 2017, Vol. 37
2017, Vol. 37文章信息
- 张睿洋, 王忠武, 韩国栋, 潘占磊, 刘芳, 武倩, 阿木尔萨那
- ZHANG Ruiyang, WANG Zhongwu, HAN Guodong, PAN Zhanlei, LIU Fang, WU Qian, AMUER Sana.
- 短花针茅荒漠草原α多样性对绵羊载畜率的响应
- The response of plant alpha diversity to different grazer stocking rate in a Stipa breviflora. desert steppe
- 生态学报. 2017, 37(3): 906-914
- Acta Ecologica Sinica. 2017, 37(3): 906-914
- http://dx.doi.org/10.5846/stxb201508191729
-
文章历史
- 收稿日期: 2015-08-19
- 网络出版日期: 2016-06-13





目前地球上的物种正以前所未有的速度灭绝, 物种丧失将会给人类生存和社会发展带来严重的威胁[1], 生存问题已从人类的范畴扩展到地球上相互依存的所有物种之间, 因此生物多样性的研究成为当今生态学界的热点[2]。草地生态系统作为陆地生态系统的重要组成部分, 为人类提供物质生产、水土保持、调节气候等多种功能和服务, 同时也是人类活动干扰最为严重的区域[3-4]。放牧活动是人类影响天然草原进化演替的主要驱动力[5-6]。许多学者的研究表明, 家畜的采食行为会对草地生态系统的结构和功能产生影响[7-9]。但关于草地植物多样性对放牧的响应机制仍存在争议。大量研究表明, 植物多样性与放牧强度间的关系可由经典的“中度干扰假说”观点解释[10-12], 即中等程度的干扰能维持较高的多样性[13-14]。但也有研究认为放牧对草地植物多样性的影响不仅取决于水分生态类型的多样性[15], 而且季节性的波动也会影响二者之间的关系[16]。
生物多样性在不同空间尺度下表征, α多样性为其中之一。α多样性即单位面积的物种数目, 主要指局部区域均匀生境中的物种数量, 也被称为生境内的多样性, 是对群落内物种数量及其相对多度的一种测量, 它反映了群落内物种通过资源竞争而形成的物种共存关系[17]。在对植物群落α多样性特征参数的研究中, 取样面积与取样数目的问题一直受到广泛关注[18-19], 但积累样方法却在研究工作中被广泛应用[20]。Magurran认为积累样方法可为确定最小取样强度提供很好的指示, 并有利于物种多度分布的研究[21]。目前, 国内关于草地α多样性的研究多集中于典型草原和草甸草原[22], 但载畜率对荒漠草原α多样性的作用机制, 以及尺度效应对α多样性的影响, 并以此确定最佳取样面积的研究仍然处于缺乏状态。
荒漠草原处在一个生态环境脆弱, 群落生产力较低, 稳定性较差, 对自然和人类活动的干扰较为敏感的地带, 且其状态转化的恢复力阈值很小, 作为荒漠草原的典型代表, 短花针茅荒漠草原在草地生态系统中占据着极特殊的位置。本研究以短花针茅荒漠草原为研究对象, 在连续11a固定载畜率的样地上, 通过测定不同绵羊载畜率下草地α多样性以及物种组成的变化, 采用巢式样方法确定荒漠草原的最佳取样面积, 探讨荒漠草原生态系统α多样性对不同载畜率以及取样尺度的响应机制, 为草地生物多样性保护和草地的可持续发展提供理论依据。
1 试验地概况试验地位于内蒙古自治区乌兰察布市四子王旗王府一队(41°47′17″N, 111°53′46″E), 海拔高度1450 m, 属中温带大陆性季风气候区, 年平均气温3.4 ℃, 年平均降水量299.4 mm, 年蒸发量约2300 mm, 无霜期90-120 d, 平均日照时数为3117.7 h;土壤为淡栗钙土。
试验区属短花针茅荒漠草原地带性植被, 草地类型为短花针茅+冷蒿+无芒隐子草。植被草层低矮, 平均高度为8 cm, 且植被较稀疏, 盖度为17%-20%, 种类组成较贫乏。建群种为短花针茅(Stipa breviflora), 优势种为冷蒿(Artemisia frigida)、无芒隐子草(Cleistogenes songorica)。主要伴生种有银灰旋花(Convolvulus ammannii)、阿尔泰狗娃花(Heteropappus altaicus)、栉叶蒿(Neopallasia pectinata)、木地肤(Kochia prostrata)、狭叶锦鸡儿(Caragana stenophylla)、羊草(Leymus chinensis) 和米氏冰草(Agropyron cristatum) 等。
2 研究方法 2.1 试验地设置放牧试验从2004年6月开始, 围封约50 hm2天然草地。试验设为随机区组试验, 将试验小区分成3个区组, 每个区组内设有4个载畜率水平, 分别为对照样地(Control Check, CK)、低水平载畜率(Light Grazing, LG):0.91羊单位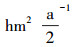
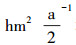
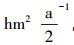
采用巢式样方法, 每个处理小区内设置3个巢式样方, 每个巢氏样方分别包含0.01、0.02、0.04、0.08、0.16、0.32、0.64、1、2.5、5、10、20、40、80、160、320、650 m217个样方(图 1), 并统计各级样方内的物种数和植物多度。

