 2017, Vol. 37
2017, Vol. 37文章信息
- 池源, 石洪华, 王恩康, 郭振, 丰爱平, 麻德明
- CHI Yuan, SHI Honghua, WANG Enkang, GUO Zhen, FENG Aiping, MA Deming.
- 庙岛群岛北五岛景观格局特征及其生态效应
- Landscape pattern characteristics and ecological effects on five northern islands of Miaodao Archipelago
- 生态学报. 2017, 37(4): 1270-1285
- Acta Ecologica Sinica. 2017, 37(4): 1270-1285
- http://dx.doi.org/10.5846/stxb201604130672
-
文章历史
- 收稿日期: 2016-04-13
- 修订日期: 2016-08-09

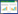




景观格局是自然和人文因子综合作用于地理空间上的表现结果, 随着人类活动的日益频繁, 地表景观格局愈发受到人为干扰的作用[1], 而景观格局特征及变化又对生态系统的结构、功能和过程产生着重要影响[2-6]。海岛是重要生态功能的贮存库, 是维护国家权益的重要平台, 也是人类居住生活的载体以及保护与利用海洋的支点[7-9]。由于独特的自身条件和复杂的外界干扰, 海岛生态系统具有明显的脆弱性, 主要表现在其面临干扰时更容易受到损害, 且难以通过自身调节能力恢复[9-12]。近年来, 海岛开发利用活动日趋频繁, 造成海岛自然景观割裂, 人工景观增多, 景观格局发生显著变化[13-16]。景观格局深刻地作用于海岛生态系统[17], 植被生产力、植物多样性和土壤肥力作为海岛生态系统功能的重要表现形式, 均受到景观类型和格局变化的影响[18-19]。岛群是区域中位置相邻海岛的群聚形态, 岛群中不同海岛在面积、形状等基础地理信息上可能存在较大不同[20], 海岛内部在地形等方面也具有明显的空间差异, 再加上人类活动的非均质发展, 造成海岛景观格局分异性愈加明显, 生态系统效应更加复杂。同时, 不同海岛相互联系又相对独立, 景观格局对人类活动响应灵敏且空间差异性显著, 使得海岛成为景观格局及其生态效应研究的天然试验场[21]。然而, 目前关于海岛景观格局特征及其与生态系统服务功能相互关系的深入分析还鲜见报道。
本文以庙岛群岛北五岛为研究区, 以海岛景观格局为核心, 基于现场调查和3S技术, 从景观尺度、海岛尺度和区块尺度对景观格局特征进行分析, 进而探讨景观格局对海岛植被净初级生产力(NPP)、植物多样性和土壤性质的影响, 以期阐释自然和人为因子影响下海岛生态系统的变化特征, 为海岛生态系统保护提供参考, 并从其区域典型性上丰富景观格局特征及其生态效应的研究。
1 研究区概况庙岛群岛位于山东半岛北侧, 黄渤海交汇处, 是我国北方海岛的集中分布区。砣矶岛、大钦岛、小钦岛、南隍城岛和北隍城岛共5个有居民海岛构成庙岛群岛北部区域的主体, 合称“北五岛”。北五岛位于渤海海峡中部, 海岛之间较为分散, 呈典型的链状分布(图 1) 。该区域属于东亚季风气候区, 年均气温12.0℃, 1月平均气温-1.6℃, 7月平均气温24.5℃;年均降水量537.1mm, 降水多集中在6—9月;日照较为充足, 年均日照时数2612h。北五岛地势起伏明显, 最高点海拔为202m, 位于大钦岛。海岛原生林木发育较差, 广泛种植了人工防护林, 以黑松和刺槐为优势种;原生草本植物种类繁多, 分布广泛[18]。北五岛以海水养殖、捕捞、旅游为主要产业, 不同海岛之间社会经济发展具有差异, 海岛作为开发和保护海洋的重要支点和海岛居民生活的主要载体, 海洋经济的发展直接带动了海岛空间的开发利用, 住宅及公共设施建造、道路码头建设、农田开垦、人工林种植等使得北五岛景观类型复杂多样。

|
| 图 1 庙岛群岛北五岛景观类型和植物群落调查样地 Fig. 1 Landscape types and plant community sampling plots on five northern islands orthern slands of Miaodao Archipelago |
采用WorldView-1卫星2013年全色波段遥感影像, 空间分辨率为0.45m。通过ArcGIS 10.0软件提取海岛轮廓, 得到海岛面积、周长等基本信息;进而开展人机交互解译, 将北五岛景观分为针叶林、阔叶林、草地、农田、裸地、建筑用地、交通用地和其他硬化地面8类, 其中, 针叶林和阔叶林分别是以黑松和刺槐为优势种的人工林, 草地主要是海岛天然原生草本群落, 农田以种植大豆和玉米等农作物为主, 裸地是指海岸和岛屿内部的裸岩和裸土地, 建筑用地包括岛内住宅和公共服务建筑, 交通用地主要包括海岛码头和岛内道路, 其他硬化地面是指建筑用地和交通用地以外人工铺装的密实地面, 主要为海带晒场和广场等。
采用LANDSAT8卫星2013年8月11日、11月15日和2014年1月2日和4月24日(代表不同季节)4个时相30m分辨率的无云影像。利用ENVI 4.7软件对影像进行裁切、辐射定标、波段运算得到不同季节的NDVI值(图 1) 。采用2011年公布的Aster GDEM第二版DEM数据, 垂直分辨率20m, 水平分辨率30m;通过ArcGIS 10.0由DEM数据中提取海拔和坡度。
2.1.2 现场调查2014年7月开展北五岛现场踏勘, 对北五岛地表覆盖类型进行现场验证, 根据验证结果对现有数据进行校正, 形成最终的北五岛景观类型矢量图(图 1) 。
2015年9月开展北五岛植物群落现场调查, 综合考虑海岛面积、群落类型和地形等因素, 共布设60个20×20m调查样地(图 1) 。运用GPS手持机和电子罗盘测量样地的经纬度、海拔、坡度和坡向。记录样方内出现的全部乔木种, 测量所有DBH≥3 cm的植株胸径、树高、冠幅等信息, 记录其存活状态;记录样方内出现的全部灌木种, 选择面积为10 m×10 m的两个对角小样方进行调查, 对其中的全部灌木分种计数, 并测量基径、高度等信息;记录样方内出现的全部草本种类, 在每个样地的4角和中心共设置5个1×1m草本植物样方, 记录样方内草本植物种类、多度、盖度、高度等信息。此外, 采用多点混合取土样方法进行表层土壤取样, 在每个样地内选取3个取样点, 经均匀混合后作为该样地的土壤样品, 在实验室内除去其中草根、大石砾等杂质, 过2 mm钢筛并磨细, 在105℃高温下烘至恒重, 称量并计算土壤含水量;全氮采用半微量凯氏法, 速效磷采用盐酸-氟化铵法, 速效钾采用乙酸铵提取法, 有机质采用重铬酸钾氧化外加热法。
2.1.3 资料收集气象数据来自长岛县气象站和烟台福山气象站监测的多年数据, 各岛的人口和生产总值来自长岛县统计局相关资料。
