样方),3种林分共864袋。同时,取3个林龄马尾松凋落叶各15 g,重复3次,在70 ℃下烘干至恒重,算出风干与烘干转换系数。自放置日期起,每3个月每种马尾松林随机取回48袋 (12个样方×4袋/次/样方),共6次取样。取回凋落叶分解袋后,清除侵入的根系、泥沙,烘至恒量,称量干质量并计算干质量残留率、分解速率和凋落叶C、N动态。
2010年8月在每林龄马尾松林选定的样地中分别用直径为5 cm的土钻沿对角线采表层 (0-5 cm) 土壤,约0.5 kg,每土样3次重复,进行背景值测定。于分解的90 d (2010年11月)、180 d (2011年3月)、270 d (2011年6月) 和360 d (2011年9月) 用直径为5 cm的土钻分别取凋落叶分解袋下面的表层土壤 (0-5 cm) 约0.5 kg,共144个土样 (4处理×3林龄×3重复×4次),带回实验室。将背景值和N沉降处理下的土壤样品去掉石砾、动植物、根系后,混匀,风干,粉碎后过1 mm和0.25 mm筛,装入自封袋待测土壤有机碳 (SOC,soil organic carbon) 和全氮含量 (TN,soil total nitrogen)。
1.3 凋落叶和土壤样品测定凋落叶化学分析:将收集的样品烘干磨碎,过0.2mm筛,用重铬酸钾氧化-外加热法测定全C (LC);用凯氏定氮法测定全N (意大利,UK152)(LN);起始凋落叶木质素和纤维素分别用酸洗涤法和中性洗涤法测定。
土壤化学性质分析:土壤养分指标采用《中华人民共和国林业行业标准方法》[22]进行测定。采用重铬酸钾外加热法测定SOC;重铬酸钾-硫酸消化法测定TN;同时称取20 g新鲜土壤于105 ℃烘12 h,测定土壤含水量。
1.4 数据处理凋落叶分解干重剩余率计算公式采用:
式中, R为凋落叶分解干重剩余率 (%);k为凋落叶风干样品与烘干样品转换系数;Mt为凋落叶t时间烘干样品重量;M0为凋落叶起始风干样品重量。
凋落叶分解速率计算公式采用: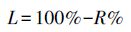
式中, L为凋落叶分解速率 (%);R为凋落叶分解干重剩余率 (%)。
利用Excel 2007和SPSS 16.0软件进行统计分析和作图,模拟N沉降下凋落叶和土壤各变量间的显著性检验采用单因子方差分析 (ANOVA) 和最小显著差异法 (LSD),显著性水平设定为α=0.05;凋落叶分解各指标与土壤养分的影响用Canoco 4.5的典范对应分析 (CCA) 模块和SPSS相关分析。
2 结果 2.1 模拟N沉降下不同林龄马尾松凋落叶分解模拟N沉降处理下凋落叶分解速率模式类似,均是先慢后快 (图 1)。20年生马尾松林凋落叶分解0-90 d时,N沉降抑制分解,360-540 d促进分解;分解540 d后,干重剩余率是高氮 (63.16±5.26)%<低氮 (64.25±4.34)%<中氮 (64.60±4.01)%<对照 (65.29±1.14)%,低、中、高氮处理凋落叶干重剩余率分别低于对照1.04%、0.69%和2.13%,且分解180 d时对照与中氮处理差异显著 (P<0.05),分解360 d中氮处理明显低于其它几个处理 (P<0.05)。30年生林分N沉降处理促进凋落叶分解,分解540 d后,干重剩余率是中氮 (58.70±2.11%)<高氮 (60.44±0.80)%<低氮 (60.67±4.40)%<对照 (60.89±2.79)%,低、中、高氮处理凋落叶干重剩余率分别低于对照0.35%、3.58%和0.73%,且分解90 d时高氮处理明显促进分解 (P<0.05)。46年生林分在N沉降下,分解540 d后,仅低氮处理促进分解,干重剩余率是低氮 (62.74±4.99)%<对照 (64.38±2.79)%<中氮 (64.63±3.40)%<高氮 (64.94±1.70)%,低氮处理凋落叶干重剩余率低于对照2.55%,中氮和高氮分别高于对照0.38%和0.86%,分解180 d时中氮处理明显低于其它处理 (P<0.05)。

|
| 图 1 不同林龄马尾松林凋落叶分解干重剩余率对N沉降的响应 Fig. 1 The response of litter leaf decomposition dry mass remaining rate to nitrogen deposition in different-aged P. massoniana stands 不同大写字母表示同一处理不同分解时间差异显著,不同小写字母表示同一分解时间不同处理间差异显著 |