|
| 图 1 样方示意图 Fig. 1 The Quadrat schematic |
采用了三类α多样性指数, 即丰富度指数(richness index)、均匀度指数(evenness index) 和多样性指数(diversity index)。
(1) 物种丰富度指数采用Margalef指数:
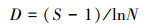
|
(2) 多样性指数采用Shannon-Wiener指数:
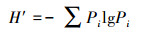
|
(3) 优势度指数采用Simpson指数:
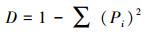
|
(4) 均匀度指数采用Pielou指数:
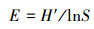
|
式中, S为物种数目;N为所有物种个体总数;Pi是一个个体属于第i种的概率, 在此用相对多度表示。
2.4 数据处理采用Excel 2007进行数据统计, SAS 9.1软件包对相关数据进行方差分析。回归分析采用对数模型y=aln (x)+b, 其中自变量x为取样面积, 因变量y为物种数或α多样性指数, a和b作为模型参数分别代表每增加一个对数单位时因变量的增长速率和单位面积为1 m2时的物种数或α多样性指数。
3 结果与分析 3.1 物种组成分析在不同载畜率水平下, 各处理之间物种组成随载畜率的升高呈减少的趋势(表 1), 对照样地共有15个物种, 相比之下LG样地少了4种植物, 分别为:狭叶锦鸡儿(Calagana stenophylla)、小叶锦鸡儿(Calagana microphylla)、兔唇花(Lagochilus ilicifolius)、栉叶蒿(Nsaleopallasia pectinata)。MG样地比起LG样地则少了羊草(Leymus Chinensis) 和阿尔泰狗娃花(Heteropappus altaicus) 两种植物, 而HG样地仅比MG样地减少了一种植物二裂委陵菜(Potentilla bifurca)。但是这些物种绝大部分在群落中所占比例并不大, 个体数量很少。通过计算物种的相对多度, 占群落95%以上的主要集中在8个物种上, 即相对多度占群落10%以上的短花针茅(Stipa breviflora)、冷蒿(Artemisia frigida)、无芒隐子草(Cleistogenes songorica)、阿氏旋花(Euphorbia humifusa), 和10%以下的克氏针茅(Stipa kryloviiRoshev)、糙隐子草(Cleistogenes squarrosa)、阿尔泰狗娃花(Heteropappus altaicus)、木地肤(Kochia prostrata)。
| 物种分类 Classification of Species |
物种名称 Species name |
载畜率水平 Level of stocking rate |
|||
| CK | LG | MG | HG | ||
| 优势种 | 短花针茅Stipa breviflora | ● | ● | ● | ● |
| Dominant | 冷蒿Artemisia frigida | ● | ● | ● | ● |
| Species | 无芒隐子草Cleistogenes songorica | ● | ● | ● | ● |
| 非优势种 | 阿氏旋花Euphorbia humifusa | ● | ● | ● | ● |
| Common Species | 木地肤Kochia prostrata | ● | ● | ● | ● |
| 细叶葱Astragalum galactites | ● | ● | ● | ● | |
| 克氏针茅Stipa kryloviiRoshev | ● | ● | ● | ● | |
| 糙隐子草Cleistogenes squarrosa | ● | ● | ● | ● | |
| 二裂委陵菜Potentilla bifurca | ● | ● | ● | ||
| 羊草Leymus Chinensis | ● | ● | |||
| 阿尔泰狗娃花Heteropappus altaicus | ● | ● | |||
| 狭叶锦鸡儿Calagana stenophylla | ● | ||||
| 小叶锦鸡儿Calagana microphylla | ● | ||||
| 兔唇花Lagochilus ilicifolius | ● | ||||
| 栉叶蒿Nsaleopallasia pectinata | ● | ||||
8个主要物种中, 在MG、HG载畜率水平下作为优势种的短花针茅和无芒隐子草的相对多度显著增加(P < 0.05), 而糙隐子草的相对多度显著减少(P < 0.05)。阿尔泰狗娃花在LG、MG、HG放牧处理下均比对照CK显著减少(P < 0.05)。同样, 木地肤的相对多度也随着载畜率的升高而下降, 其中对照CK和处理LG、MG、HG以及LG和HG之间差异显著(P < 0.05), 而LG和MG、MG和HG之间则没有显著性差异(P>0.05)。而冷蒿、阿氏旋花、克氏针茅的相对多度则对载畜率的升高没有显著性的变化(图 2)。