2.2 景观格局空间特征分析以北五岛景观类型矢量图为基础, 通过ArcGIS 10.0中的Patch Analyst模块, 采用景观格局指数, 从景观尺度、海岛尺度和区块尺度对景观格局特征进行分析。由于Patch Analyst模块中景观格局指数数量较多, 且相互之间具有一定的相关性, 这里挑选具有代表性的景观格局指数:斑块密度(PD)反映景观格局破碎化程度, 边缘密度(ED)反映景观格局边缘效应, 平均形状指数(MSI)反映景观斑块形状复杂性。同时, 为了反映人类活动对自然生态系统的干扰强度, 本文建立了人为干扰指数(HII)指标, 其具体计算方法如下:
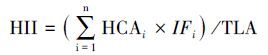
|
(1) |
式中, HCAi表示人工景观的面积, HCA1-HCA6分别表示交通用地、建筑用地、其他硬化地面、农田、黑松林和刺槐林;IFi表示第i类人工景观对海岛生态系统的影响系数, 不同人工景观对海岛生态系统的影响方式和程度具有较大差别, 考虑到人工景观的特征和海岛实际情况, IF1取1.0, IF2和IF3取0.6, IF4取0.4, IF5和IF6取0.2;TLA表示分析尺度内的景观总面积。
采用Patch Analyst模块中的Spatial Statistics工具, 计算景观尺度和海岛尺度的景观格局指数;利用Fishnet工具将北五岛划分为200m×200m区块单元, 采用Analysis by Regions工具分析不同区块单元的景观格局指数[17]。
2.3 景观格局的生态效应分析基于收集资料、现场调查数据和遥感影像, 分别计算海岛NPP、植物多样性和土壤性质3方面生态效应指标。
借助不同季节的遥感影像, 依据CASA模型[22], 构建海岛NPP估算模型如下:
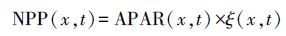
|
(2) |

|
(3) |
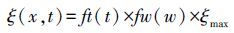
|
(4) |
式中,NPP(x, t)为x点t月净初级生产力;APAR(x, t)是指x点t月吸收的光合有效辐射(MJ m-2 month-1);ξ(x, t) 表示x点t月的实际光能利用率(g C/MJ);PAR(x, t)为x点t月的光合有效辐射(MJ m-2 month-1);FPAR(x, t)为x点t

|
| 图 2 北五岛NPP空间分布特征 Fig. 2 NPP distribution characteristics on five northern islands |
庙岛群岛乔木和灌木种类总体较少, 但草本植物发育完整且分布广泛[19], 这里以草本植物为代表进行海岛植物多样性计算。现场样地调查共记录草本植物93种, 分属28科、68属。计算各样地内草本植物的重要值, 方法如下[23]:
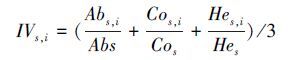
|
(5) |
式中, IVs, i为样地s中物种i的重要值, Abs, i为样地s内物种i的多度, Abs为样地s物种多度之和, Cos, i为样地s内物种i的盖度, Cos为样地s物种盖度之和, Hes, i为样地s内物种i的高度, Hes为样地s物种高度之和。
采用目前在国内外相关研究中普遍应用的Shannon-Wiener指数(H′)和Pielou指数(E)计算北五岛草本植物多样性, 前者侧重于反映群落物种的复杂程度, 后者则更加强调群落物种的均匀度。计算方法如下[24]:
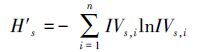
|
(6) |

|
(7) |
式中, H′s、Es分别为样地s的Shannon-Wiener指数和Pielou指数, Ns为样地s的物种数量。基于60个样地的多样性计算结果, 通过ArcGIS 10.0空间分析模块中的内插法进行多样性空间插值, 对比不同空间插值结果, 发现反距离权重(IDW)内插法的结果误差相对较小, 更符合海岛实际情况, 因此这里采用IDW内插法模拟海岛植物多样性空间分布特征。
统计海岛土壤含水量、总氮、速效磷、速效钾和有机质因子, 采用IDW内插法得到海岛土壤性质空间分布特征。计算土壤肥力指数, 方法如下:
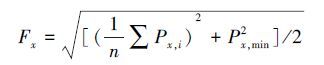
|
(8) |

|
(9) |
式中, Fx为x点的土壤肥力指数, Px, i为x点因子i的肥力状况, Cx, i为x点因子i的现状值, Si为因子i的标准值, 包括总氮、速效磷、速效钾和有机质4项因子, 采用《绿色食品产地环境质量标准》(NY/T391-2000) 中土壤肥力分级参考指标的旱地Ⅰ级作为标准值。最终以土壤含水量(MC)和土壤肥力(F)作为景观格局对土壤性质的生态效应指标。
分别从景观尺度、海岛尺度、区块尺度分析景观格局对海岛NPP、植物多样性和土壤性质的影响。景观尺度上, 采用单因素方差分析探讨不同景观类型各生态效应指标的差异和特征;海岛尺度上, 采用回归分析分析各岛景观格局指数与生态效应指标的关系;区块尺度上, 运用Zonal Statistics工具, 提取不同内区块内各生态效应指标的平均值, 采用相关分析法探讨区块尺度上景观格局指数与生态效应指标的简单相关性, 进而以植被景观为控制变量, 分析景观格局指数与生态效应指标两两之间的偏相关性。
3 结果与分析 3.1 北五岛景观格局空间特征 3.1.1 景观尺度不同景观类型中, 针叶林、草地、阔叶林面积较大, 其次为建筑用地, 裸地也具有一定规模, 其他硬化地面、交通用地和农田面积较小, 不足100hm2(表 1) 。草地和针叶林各项景观格局指标均较高, 阔叶林边缘密度和平均形状指数也较高, 但斑块密度偏低;建筑用地和交通用地分别占据了平均形状指数的最低值和最高值, 但前者斑块密度较高, 后者斑块密度较低;裸地各项指标均处于中间位置, 农田和其他硬化地面各景观指标均较低(表 2) 。
| 项目Items | 针叶林 Coniferous forest | 阔叶林 Broad-leaf forest | 草地 Grassland | 农田 Farmland | 裸地Bare land | 建筑用地 Construction land | 交通用地 Traffic land | 其他硬化地面 Other hardened ground | 总计Total |
| 砣矶岛 | 157.08 | 213.92 | 158.96 | 10.72 | 42.10 | 119.54 | 17.76 | 2.13 | 722.20 |
| 大钦岛 | 169.73 | 74.17 | 179.14 | 2.94 | 41.65 | 101.92 | 8.54 | 69.92 | 648.01 |
| 小钦岛 | 36.13 | 17.11 | 35.63 | 0 | 14.98 | 11.87 | 1.38 | 0 | 117.09 |
| 南隍城岛 | 64.48 | 15.75 | 44.20 | 0 | 29.51 | 28.78 | 6.35 | 0 | 189.06 |