模拟N沉降处理下,4种处理均是30年生马尾松林分解最快,分解540 d时,30年生马尾松林对照处理较20年生和46年生快7.23%和5.74%,低氮处理分别快5.89%和3.41%,中氮处理分别快10.04%和10.10%,高氮处理分别快4.49%和7.43%。林分、N沉降和分解时间对分解影响均差异显著 (表 2,P<0.05),在N沉降作用下,分解前期促进分解,处理后期与对照相比抑制分解,随着时间的延续,N沉降对分解的影响越来越明显;而林龄对N沉降的响应主要表现在30年生林分对N沉降的正响应。
| 变异来源Source of variance | df | 平方和Mean square | F | P |
| 林龄Stand age | 2 | 87.09 | 39.36 | 0.00** |
| 分解时间Decomposition time | 5 | 1661.84 | 751.12 | 0.00** |
| N沉降N deposition | 3 | 5.52 | 2.50 | 0.08* |
| * P<0.05, ** P<0.01 | ||||
模拟N沉降抑制20年生和46年生马尾松林凋落叶分解过程中C释放 (图 2)。20年生林分分解前180 d中,N沉降促进凋落叶C释放,分解540 d后,凋落叶C含量中对照处理最低,C含量释放相对较高,C释放顺序是中氮 (1.98%)<低氮 (7.40%)<高氮 (7.84%)<对照 (8.67%),其中分解270 d时低氮和高氮差异显著 (P<0.05),分解540 d时对照与中氮处理差异显著 (P<0.05)。30年生林分各处理对C释放没有规律,处理540 d后释放顺序是对照 (5.66%)<高氮 (5.94%)<中氮 (10.68%)<低氮 (11.21%),与20年生林分相反,30年生林分凋落叶N沉降促进C释放,且中氮、高氮处理差异显著 (P<0.05)。46年生林分中,分解前180 d N沉降促进C释放,分解540 d后中、高氮处理明显抑制C释放并有少量固持,对照处理C含量最低,处理540 d后C释放顺序是高氮 (-0.19%)<中氮 (-0.52%)<低氮 (1.78%)<对照 (2.40%);分解270 d和360 d时,中氮处理明显高于对照 (P<0.05),分解450 d时高氮处理明显高于低氮处理 (P<0.05)。N沉降对30年生林分C释放影响最大,20年生次之,46年生最小。

|
| 图 2 不同林龄马尾松林凋落叶分解C、N和C/N比对N沉降的响应 Fig. 2 The response of litter leaf decomposition carbon, nitrogen contents and C/N ratio on different-aged P. massoniana under simulated nitrogen deposition 不同大写字母表示同一处理不同分解时间差异显著,不同小写字母表示同一分解时间不同处理间差异显著 |
模拟N沉降对凋落叶分解过程中N含量均净固持 (图 2)。20年生林分中N沉降处理对N固持影响不稳定,处理540 d后N含量顺序是低氮<高氮<中氮<对照,固持量分别为28.60%、34.08%、36.77%和42.47%,N沉降没有促进N固持;分解180 d和270 d时中氮和低氮之间差异显著 (P<0.05),450 d时中氮和对照差异显著 (P<0.05)。30年生林分N沉降处理对N固持影响不稳定,处理540 d后N含量顺序是对照<中氮<高氮<低氮,分别增加39.87%、41.53%、45.57%、45.72%,后期固持N含量比前期固持N含量稍快且分解360 d时N沉降各处理均与对照差异显著 (P<0.05)。46年生林分中高氮含量较对照中N含量高且分解360 d和540 d时中氮处理与对照差异显著 (P<0.05),处理540 d后,N含量顺序是对照<低氮<高氮<中氮,增量分别为34.51%、38.98%、51.08%、55.85%。
N沉降对马尾松林凋落叶分解C/N比影响差异显著 (图 2)。N沉降对3种马尾松林凋落叶C/N比影响类似,20年生林分分解90-270 d时C/N比下降明显且分解180 d时低氮与中氮差异显著 (P<0.05),分解270 d时低氮和高氮差异显著 (P<0.05),处理540 d后顺序是对照<高氮<中氮<低氮,分别下降36.19%、31.44%、28.71%、28.38%。30年生和20年生林分变化类似,各处理下降幅度相对均衡,540 d后N沉降处理促进C/N比下降,对照 (32.95%)<高氮 (35.90%)<中氮 (37.54%)<低氮 (39.68%),分解180 d时中氮和对照差异显著 (P<0.05),360 d时对照与N沉降各处理差异均显著 (P<0.05)。46年生林分处理540 d后C/N比降低比重分别是对照 (27.99%)<低氮 (29.92%)<高氮 (34.01%)<中氮 (34.62%) 且分解540 d时对照与中氮处理差异显著 (P<0.05)。