|
| 图 2 不同载畜率下主要物种的相对多度(P < 0.05) Fig. 2 The relative density of major species in different stocking rate |
4个载畜率水平下物种数目均随取样面积的增大而增加, 取样面积从0.01 m2到40 m2的尺度上物种数目快速增加, 40 m2以后增加速度趋于平缓(图 3), 而进入平衡态的这一点应为最小取样数。通过回归分析可以看出, 不同载畜率下物种数目的增加符合对数增长模型:y=aln (x)+b, 实际观测值和拟合结果基本一致。

|
| 图 3 种-面积关系曲线图 Fig. 3 The relationship between number of species and sample area |
取样面积在0.01 m2到0.08 m2时, 各处理间无显著性差异(P>0.05);在0.16 m2到80 m2时, CK、LG极显著高于MG和HG (P < 0.01), 而CK和LG、MG和HG之间则无显著差异(P>0.05)。在160 m2以后的尺度上HG开始极显著低于其他处理(P < 0.01), 650 m2时LG和CK之间出现极显著差异(P < 0.01)。
3.3 α多样性分析由Margalef丰富度指数、Shanon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数的回归分析可知(图 4-图 7), 对数模型y=aln (x)+b能够较好的反映α多样性各类指数与取样面积之间的关系。Pielou均匀度指数随着取样面积的增加而减小, 在对数模型中参数a为负值。其余指数均显示出随着样方面积的扩大而增大的趋势。在曲线进入平衡态之前, α多样性指数明显受取样尺度的影响, 样方面积越大多样性指数的变化速率越大。在不同载畜率处理下各指数和取样面积关系曲线的拐点均出现在40 m2左右的尺度上, 之后多样性指数值不再明显增加或减少, 曲线逐渐趋于缓和。

|
| 图 4 Margalef丰富度指数-取样面积关系曲线图 Fig. 4 The relationship between Margalef indices and sample size |