| 北隍城岛 | 103.64 | 34.48 | 36.14 | 0 | 48.81 | 43.47 | 2.75 | 0 | 269.27 |
| 总计Total | 531.05 | 355.43 | 454.06 | 13.66 | 177.04 | 305.58 | 36.76 | 72.05 | 1945.64 |
| 景观类型 Landscape | PD | ED | MSI |
| 针叶林 Coniferous forest | 0.158 | 117.16 | 2.52 |
| 草地 Grassland | 0.212 | 160.37 | 2.27 |
| 裸地 Bare land | 0.112 | 74.4 | 2.06 |
| 交通用地 Traffic land | 0.02 | 52.97 | 6.91 |
| 阔叶林 Broad-leaf forest | 0.074 | 85.43 | 2.24 |
| 农田 Farmland | 0.019 | 5.55 | 1.48 |
| 建筑用地 Construction land | 0.187 | 58.63 | 1.44 |
| 其他硬化地面Other hardened ground | 0.069 | 28.19 | 2.03 |
不同海岛面积差异明显, 各岛的景观结构总体较为相似, 但又互有差异。面积较大的海岛景观类型更加多样, 农田和其他硬化地面仅在砣矶岛和大钦岛出现, 农田主要分布于砣矶岛, 其他硬化地面主要以海带晒场的形式集中于大钦岛;北隍城岛面积处于中间位置, 却拥有最大规模的裸地;南隍城岛面积较小, 但交通用地面积明显较高;小钦岛面积最小, 其各类景观规模也都处于最小值(表 1) 。就景观格局指数而言, 各岛斑块密度由大到小依次为砣矶岛、大钦岛、小钦岛、南隍城岛和北隍城岛, 平均形状指数与之完全相反, 边缘密度大小顺序除了南隍城岛明显较高, 其他与斑块密度基本一致, 人为干扰指数大钦岛和砣矶岛明显较高, 其次为南隍城岛和北隍城岛, 小钦岛最小(表 3) 。
| 海岛Island | PD | ED | MSI | HII |
| 北五岛 Five northern islands | 0.851 | 582.7 | 2.18 | 0.226 |
| 大钦岛Daqin Island | 0.832 | 569.96 | 2.12 | 0.248 |
| 南隍城岛Nanhuangcheng Island | 0.603 | 586.75 | 2.63 | 0.21 |
| 砣矶岛Tuoji Island | 1.098 | 655.4 | 2.04 | 0.228 |
| 小钦岛Xiaoqin Island | 0.632 | 548.24 | 2.29 | 0.164 |
| 北隍城岛Beihuangcheng Island | 0.512 | 435.18 | 2.98 | 0.21 |
各景观格局指标在区块尺度上均表现出了更加明显的异质性(图 3) 。斑块密度和边缘密度在砣矶岛和大钦岛内部空间分布差异显著, 但在南隍城岛和北隍城岛上不明显, 平均形状指数则与之表现出相反的特征;人为干扰指数空间分布呈现“两极化”特征, 在砣矶岛和大钦岛尤为典型, 说明人为干扰在海岛内部空间上具有区域集中性。同时, 小钦岛由于面积较小, 其内部景观格局具有一定的空间分异性, 但不甚显著。

|
| 图 3 北五岛景观格局空间特征 Fig. 3 Spatial characteristics of landscape pattern on |
不同景观类型的各项生态效应指标均具有显著性差异(P<0.01) 。就NPP而言, 阔叶林最高, 针叶林、农田和草地其次, 交通用地、其他硬化地面、建筑用地和裸地较低。就多样性指标而言, Shannon-wiener指数在针叶林和草地中明显较高, 建筑用地、裸地、交通用地和其他硬化地面其次, 阔叶林和农田较低;Pielou指数在针叶林中最高, 其次为草地、农田、交通用地和裸地, 阔叶林、建筑用地和其他硬化地面总体偏低。就土壤指标而言, 含水量在针叶林和刺槐林较高, 其次为草地和农田, 其余景观类型总体偏低;肥力在针叶林、阔叶林和草地中较高, 农田则明显偏低, 其余类型则处于中间位置(表 4) 。
| 景观类型 Landscape | NPP/(g C m-2 a-1) | H′ | E | MC/% | F |
| 针叶林Coniferous forest | 408.11 | 1.710 | 0.873 | 21.66 | 1.83 |
| 阔叶林Broad-leaf forest | 584.00 | 1.563 | 0.864 | 21.79 | 1.76 |
| 草地Grassland | 308.20 | 1.712 | 0.871 | 20.16 | 1.67 |
| 农田Farmland | 390.72 | 1.489 | 0.871 | 19.41 | 1.38 |
| 裸地Bare land | 104.46 | 1.595 | 0.868 | 16.16 | 1.53 |
| 建筑用地Construction land | 161.51 | 1.588 | 0.864 | 15.632 | 1.48 |
| 交通用地Traffic land | 265.18 | 1.643 | 0.869 | 16.32 | 1.39 |
| 其他硬化地面Other hardened ground | 200.80 | 1.639 | 0.863 | 16.464 | 1.44 |
不同海岛的各项生态效应指标也均具有显著性差异(P<0.01) 。分析海岛尺度上景观格局指数与生态效应指标的关系可得(图 4—图 6) , NPP平均密度、Shannon-Wiener指数和土壤含水量与各景观格局指数均未表现出明显的关系, Pielou指数随着边缘密度的增加显著上升, 土壤肥力随着边缘密度的增加显著下降, 但与其他指数未见明显关系。

|
| 图 4 海岛尺度上景观格局指数与NPP平均密度的关系 Fig. 4 Relationship between landscape pattern indices and mean density of NPP in island scale |

|
| 图 5 海岛尺度上景观格局指数与生物多样性的关系 Fig. 5 Relationship between landscape pattern indices and biodiversity in island scale |

|
| 图 6 海岛尺度上景观格局指数与土壤性质的关系 Fig. 6 Relationship between landscape pattern indices and soil property in island scale |