2.3 不同林龄马尾松林凋落叶分解中土壤有机碳和总氮动态不同林龄马尾松林土壤有机碳季节变化差异明显 (图 3)。20年生林分各处理季节变化不同,对照处理中土壤有机碳是3月<11月<9月<6月且差异显著 (P<0.05),低、中氮处理均是3月<6月<11月<9月,高氮处理是9月<6月<3月<11月,且3月高氮处理明显高于其它处理 (P<0.05);30年生马尾松林分对照、低和高氮处理是3月<9月<11月<6月,中氮处理是3月<9月<6月<11月,且只有高氮处理的6月份明显高于其它月份 (P<0.05);46年生马尾松林对照处理是6月<11月<3月<9月,低氮处理是6月<3月<9月<11月,中和高氮处理是9月<6月<3月<11月。4个季节均各处理均是30年生林分最低。

|
| 图 3 不同林龄马尾松林土壤有机碳、总氮及有机碳/总氮比季节动态对N沉降的响应 Fig. 3 The season dynamic of soil organic carbon, total nitrogen and the ratio of soil organic carbon/total nitrogen ratio on different-aged P. massoniana under simulated nitrogen deposition 不同大写字母表示同一处理季节动态差异显著 (P<0.05),不同小写字母表示同一月份不同处理差异显著性 (P<0.05) |
模拟N沉降处理下20年生和46年生马松林土壤总氮含量季节变化明显 (P<0.05)(图 3)。20年生马尾松林对照处理是11月<3月<6月<9月,低氮处理是6月<9月<3月<11月,中、高氮均是3月<9月<6月<11月;30年生马尾松林分除对照处理外均是9月最低;46年生马尾松林分均是11月最低,3月或6月最高。11月和9月,各处理均是30年生 (高氮处理外)<46年生<20年生;3月和6月各处理中除中氮外均是30年生马尾松林分最低。
模拟N沉降下C/N比季节变化明显 (P<0.05)(图 3)。20年生马尾松林对照处理中C/N比11月最高,之后相对恒定,低氮处理季节变化较小,中、高氮处理均是11月<6月<9月<3月且差异显著 (P<0.05);30年生马尾松林则是3月最低,6月最高 (MN除外) 且差异显著;46年生林分均是6月最低,11月最高。3种林龄各处理C/N比相对稳定,20年生林分C/N比在0.85-3.26间波动,30年生林分0.64-4.00间波动,46年生林分在0.90-4.80间波动,11月和9月相对较高,3月和6月相对较低。
2.4 模拟N沉降下不同马尾松林凋落叶-土壤C、N及其化学计量关系分析模拟N沉降处理下不同林龄马尾松林凋落叶-土壤分解中C、N及化学计量相互关系显著 (图 4)。4种处理下土壤有机碳、总氮及其SOC/TN比受凋落叶分解速率、凋落叶分解C和N动态的影响较大,与凋落叶C、N释放动态和分解速率成正相关,与凋落叶C/N比成负相关,即箭头越长、离其距离越远关系越紧密,对土壤养分影响越大。模拟N沉降4种处理中凋落叶化学计量不同,对照处理中分解速率<凋落叶C含量<凋落叶C/N比<凋落叶N含量,低、中和高氮处理中则是凋落叶C含量<分解速率<凋落叶N含量<凋落叶C/N比。土壤养分间同象限内夹角越小越相关,垂直则说明基本不相关,不同象限夹角余弦值为负值说明呈负相关。土壤有机碳与凋落叶C、N和分解速率成正比,与凋落叶C/N比呈显著负相关;土壤总氮与凋落叶分解速率、凋落叶N含量呈正相关,SOC/TN比与凋落叶C、N含量相关 (表 3)。

|
| 图 4 模拟N沉降下马尾松林凋落叶分解-土壤C、N及其化学计量动态的典范对应分析 Fig. 4 The canonical correspond analysis on litter leaf decomposition-soil C, N releasing dynamic and its stoichiometry on Pinus massoniana stands under simulated N deposition DR:凋落叶分解Litter leaf decomposition rate; SOC:土壤有机碳Soil organic carbon; TN:全氮Soil total nitrogen |
| 项目 Items | 凋落叶解 速率DR | 凋落叶碳 Litter leaf C | 凋落叶氮 Litter leaf N | 凋落叶C/N比 Litter C/N ratio | 土壤有机碳 SOC | 土壤总氮 TN | 土壤有机碳/ 全氮比 SOC/TN ratio |
| 凋落叶分解速率DR | 1 | ||||||
| 凋落叶碳litter leaf C | 0.48** | 1 | |||||
| 凋落叶氮litter leaf N | -0.263** | 0.111 | 1 | ||||
| 凋落叶C/N比C/N ratio | 0.416** | 0.250** | -0.918** | 1 | |||
| 土壤有机碳SOC | 0.275** | 0.284** | 0.418** | -0.342** | 1 | ||
| 土壤全氮TN | 0.265** | 0.098 | 0.235** | -0.212* | 0.569** | 1 | |
| 土壤有机碳/全氮比SOC/TN ratio | 0.014 | 0.192* | 0.190* | -0.142 | 0.429** | -0.449** | 1 |
| *P<0.05, ** P<0.01 | |||||||
N沉降在一定程度上满足了微生物对N素的需求,从而对凋落叶分解有一定的促进作用[23]。在氮沉降过程中,适量NO3-的输入可被森林生态系统所利用,过剩的NO3-将通过淋溶或反硝化作用从土壤中除去或有少部分在土壤中积累下来,当氮通量 (矿化和外部输入) 与生物和土壤的吸收能力相当时,生态系统可被认为是达到了“氮饱和”[24];因此,“氮饱和”程度是影响凋落叶分解的关键。本研究中模拟N沉降540 d后仅促进20年生和30年生马尾松林凋落叶分解,46年生马尾松林仅低氮处理促进分解,说明同一树种不同起始N含量对N沉降的响应不同,N沉降处理促进凋落叶起始N含量低的凋落叶分解,起始N含量高的凋落叶分解过程中易达到“氮饱和”;低氮处理明显增加凋落叶年分解速率,而高氮处理明显降低凋落叶年分解速率,因为N沉降提高有效态氮和土壤酶活性[25],也可能是高氮处理造成无机N富集后阻碍木质素和纤维素分解[14]。Knorr等[26]研究表明外源氮通常会促进N含量高或木质素含量低的凋落叶分解,而对N含量低或木质素含量高的凋落叶分解通常有抑制作用。Hobbi[27]研究结果表明N沉降通过凋落物加快N循环,N沉降对木质素含量高的凋落物比对木质素含量低的凋落物作用大,同时加快木质素含量低的凋落物生态系统短期碳损失。相反,本研究中模拟N沉降促进凋落叶N含量低的30年生马尾松林凋落叶分解,主要因为30年生马尾松林凋落叶和土壤中氮含量均较低,N输入缓解了N的限制,降低了C/N比、木质素/N比而促进分解,提高了凋落叶底物质量,从而加快了凋落叶的分解速率[28]。
N沉降对不同凋落叶分解阶段的影响不同:早期N沉降可能增加分解者的活性因而促进分解,后期阶段N沉降积累的N易于诱发C底物而限制分解者的活性[29]。汪金松[30]研究表明N沉降显著促进人工林针叶早期分解,抑制其后期分解。本研究中20年生林分、N沉降和分解时间对分解影响均差异显著 (表 2),在N沉降作用下,分解前期促进分解,处理后期与对照相比抑制分解,随分解的进行N沉降对分解的影响越来越明显 (46年生);而林龄对N沉降的响应主要表现在30年生林分对N沉降的正响应,支持莫江明等[31]的研究结果,即N沉降对凋落叶分解的影响随林分、树种和分解阶段不同,N沉降仅在试验开始3个月对马尾松针凋落叶分解存在明显的促进作用 (P<0.05)。相反,徐国良等[32]在南亚热带对N沉降下分解动态研究表明,N沉降处理抑制季风林凋落叶分解且随时间的差异愈来愈明显,促进混交林 (pine and leaf-broad mixed) 和针叶林 (pine forest) 凋落叶后期分解 (18个月),且这种差异由季风林达到“氮饱和”造成。王家骏等[33]研究表明模拟N沉降对米槠 (Castanopsis carlesii) 叶凋落叶的分解起到了抑制作用,在分解的720 d,中氮处理对分解速率影响较小,高氮处理抑制分解;涂利华等[14]研究表明在N沉降处理下,亮叶桦 (Betula luminifera) 凋落叶分解4-12个月,N沉降处理中的凋落叶分解速率均低于对照;在分解1年后N沉降处理比对照慢12.9%-19.6%。通常,土壤氮素含量较低的生态系统,凋落物的N素含量也较低,生态系统可能处于氮不饱和状态,N素成为限制生态系统功能的因子,一定程度外源N的输入可能弥补了生态系统对N素的需求而促进分解[11];N沉降对凋落叶分解的影响不大可能因为该森林具有较强的自我调节能力,调节系统“氮饱和”平衡点,适应N输入;N沉降抑制凋落叶分解可能因为无机N的积累改变了生态系统总的营养平衡,抑制了参与凋落物分解的胞外酶活性、微生物和动物的活动[32]。