区块尺度上景观格局指数与生态效应指标的简单相关分析结果显示(表 5) , NPP平均密度与斑块密度、边缘密度和平均形状指数呈显著正相关, 与人为干扰指数呈显著负相关;Shannon-Wiener指数与斑块密度、边缘密度和人为干扰指数呈显著负相关, 与平均形状指数呈显著正相关, Pielou指数与人为干扰指数显著负相关, 与其他指标未见显著相关性;土壤含水量与斑块密度、边缘密度和平均形状指数呈显著正相关, 与人为干扰指数呈显著负相关, 土壤肥力与各景观格局指数均无显著相关性。以植被景观为控制变量的偏相关分析结果与简单相关分析表现出了明显的差异(表 5) , NPP和土壤含水量与大部分景观格局指数的相关系数明显减小, 相关性变为不显著;Shannon-Wiener指数与各景观格局指数的偏相关性仍显著, 其中与斑块密度和边缘密度的负相关性明显增大, 与平均形状指数和人为干扰指数的相关性有所减弱;Pielou指数与各景观格局指标的相关性仍不显著;土壤养分与景观格局指标的偏相关性相比简单相关性有了明显的增强, 其中与斑块密度和边缘密度呈显著负相关, 与人为干扰指数呈显著正相关。
| 项目 Items | PD | ED | MSI | HII | 项目 Items |
| 简单相关 | NPP | 0.239** | 0.293** | 0.229** | -0.593** |
| Simple correlation | H′ | -0.137** | -0.096** | 0.120** | -0.211** |
| E | 0.005 | 0 | 0.051 | -0.094* | |
| MC | 0.187** | 0.234** | 0.116** | -0.108** | |
| F | -0.013 | 0.018 | 0.054 | -0.054 | |
| 偏相关 | NPP | -0.036 | 0.099* | 0.091 | 0.141** |
| Partial correlation | H′ | -0.203** | -0.192** | 0.096* | -0.165** |
| E | -0.022 | -0.039 | -0.04 | -0.071 | |
| MC | 0.07 | 0.074 | 0.064 | 0.026 | |
| F | -0.174** | -0.173** | 0.002 | 0.091* | |
| **在0.01水平(双侧)显著相关, *在 0.05水平(双侧)显著相关 | |||||
(1) 研究区典型性
庙岛群岛全境属山东省长岛县管辖, 北五岛是长岛县北部的5个有居民海岛, 与之相对应的是由南长山岛、北长山岛、庙岛、小黑山岛和大黑山岛组成的“南五岛”, 南五岛是长岛县政治、经济和文化中心, 距离大陆较近, 海岛分布聚集, 人类开发利用活动更加剧烈, 岛陆之间和海岛之间交通相对频繁, 也得到了诸多学者的关注和研究[13, 18-19]。相对而言, 北五岛距离大陆较远, 海岛分布较为分散, 航运交通不甚发达, 人类活动程度相对南五岛而言较低, 相关研究也鲜见报道。然而, 北五岛各岛面积大小不一, 轮廓形状各异, 海岛之间开发利用程度差别较大, 人类活动具有明显的空间分异性, 使得北五岛各岛之间和海岛内部景观格局差异的驱动因子更加复杂, 开展北五岛景观格局特征及其与生态系统服务功能的相互关系研究具有重要的意义和独特的优势。
(2) 人为干扰指数的建立
北五岛景观分类得出的8种景观类型中, 仅有草地和裸地属于海岛自然景观, 其余的黑松林、刺槐林、建筑用地、交通用地、其他硬化地面和农田均属于人工景观。人工景观对原生生态系统带来干扰但影响程度不一, 为了表征海岛受到人为干扰的程度及其空间特征, 本文提出了人为干扰指数(HII)指标, 在该指标的计算中, 不同人工景观影响系数(IF)的确定十分关键。交通用地不但在施工过程中深刻改变海岛地形地貌, 破坏生物栖息地, 在运营期还通过船舶、车辆排放污染物等方式对海岛带来持续的影响[9], 其影响程度最高, 影响系数取1;建筑用地和其他硬化地面的影响主要表现在施工期的改变海岛地表形态、破坏植被, 之后的影响相对较小[9], 影响系数取0.8;农田开垦往往将自然界的植物群落改造成大面积种植特定的农作物, 改变原生植物群落结构, 影响生物多样性[25], 影响系数取0.4;人工林虽然能够帮助维持海岛生态系统稳定性, 但实质上也是一种人为干扰, 可能会对原生植物群落构成威胁[26-27], 影响系数取0.2, 进而计算北五岛人为干扰程度及其空间分布特征。人为干扰指数考虑了不同人类活动的影响程度差异, 同时具有计算简便、数据可获取性强以及普适性等优点, 可为合理地开发利用海岛提供重要参考。
(3) 景观格局生态效应指标的选择
景观格局演变的生态效应涉及大气环境效应、水环境效应、生态系统服务效应、城市热岛效应等方面[1, 28-29]。对于海岛生态系统而言, 特别是本文研究的庙岛群岛北五岛, 人为影响下景观格局变化带来的污染物排放量总体较小, 海岛基本无地表径流, 因此引起的大气和水环境效应较小;海岛城镇建设能够造成地表温度的空间差异[30], 但北五岛城镇建设程度相对不高, 热岛效应也有限。实际上, 北五岛景观格局演变主要表现在自然景观面积不断萎缩且被割裂, 人工景观面积增大, 景观类型不断增多, 景观格局空间分异性增强, 进而对海岛生态系统产生影响。海岛NPP不仅能够直接反映植被群落的生产能力, 还是判定生态系统碳源/汇和调节生态过程的主要因子[31], 而且对不同开发利用类型响应灵敏[18];海岛植物多样性在维持和调控生态系统物质循环、能量流动和稳定性等方面具有重要的基础性作用[32-33], 同时也越来越受到人类活动的干扰[19];土壤作为海岛生态系统的基底, 不但为植物生长提供场所和养分, 还在有机质还原、元素循环中发挥着关键作用[34], 而景观格局变化对土壤生态过程构成重要影响, 不同景观类型条件下的土壤含水量、营养元素往往存在显著差异[35-36]。因此, 本文选择海岛NPP、植物多样性和土壤性质3方面进行景观格局的生态效应分析, 能够较为全面地反映景观格局影响下海岛生态系统的变化特征。
4.1.2 景观格局空间特征成因从景观尺度来看, 植被景观(针叶林、阔叶林和草地)构成了北五岛的景观基质, 其规模较大且分布广泛, 阔叶林多分布在坡脚, 往往连片分布, 连通性较好, 斑块密度较小;针叶林多在背坡、坡肩、坡顶等区域, 地形较为复杂, 既存在连片分布的情况, 又有较多的小斑块分布于其他植被景观中, 使得斑块密度较高;草地景观在北五岛整个区域分布广泛, 有的与针叶林、阔叶林毗邻, 有的在建筑用地中零星分布, 从而拥有着最高的斑块密度。另外, 植被生长受到复杂地形影响[37], 使得斑块轮廓较为曲折, 同时各种植被景观往往彼此相连, 边界绵长且复杂, 使得各类植被景观的边缘密度和平均形状指数均较大。建筑用地规模仅次于植被景观, 集中分布在海拔较低、坡度较小的区域, 在其他区域呈零星分布状态[19], 斑块密度较高, 其斑块形状规整, 边界平直, 平均形状指数处于最小值。道路用地是交通用地的主要组成部分, 其景观特征细长, 曲折蜿蜒地在海岛上分布, 从而拥有最大的平均形状指数, 但其规模有限, 且连通性较好, 因而斑块密度较低。裸地一部分为裸岩, 主要分布于海岛四周, 一部分位于海岛内部以裸土地的形式存在, 裸岩斑块形状细长, 边界较为平直, 裸土地单个斑块面积小, 形状不规则, 呈分散分布状态, 使得裸地各景观指标总体上处于中间位置。其他硬化地面和农田面积均较小, 仅在砣矶岛和大钦岛有分布, 斑块面积较小且形状规整, 二者各项景观格局指标均偏低。