3.2 模拟N沉降对马尾松林凋落叶分解-土壤C、N化学计量的影响3种马尾松林凋落叶C、N含量和C/N比对N沉降响应明显。20年生和46年生马尾松林在处理540 d后N沉降处理碳释放速率均小于对照,而30年生林分中N沉降各处理碳释放率均明显高于对照 (图 2),说明模拟N沉降处理起始C含量高的凋落叶不利于C释放,起始C含量低的凋落叶促进C释放;N沉降促进30年生和46年生N固持,分解540 d后,30年生马尾松林氮处理中N含量分别比对照高1.19%-4.19%,46年生则分别高3.32%-15.86%,20年生马尾松则分别较对照低4.00%-9.73%(图 2),说明凋落叶分解中N含量固持与起始N含量有关,起始N含量低的凋落叶N固持量高于起始N含量高的凋落叶,因为N沉降影响分解者群落适应低N底物而分解者所需要的化学计量比[34],N沉降降低了30年生马尾松林的C/N比,缓解了N的相对限制,刺激了微生物活性,与30年生相对较快的分解速率一致,表明马尾松分解者群落有一个较高的化学计量分解比,凋落叶分解过程中养分积累或释放由起始N含量和微生物分解者的化学计量决定。因此,N素增加对凋落叶C含量的影响较复杂,既取决于生态系统分解者的种类和活性,也受到气候、树种及立地条件等[28]。
模拟N沉降对马尾松林土壤有机碳、总氮及有机碳/总氮比动态变化影响明显,N沉降促进低土壤养分马尾松林凋落叶分解并提高土壤肥力。N沉降处理下,30年生马尾松林中各个月份土壤有机质对照均低于加N处理,而土壤总氮含量生长季 (3-9月) 对照低于N处理,非生长季 (11-3月) 对照处理土壤总氮高于N处理 (图 3),说明土壤总氮含量低的林分生长季受N限制明显;20年生马尾松林N沉降初期 (11月) 中氮和高氮处理下土壤总氮明显增加 (P<0.05),46年生马尾松林随着分解的进行土壤总氮先增加后降低且各处理差异不显著,可能因为N沉降增加了土壤和凋落物层的矿质氮含量,增加了土壤的矿化硝化,从而增加了土壤中的有效氮 (硝态氮和铵态氮) 的量[23],随后植物和土壤在短期内吸收了有效态氮 (尽管森林土壤N饱和)[35],使20年生和46年生马尾松林土壤中有效氮素增加后被快速利用和流失致使土壤总氮含量降低或不变。涂利华等[13]对华西雨屏区苦竹 (Pleioblastus amarus) 人工林模拟N沉降试验1年研究表明,N沉降显著增加了土壤总有机碳、全氮,N沉降提高了土壤中C、N元素的活性,土壤有机C和养分对N沉降呈正响应。相反,汪金松[30]研究表明N沉降引起土壤各层有机C含量减少,表层土壤全N含量增加。亦有研究表明N添加降低土壤中原有有机质和腐殖的分解,土壤异养呼吸和总呼吸的降低导致土壤有机碳吸存增加[29]。总之,不同森林类型,N沉降通过土壤N含量影响植物的养分利用效率[3]、短期N固持和生态系统N循环[35],通过对土壤有机质的作用 (通过多酚混合物)、植物C分配、微生物群落结构的改变 (如从真菌为主转化成细菌群落为主),微生物酶产量[29]和根系分泌物的变化使分配到根系的C含量降低[3],致使土壤总有机碳和总氮有不同的响应。
凋落物通过分解控制土壤中短期有机质的分解和积累而调节植物-土壤关系,土壤肥力与凋落叶N动态相关,肥力较高的土壤通常有较高的N固持或N释放[36],凋落叶分解速率中凋落叶N含量和土壤N有效性相关[37]。王家骏等[33]研究表明N沉降使中亚热带常绿阔叶林土壤-凋落叶内可得性N增加,改变了原有的养分平衡,抑制了微生物活性,降低了分解酶的活性而调节米槠叶凋落叶的分解速率。本研究中模拟N沉降处理下不同林龄马尾松林凋落叶分解过程中发现土壤-凋落叶C、N化学计量与分解相互作用显著 (图 4),土壤有机质与凋落叶C、N和分解速率成正比,与凋落叶C/N比呈显著负相关;土壤总氮与凋落叶分解速率、凋落叶N含量呈正相关,土壤有机质/总氮比与凋落叶C、N含量相关,说明凋落叶-土壤碳、氮及化学计量相互作用显著,且凋落叶质量和土壤养分低的生态系统土壤C对N沉降响应更明显 (30年生马尾松林)[38],凋落叶分解过程中有效N的变化与土壤特征如微生物群落相联系[39]。总之,研究表明N沉降改变了森林土壤微生物群落结构及其土壤真菌群落,从而改变凋落叶分解过程和土壤C、N循环[40],通常以难分解凋落物、白腐真菌为主要分解者的生态系统对N沉降呈现负响应,而含有易分解凋落物、维持不同真菌种群的生态系统对N沉降呈现正响应;土壤养分含量低的生态系统土壤C对N沉降响应更显著[38]。
| [1] | Liu X J, Duan L, Mo J M, Du E, Shen J L, Lu X K, Zhang Y, Zhou X B, He C E, Zhang F S. Nitrogen deposition and its ecological impact in China:an overview. Environment Pollution, 2011, 159(10): 2251–2264. DOI:10.1016/j.envpol.2010.08.002 |
| [2] | 刘文静, 康荣华, 张婷, 朱婧, 段雷. 我国西南地区氮饱和马尾松林土壤和植物15N自然丰度对长期氮施加的响应. 环境科学, 2015, 36(8): 2981–2987. |
| [3] | Gu F X, Zhang Y D, Huang M, Tao B, Yan H M, Guo R, Li J. Nitrogen deposition and its effect on carbon storage in Chinese forests during 1981-2010. Atmospheric Environment, 2015, 123(A): 171–179. |
| [4] | Frey S D, Ollinger S, Nadelhoffer K, Bowden R, Brzostek E, Burton A, Caldwell B A, Crow S, Goodale C L, Grandy A S, Finzi A, Kramer M G, Lajtha K, LeMoine J, Martin M, McDowell W H, Minocha R, Sadowsky J J, Templer P H, Wichings K. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests. Biogeochemistry, 2014, 121(2): 305–316. DOI:10.1007/s10533-014-0004-0 |
| [5] | Du Y H, Guo P, Liu J Q, Wang C Y, Yang N, Jiao Z X. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Global Change Biology, 2014, 20(10): 3222–3228. DOI:10.1111/gcb.2014.20.issue-10 |
| [6] | Austin A T, Ballaré C L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(10): 4618–4622. DOI:10.1073/pnas.0909396107 |
| [7] | 宋新章, 江洪, 张慧玲, 余树全, 周国模, 马元丹, ChangS X. 全球环境变化对森林凋落物分解的影响. 生态学报, 2008, 28(9): 4414–4423. |
| [8] | Ge X G, Zeng L X, Xiao W F, Huang Z L, Geng X S, Tan B W. Effect of litter substrate quality and soil nutrients on forest litter decomposition:A review. Acta Ecologica Sinica, 2013, 33(2): 102–108. DOI:10.1016/j.chnaes.2013.01.006 |
| [9] | Berg B, Matzner E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environmental Reviews, 1997, 5(1): 1–25. DOI:10.1139/a96-017 |
| [10] | 方运霆, 莫江明, GundersenP, 周国逸, 李德军. 森林土壤氮素转换及其对氮沉降的响应. 生态学报, 2004, 24(7): 1523–1531. |
| [11] | 徐国良, 莫江明, 周国逸. 模拟氮沉降增加对南亚热带主要森林土壤动物的早期影响. 应用生态学报, 2005, 16(7): 1235–1240. |
| [12] | 李考学. 氮沉降对长白山两种主要针叶树种凋落物分解的影响. 东北林业大学学报, 2007, 35(2): 17–19. |
| [13] | 涂利华, 胡庭兴, 张健, 李仁洪, 戴洪忠, 雒守华. 模拟氮沉降对华西雨屏区苦竹林土壤有机碳和养分的影响. 植物生态学报, 2011, 35(2): 125–136. |