从海岛尺度来看, 海岛景观格局一方面受到海岛自然特征的制约, 另一方面, 社会经济条件的差异也是重要的驱动因子。分析各岛景观格局与自然和人文因子的关系, 选取海岛面积(Area)、轮廓复杂度(ISI)、人口、生产总值(GDP)共4项因子(图 7) , 其中轮廓复杂度用海岛平均形状指数(Island Shape Index, ISI)表示, 其计算方法为:ISI=Length/[2×(π×Area)0.5], 海岛形状为圆形时, ISI为1, 形状越来越复杂和不规则, ISI逐渐增大[38]。海岛面积、人口和GDP与景观格局指数的关系表现出了相同的特征, 随着海岛面积、人口和GDP的增加, 斑块密度和人为干扰指数均明显增大, 其他指数变化特征不明显;海岛轮廓复杂度与各景观格局指数均未见明显关系。经相关分析可得, 海岛面积、人口和GDP三者之间两两呈显著正相关。可以发现, 海岛面积的增大一方面使得可容纳的景观类型和斑块数量增多, 另一方面也带来了海岛人口和GDP的显著增加;人口和GDP作为衡量海岛社会经济状况的基本指标, 直接带来人工景观的增多以及自然景观的缩减和变化。

|
| 图 7 各岛景观格局指数与影响因子的关系 Fig. 7 Relationship between landscape pattern indices and impact factors in different islands |
从区块尺度来看, 景观格局可能受到地形的制约。地形因子与光照、温度、水分等生态条件有密切的关系, 体现了对生境条件的综合性指示能力[39], 同时能够对人类活动构成制约, 影响景观格局的形成与变化[40]。提取各区块的平均海拔和坡度, 并与景观格局指数进行相关分析, 结果显示斑块密度、边缘密度和平均形状指数与海拔呈显著正相关, 与坡度无显著相关性, 人为干扰指数与海拔和坡度均呈显著负相关(表 6) 。结合上述分析, 海拔较低、坡度较小的区域往往连片分布着建筑用地, 其生态系统影响系数较高, 海拔较高、坡度较大的位置多分布着天然草地、裸地以及影响系数较小的人工林, 使得人为干扰指数与海拔和坡度呈负相关;海拔较高的区域除了植被景观外, 还分散分布着诸多建筑用地、交通用地和其他硬化地面小斑块, 加之植被景观本身斑块密度、边缘密度和平均形状指数较高, 使得这海拔与这3个景观格局指标呈显著正相关;较大的坡度对人类开发行为构成限制, 且随着坡度的增大, 裸地逐渐增多, 而裸地的各项景观指标均适中, 使得坡度与各指标的相关性不显著。不同景观格局指数之间也表现出了明显的相关性, 斑块密度、边缘密度和平均形状指数三者之间两两呈显著正相关, 人为干扰指数与斑块密度和边缘密度呈显著正相关, 与平均形状指数未见显著相关性, 这是由于人类活动直接带来了斑块数量和边缘长度的增加, 因为构建了形状复杂的人工林和形状规整的建筑用地、交通用地, 使得其与平均形状指数相关性不显著。可以发现, 地形对海岛景观格局特征构成了明显的影响。
| 项目 Items | PD | ED | MSI | HII |
| 海拔 Altitude | 0.314** | 0.420** | 0.081* | -0.179** |
| PD | 1 | |||
| MSI | 0.264** | 0.316** | 1 | |
| 项目 Items | PD | ED | MSI | HII |
| 坡度 Slope | -0.048 | 0.042 | 0.031 | -0.482** |
| ED | 0.904** | 1 | ||
| HII | 0.168** | 0.182** | 0.013 | 1 |
综上, 海岛面积是北五岛景观格局的基本因子, 面积大小直接决定了其能够承载的人口和经济规模;地形是景观格局的重要限制因子, 地形能够对人类活动和植被空间分布进行限制;人类活动是景观格局变化的主要驱动因子, 使得不同景观类型具有鲜明的格局特征, 不同海岛和不同区块的景观格局具有明显的空间异质性。
4.1.3 景观格局生态效应探析就景观尺度而言, 针叶林的NPP、多样性和土壤水肥均处于较高位置, 阔叶林除了多样性偏低外, 其余指标也均较高, 这说明人工林在固碳、提升土壤质量方面发挥着重要作用, 这与王晓丽等[41]和马志敏等[36]的研究结果相一致;现场调查发现, 北五岛阔叶林下草本植物多为单一优势种如中华隐子草、披针叶苔草等, 盖度较高, 生物量丰富, 但多样性较低;针叶林下草本植物生物量相对较小, 但种类丰富多样, 多样性指数较高[19]。草地的多样性和土壤水肥指标值也较高, 这一方面说明海岛原生植物群落拥有较为丰富的多样性, 另一方面也表明了草地在保水保肥方面具有一定优势, 这与高君亮等[42]的研究结果相一致, 但由于个体特征, 草地NPP处于中间位置。农田作为人工干预较强的植被景观, 其物种单一, 且北五岛农田开垦总量较小, 基本无农田土壤增肥措施, 使得农田生物多样性和土壤指标均偏低, 这与尤民生等[25]与李东等[43]的研究结果相呼应。解译精度和海岛实际情况使得非植被景观中混有部分植被, 因此裸地、建筑用地、交通用地和其他硬化地面也具有一定的生态效应指标值, 但总体处于较低位置。可以发现, 人工林景观总体上带来了正向的生态效应, 这表明海岛人工林种植取得了明显的成效, 也说明了黑松和刺槐是海岛人工林种植的理想树种[18]。同时, 城乡建设、农田开垦、道路建造等人类活动不可避免地会对海岛生态系统构成威胁, 控制开发利用规模、优化建设布局与改进建造方式是减少其负面影响的重要措施。
就海岛尺度而言, 景观格局指数与生态效应指标明显的相关关系仅见于Pielou指数与边缘密度、土壤肥力与边缘密度, 其他的相互关系均不显著, 海岛尺度上生态效应指标对景观格局的响应较不灵敏, 这与海岛面积差异以及各岛生态系统特殊状况有关。地表覆盖类型及植被生长状况是海岛NPP最主要的影响因子[18], 各岛的植被景观占比总体上差异不大, 而代表植被生长状况的NDVI值却表现出了显著的差异, 砣矶岛和北隍城岛较高, 小钦岛居中, 大钦岛和南隍城岛则明显偏低。结合现场调查可知, 砣矶岛和北隍城岛植被景观连片分布特征更加明显, 景观内部区域受到干扰较小, 植被较为茂密, 长势良好;大钦岛人为干扰最为强烈, 尤其是海带晒场的建造对植被的规模和质量均造成了威胁, 南隍城岛的牛羊放牧也对植被生长带来压力。经分析各岛的NPP平均密度与NDVI表现出了明显的相关性(R2=0.70) , 这表明各岛的NPP更多地受到植被生长状况的影响, 使得其与景观格局的关系不显著。多样性指标在南隍城岛相对较高, 而在北隍城岛最低, 这可能是由于南隍城岛对外和内部的交通更为频繁, 带来了物种多样性的增加, 而北隍城岛作为庙岛群岛最北端的海岛, 交通不甚发达, 同时其裸地面积过大, 造成海岛整体多样性水平较低。土壤含水量在小钦岛远低于其余四岛, 可能由于小钦岛面积最小, 汇水蓄水能力差;土壤养分在北隍城岛最高, 在南隍城岛则最低, 这可能与海岛植被生长状况有关。