| [14] | 涂利华, 胡红玲, 胡庭兴, 张健, 雒守华, 戴洪忠. 华西雨屏区亮叶桦凋落叶分解对模拟氮沉降的响应. 植物生态学报, 2012, 36(2): 99–108. |
| [15] | 弓晓静, 余明泉, 胡小飞, 郭恢财, 陈伏生. 氮磷添加对红壤区城郊湿地松林凋落叶分解的影响. 生态学杂志, 2010, 29(12): 2327–2333. |
| [16] | 鲁显楷, 莫江明, 董少峰. 氮沉降对森林生物多样性的影响. 生态学报, 2008, 28(11): 5532–5548. |
| [17] | 宋学贵, 胡庭兴, 鲜骏仁, 肖春莲. 川南天然常绿阔叶林土壤酶活性及其对模拟N沉降的响应. 生态学报, 2009, 29(3): 1234–1240. |
| [18] | 袁颖红, 樊后保, 刘文飞, 张子文, 孟庆银, 胡锋, 李辉信. 模拟氮沉降对杉木人工林土壤可溶性有机碳和微生物量碳的影响. 水土保持学报, 2012, 26(2): 138–143. |
| [19] | 葛晓改.三峡库区马尾松林凋落物分解及对土壤碳库动态的影响研究[D].北京:中国林业科学研究院, 2012, 193-193. |
| [20] | 肖文发, 李建文, 于长青, 马娟, 程瑞梅, 刘少英, 王金锡, 葛继稳. 长江三峡库区陆生动植物生态. 重庆: 西南师范大学出版社, 2000. |
| [21] | 葛晓改, 肖文发, 曾立雄, 黄志霖, 付甜, 封晓辉. 不同林龄马尾松凋落物基质质量与土壤养分的关系. 生态学报, 2012, 32(3): 852–862. |
| [22] | 国家林业局. 中华人民共和国林业行业标准-森林土壤分析方法. 北京: 中国标准出版社, 2000. |
| [23] | 樊后保, 刘文飞, 李燕燕, 廖迎春, 袁颖红, 徐雷. 亚热带杉木 (Cunninghamia lanceolata) 人工林生长与土壤养分对氮沉降的响应. 生态学报, 2007, 27(11): 4630–4642. |
| [24] | 肖辉林. 大气氮沉降对森林土壤酸化的影响. 林业科学, 2001, 37(4): 111–116. |
| [25] | Song X Z, Zhou G M, Gu H H, Qi L H. Management practices amplify the effects of N deposition on leaf litter decomposition of the Moso bamboo forest. Plant and Soil, 2015, 395(1): 391–400. |
| [26] | Knorr M, Frey S D, Curtis P S. Nitrogen additions and litter decomposition:a meta-analysis. Ecology, 2005, 86(12): 3252–3257. DOI:10.1890/05-0150 |
| [27] | Hobbie S E. Interactions between litter lignin and soil nitrogenitter availability during leaf litter decomposition in a Hawaiian Montane forest. Ecosystems, 2000, 3(5): 484–494. DOI:10.1007/s100210000042 |
| [28] | 樊后保, 刘文飞, 徐雷, 李燕燕, 廖迎春, 王启其, 张子文. 氮沉降下杉木 (Cunninghamia lanceolata) 人工林凋落叶分解过程中C、N元素动态变化. 生态学报, 2008, 28(6): 2546–2553. |
| [29] | Tonitto C, Goodale C L, Weiss M S, Frey S D, Ollinger S V. The effect of nitrogen addition on soil organic matter dynamics:a model analysis of the Harvard Forest Chronic Nitrogen Amendment Study and soil carbon response to anthropogenic N deposition. Biogeochemistry, 2014, 117(2-3): 431–454. DOI:10.1007/s10533-013-9887-4 |