就区块尺度而言, NPP平均密度与各景观格局指数的简单相关指数具有显著性, 但以植被景观为控制变量的偏相关性明显减弱。植被景观往往拥有较高的NPP平均密度, 建筑用地、交通用地、裸地和其他硬化地面则相反, 植被景观总体上斑块密度、边缘密度和平均形状指数较高, 人为干扰指数较低, 再加上该尺度的评价单元具相同的面积, 且忽略了不同海岛的差异, 使得区块尺度上NPP与景观格局指数表现出明显的简单相关性;偏相关性的不显著说明了NPP平均密度对于景观格局破碎度、边缘效应和形状复杂性的响应不灵敏。多样性指标中, Shannon-Wiener指数与斑块密度和边缘密度呈显著负相关, 且偏相关性比简单相关性更强, 景观破碎化对生物多样性的威胁已经得到诸多学者的研究证实[3, 44-45], 边缘效应同样是生物多样性的重要影响因素[46-47], 北五岛植物多样性除了受到景观类型的影响之外, 景观格局的破碎度和边缘效应产生了更加重要的作用。Pielou指数与景观格局指数的简单相关分析和偏相关分析结果均不显著, 表明了景观格局破碎度、边缘效应和形状复杂性对群落物种的均匀度影响较小。土壤指标中, 土壤含水量与景观格局指数的关系与NPP指标表现出相同的特征, 这是由于土壤含水量的空间分异主要受到景观类型分布特征的影响[48], 与景观格局的破碎化、边缘效应等关系不明显;与之相反, 土壤养分与景观格局指数的简单相关性不明显, 但剔除了植被景观影响后表现出了显著的负相关性, 土壤养分的分布和迁移受到景观镶嵌格局的制约[48], 景观格局的破碎度和边缘效应对北五岛土壤养分带来了明显的影响。值得注意的是, 人为干扰指数与生态效应指标的简单相关性基本为显著负相关, 但偏相关性出现了正相关的情形。这可能是由于人为干扰指数本身就是由各类景观类型规模经计算得出, 以植被景观为控制变量的偏相关分析的意义不明显, 而简单相关分析结果能够更准确地反映人类活动对海岛生态系统的影响, 人为干扰强度的增大相应地带来了各项生态效应指标的降低。
格局和过程始终是生态学中极富挑战性的课题[49], 尺度也是景观生态学研究中的重要因子[50], 不同尺度的选择对景观格局分析结果及其生态效应具有重要影响[51-53]。本文的研究结果显示不同尺度景观格局的生态效应具有差别, 能够为全面揭示海岛人类活动影响下自然生态系统的变化特征提供重要参考。在下一步的工作中, 应当进一步开展不同尺度上海岛生态系统特征及影响因子的研究。
4.2 结论本文以庙岛群岛北五岛为研究区, 基于现场调查和3S技术, 从景观尺度、海岛尺度和区块尺度分析景观格局空间特征, 进而探讨海岛景观格局与NPP、植物多样性和土壤性质的关系。主要结论如下:
(1) 北五岛景观尺度、海岛尺度和区块尺度的景观格局均表现出了空间差异在景观尺度上, 针叶林、阔叶林和草地3类植被景观面积最大, 斑块密度、边缘密度和平均形状指数总体较高, 建筑用地也具有较大规模, 其斑块密度较高, 平均形状指数处于最低值, 裸地也具有一定规模, 其各项景观格局指标处于中间位置, 其他景观类型面积相对较小。在海岛尺度上, 随着海岛面积、人口和GDP的增加, 斑块密度和人为干扰指数均明显增大, 其他指数变化特征不明显;海岛轮廓复杂度与各景观格局指数均未见明显关系。在区块尺度上, 斑块密度、边缘密度和平均形状指数与海拔呈显著正相关, 人为干扰指数与海拔和坡度均呈显著负相关;不同景观格局指数之间也表现出了明显的相关性, 斑块密度、边缘密度和平均形状指数三者之间两两呈显著正相关, 人为干扰指数与斑块密度和边缘密度呈显著正相关。
(2) 海岛面积是北五岛景观格局的基本因子面积大小直接决定了其能够承载的人口和经济规模;地形是景观格局的重要限制因子, 地形能够对人类活动和植被空间分布进行限制;人类活动是景观格局变化的直接驱动因子, 使得不同景观类型具有鲜明的格局特征, 不同海岛和不同区块的景观格局具有明显的空间异质性。
(3) 海岛景观格局的生态效应与尺度密切相关景观尺度上, 各项生态效应指标在不同景观类型上均具有显著差异, 在针叶林、阔叶林、草地中相对较高;海岛尺度上的生态效应指标对景观格局的响应不甚灵敏;区块尺度上, 生态效应指标与景观格局指数表现出了显著的简单相关性和偏相关性, 但二者结果具有差异。NPP和土壤水分主要受到景观类型和植被生长状况的影响, 多样性和土壤养分同时受到景观类型和景观格局破碎度、边缘效应的影响, 人为干扰强度的增大带来了各项生态效应指标的降低。控制建设规模、优化景观布局与改进开发利用方式是维系海岛生态系统稳定性的重要措施。
| [1] | 陈利顶, 孙然好, 刘海莲. 城市景观格局演变的生态环境效应研究进展. 生态学报, 2013, 33(4): 1042–1050. |
| [2] | Cook E A. Landscape structure indices for assessing urban ecological networks. Landscape and Urban Planning, 2002, 58(24): 269–280. |
| [3] | Ramalho C E, Laliberté E, Poot P, Hobbs R J. Complex effects of fragmentation on remnant woodland plant communities of a rapidly urbanizing biodiversity hotspot. Ecology, 2014, 95(9): 2466–2478. DOI:10.1890/13-1239.1 |
| [4] | Strohbach M W, Haase D. Above-ground carbon storage by urban trees in Leipzig, Germany. Landscape and Urban Planning, 2012, 104(1): 95–104. DOI:10.1016/j.landurbplan.2011.10.001 |
| [5] | 李锋, 叶亚平, 宋博文, 王如松. 城市生态用地的空间结构及其生态系统服务动态演变——以常州市为例. 生态学报, 2011, 31(19): 5623–5631. |
| [6] | 俞孔坚, 乔青, 李迪华, 袁弘, 王思思. 基于景观安全格局分析的生态用地研究——以北京市东三乡为例. 应用生态学报, 2009, 20(8): 1932–1939. |
| [7] | 齐连明, 张祥国, 李晓. 国内外海岛保护与利用政策比较研究. 2013. |
| [8] | Jupiter S, Mangubhai S, Kingsford R T. Conservation of biodiversity in the Pacific islands of Oceania:challenges and Opportunities Pacific. Pacific Conservation Biology, 2014, 20(2): 206–220. DOI:10.1071/PC140206 |
| [9] | 池源, 石洪华, 郭振, 丁德文. 海岛生态脆弱性的内涵、特征及成因探析. 海洋学报, 2015, 37(12): 93–105. |
| [10] | Qie L, Lee T M, Sodhi N S, Lim S L H. Dung beetle assemblages on tropical land-bridge islands:small island effect and vulnerable species. Journal of Biogeography, 2011, 38(4): 792–804. DOI:10.1111/jbi.2011.38.issue-4 |
| [11] | Katovai E, Burley A L, Mayfield M M. Understory plant species and functional diversity in the degraded wet tropical forests of Kolombangara Island, Solomon Islands. Biological Conservation, 2012, 145(1): 214–224. DOI:10.1016/j.biocon.2011.11.008 |
| [12] | 王晓丽, 王嫒, 石洪华, 郑伟, 周然. 海岛陆地生态系统固碳估算方法. 生态学报, 2014, 34(1): 88–96. |
| [13] | 王常颖, 张杰, 辛红梅, 付军. 基于景观格局的海岛开发潜在生态风险评价. 生态学报, 2008, 28(6): 2811–2817. |
| [14] | 林明太, 孙虎, 郭斌. 福建旅游型海岛景观格局变化——以妈祖圣地湄洲岛为例. 生态学杂志, 2010, 29(7): 1414–1419. |
| [15] | Tzanopoulos J, Vogiatzakis I N. Processes and patterns of landscape change on a small Aegean island:the case of Sifnos, Greece. Landscape and Urban Planning, 2011, 99(1): 58–64. DOI:10.1016/j.landurbplan.2010.08.014 |
| [16] | 池源, 石洪华, 丰爱平. 典型海岛景观生态网络构建——以崇明岛为例. 海洋环境科学, 2015, 34(3): 433–440. |
| [17] | 孙永光, 康婧, 王伟伟, 王传珺, 付元宾, 李培英, 索安宁. 海岛植被景观异质性空间特征对地形变化的响应——以大长山岛为例. 生态学杂志, 2015, 34(6): 1705–1712. |
| [18] | 池源, 石洪华, 王晓丽, 李捷, 丰爱平. 庙岛群岛南五岛生态系统净初级生产力空间分布及其影响因子. 生态学报, 2015, 35(24): 8094–8106. |
| [19] | Chi Y, Shi H H, Wang X L, Qin X B, Zheng W, Peng S T. Impact factors identification of spatial heterogeneity of herbaceous plant diversity on five southern islands of Miaodao Archipelago in North China. Chinese Journal of Oceanology and Limnology, 2016. |
| [20] | Vogiatzakis I, Pungetti G, Mannion A. Mediterranean Island Landscapes:Natural and Cultural Approaches. Springer, 2008. |
| [21] | J B, R E. The Theory of Island Biogeography Revisited. Princeton: Princeton University Press, 2010. |
| [22] | Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production:a process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811–841. DOI:10.1029/93GB02725 |
| [23] | 张金屯. 数量生态学. 北京: 科学出版社, 2004. |
| [24] | 马克平, 刘玉明. 生物群落多样性的测度方法:Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231–239. |
| [25] | 尤民生, 刘雨芳, 侯有明. 农田生物多样性与害虫综合治理. 生态学报, 2004, 24(1): 117–122. |
| [26] | Rosoman G. The Plantation Effect. Greenpeace New Zealand, 1994: 48. |
| [27] | Michelsen O, McDevitt J E, Coelho C R V. A comparison of three methods to assess land use impacts on biodiversity in a case study of forestry plantations in New Zealand. The International Journal of Life Cycle Assessment, 2014, 19: 1214–1225. DOI:10.1007/s11367-014-0742-1 |
| [28] | Pauleit S, Ennos R, Golding Y. Modeling the environmental impacts of urban land use and land cover change-a study in Merseyside, UK. Landscape Urban Planning, 2005, 71(2-4): 295–310. DOI:10.1016/S0169-2046(04)00083-0 |
| [29] | Lovell S T, Johnston D M. Creating multifunctional landscapes:how can the field of ecology inform the design of the landscape?. Frontiers in Ecology and the Environment, 2009, 7(4): 212–220. DOI:10.1890/070178 |