| [30] | 汪金松.模拟氮沉降对暖温带油松林土壤碳循环过程的影响[D].北京:北京林业大学, 2013, 149-149. |
| [31] | 莫江明, 薛璟花, 方运霆. 鼎湖山主要森林植物凋落物分解及其对N沉降的响应. 生态学报, 2004, 24(7): 1413–1420. |
| [32] | 徐国良, 莫江明, 周国逸, 薛璟花. 氮沉降下鼎湖山森林凋落物分解及与土壤动物的关系. 生态环境, 2005, 14(6): 901–907. |
| [33] | 王家骏, 程煜, 杨玉盛, 林成芳, 林伟盛, 彭建勤. 米槠叶凋落物分解及养分释放对模拟N沉降的响应. 福建林学院学报, 2014, 34(2): 113–119. |
| [34] | Finn D, Page K, Catton K, Strounina E, Kienzle M, Robertson F, Armstrong R, Dalal R. Effect of added nitrogen on plant litter decomposition depends on initial soil carbon and nitrogen stoichiometry. Soil Biology and Biochemistry, 2015, 91: 160–168. DOI:10.1016/j.soilbio.2015.09.001 |
| [35] | Gurmesa G A, Lu X K, Gundersen P, Mao Q G, Zhou K J, Fang Y T, Mo J M. High retention of 15N-labeled nitrogen deposition in a nitrogen saturated old-growth tropical forest. Global Change Biology, 2016. DOI:10.1111/gcb.13327 |
| [36] | Norris M D, Avis P G, Reich P B, Hobbie S E. Positive feedbacks between decomposition and soil nitrogen availability along fertility gradients. Plant and Soil, 2013, 367(1-2): 347–361. DOI:10.1007/s11104-012-1449-3 |
| [37] | Hobbie S E, Vrrousek P M. Nutrient limitation of decomposition in Hawaiian forests. Ecology, 2000, 81(7): 1867–1877. DOI:10.1890/0012-9658(2000)081[1867:NLODIH]2.0.CO;2 |
| [38] | Waldrop M P, Zak D R, Sinsabaugh R L, Gallo M, Lauber C. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecological Applications, 2004, 14(4): 1172–1177. DOI:10.1890/03-5120 |
| [39] | Delgado-Baquerizo M, García-Palacios P, Milla R, Gallardo A, Maestre F T. Soil characteristics determine soil carbon and nitrogen availability during leaf litter decomposition regardless of litter quality. Soil Biology and Biochemistry, 2015, 81: 134–142. DOI:10.1016/j.soilbio.2014.11.009 |
| [40] | Freedman Z B, Romanowicz K J, Upchurch R A, Zak D R. Differential responses of total and active soil microbial communities to long-term experimental N deposition. Soil Biology and Biochemistry, 2015, 90: 275–282. DOI:10.1016/j.soilbio.2015.08.014 |