| [30] | 池源, 石洪华, 王晓丽, 丰爱平. 庙岛群岛南五岛地表温度时空特征及影响因子. 生态学杂志, 2015, 34(8): 2309–2319. |
| [31] | Field C B, Behrenfeld M J, Randerson J T, Falkowski P. Primary production of the biosphere:integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237–240. DOI:10.1126/science.281.5374.237 |
| [32] | Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441: 629–632. DOI:10.1038/nature04742 |
| [33] | 马克平. 生物多样性与生态系统功能的实验研究. 生物多样性, 2013, 21(3): 247–248. |
| [34] | 欧阳志云, 王如松, 赵景柱. 生态系统服务功能及其生态经济价值评价. 应用生态学报, 1999, 10(5): 635–640. |
| [35] | 陈浮, 濮励杰, 彭补拙, 包浩生. 新疆库尔勒市土地利用变化对土壤性状的影响研究. 生态学报, 2001, 21(8): 1290–1295. |
| [36] | 马志敏, 吕一河, 孙飞翔, 王江磊. 黑河中游荒漠绿洲区土地利用的土壤养分效应. 生态学报, 2013, 33(19): 6328–6334. |
| [37] | 石洪华, 王晓丽, 王嫒, 刘振英, 麻德明. 北长山岛森林乔木层碳储量及其影响因子. 生态学报, 2013, 33(19): 6363–6372. |
| [38] | 彭思羿, 胡广, 于明坚. 千岛湖岛屿维管β植物多样性及其影响因素. 生态学报, 2014, 34(14): 3866–3872. |
| [39] | Currie J, Paquin V. Large-scale biogeographical patterns of species richness of trees. Nature, 1987, 329: 326–327. DOI:10.1038/329326a0 |
| [40] | 郭洪峰, 许月卿, 吴艳芳. 基于地形梯度的土地利用格局与时空变化分析——以北京市平谷区为例. 经济地理, 2013, 33(1): 160–166. |
| [41] | 王晓丽, 王媛, 石洪华, 彭士涛, 宫立鹏, 覃雪波. 山东省长岛县南长山岛黑松和刺槐人工林的碳储量. 应用生态学报, 2013, 24(5): 1263–1268. |
| [42] | 高君亮, 罗凤敏, 高永, 原伟杰, 王淮亮, 党晓宏. 阴山北麓不同土地利用类型土壤养分特征分析与评价. 草业学报, 2016, 25(4): 230–238. |
| [43] | 李东, 王子芳, 郑杰炳, 高明. 紫色丘陵区不同土地利用方式下土壤有机质和全量氮磷钾含量状况. 土壤通报, 2009(2): 310–314. |
| [44] | Opdam P, Wascher D. Climate change meets habitat fragmentation:linking landscape and biogeographical scale levels in research and conservation. Biological Conservation, 2004, 117(3): 285–297. DOI:10.1016/j.biocon.2003.12.008 |
| [45] | Wood E M, Pidgeon A M, Radeloff V C, Helmers D, Culbert P D, Keuler N S, Flather C H. Housing development erodes avian community structure in U.S. protected areas. Ecological Applications, 2014, 24():1445-1462. |
| [46] | Young A, Mitchell N. Microclimate and vegetation edge effects in a fragmented podocarp-broadleaf forest in New Zealand. Biological Conservation, 1994, 67(1): 63–72. DOI:10.1016/0006-3207(94)90010-8 |
| [47] | Conway A J, Danby R K. Recent advance of forest-grassland ecotones in southwestern Yukon. Canadian Journal of Forest Research, 2014, 44(5): 509–520. DOI:10.1139/cjfr-2013-0429 |
| [48] | 傅伯杰, 陈利顶, 王军, 孟庆华, 赵文武. 土地利用结构与生态过程. 第四纪研究, 2003, 23(3): 247–255. |
| [49] | Holling C S. Cross-scale morphology, geometry and dynamics of ecosystems. Ecological Monographs, 1992, 62: 447–502. DOI:10.2307/2937313 |
| [50] | Levin S A. The problem of pattern and scale in ecology. Ecology, 1992, 73: 1943–1967. DOI:10.2307/1941447 |
| [51] | Wu J G, Loucks O L. From balance of nature to hierarchical patch dynamics:a paradigm shift in ecology. The Quarterly Review of Biology, 1995, 70(4): 439–466. DOI:10.1086/419172 |
| [52] | 崔步礼, 常学礼, 左登华, 宋芙蓉. 沙地景观中矢量数据栅格化方法及尺度效应. 生态学报, 2009, 29(5): 2463–2472. |
| [53] | 王德智, 邱彭华, 方源敏. 丽香铁路建设对沿线景观格局影响的尺度效应及其生态风险. 应用生态学报, 2015, 26(8): 2493–2503. |