|
| 图 5 Shanon-Wiener指数-取样面积关系曲线 Fig. 5 The relationship between Shanon-Wiener indices and sample size |

|
| 图 6 Simpson指数-取样面积关系曲线图 Fig. 6 The relationship between Simpson indices and sample size |

|
| 图 7 Pielou指数-取样面积关系曲线图 Fig. 7 The relationship between Pielou indices and sample size |
Margalef指数的分析表明(图 4), 当取样面积在0.16 m2时, 处理MG、HG和对照CK开始出现极显著性差异(P < 0.01), 0.64 m2下处理LG极显著高于MG、HG (P < 0.01)。而在达到160 m2后处理HG开始与其他处理出现极显著差异(P < 0.01)。
Shanon-Wiener多样性指数在处理HG与对照CK在0.16 m2的尺度上开始出现显著性差异(P < 0.05), 达到0.32 m2后处理MG、HG和LG、CK出现极显著差异(P < 0.01), 80 m2时HG极显著低于其他处理(P < 0.01)。而在达到160 m2后, 4个载畜率水平之间均存在极显著差异(P < 0.01)(图 5)。
Simpson优势度指数在0.01 m2尺度上处理HG出现了显著差异(P < 0.05), 但由于取样面积过小不足以说明问题, 处理HG与CK、LG之间在0.32 m2尺度上出现极显著差异, 在160 m2后与各处理间均出现极显著差异(P < 0.01), 而处理MG在2.5 m2后开始显著低于对照CK (P < 0.01)。直到650 m2时处理LG与CK出现极显著差异(P < 0.01)(图 6)。
Pielou均匀度在0.04 m2的尺度上, 处理MG、HG和对照CK开始出现显著差异(P < 0.05), 在0.32 m2到2.5 m2之间LG显著大于MG、HG (P < 0.05), 达到5 m2以后处理HG显著低于CK、LG。在320 m2时CK和LG处理之间出现极显著性差异(P < 0.01)。而从整体看LG与MG、MG与HG之间并没有明显的差异性(图 7)。
4 讨论在现代生态学领域内, 对群落物种多样性的研究首先面临的问题就是确定取样面积的大小和数目, 即种-面积关系, 学者们对此问题进行了大量的研究讨论。在一定尺度范围内, 各群落的物种数目均随取样面积的增加而增加, 这是由于受到环境和资源异质性的影响, 随着面积的扩大, 出现的斑块效应导致物种增加[23]。也有研究认为群落物种数目同时受取样面积和样方形状的影响, 由于边缘效应长方形样方可以获得与面积加倍的正方形样方相近的取样效果[24], 因此采用巢式样方是研究种-面积关系的适宜方法。在本研究中, 物种数量曲线在取样面积较高水平40 m2时才进入平衡点, 与白永飞和杨利民等人在贝加尔针茅、克氏针茅、大针茅草原的研究结论不一致[24-25]。一方面, 由于不同尺度样地内的微地形条件、土层厚度变化与灌木丛导致的水分富集效应使荒漠草原土壤水分呈现不同程度斑块分布, 变异系数随取样面积的增加呈先增大, 后略减小至趋于平稳的趋势, 而随着试验取样尺度的不断增大, 样地土壤厚度不同与地面凹凸所造成的土壤斑块格局的控制作用被弱化[26-27]。另一方面, 家畜的排泄与踩踏使地表土壤结构与土壤养分发生改变, 而土壤的异质性决定了植物空间格局的异质性, 因此放牧对植物种群空间分布的影响比较明显, 刘红梅等人研究指出短花针茅荒漠草原在放牧条件下土壤碳氮养分与植物群落受随机因素引起的空间变异较大, 最佳的采样尺度应为更大的釆样幅度和更小的采样间距[28-29]。因此在较大的取样面积尺度上才能反映出生境和资源的多样性, 从而体现物种数目的增长曲线。通过回归分析, 对数模型:S=alnA+b能够较好地反映草原群落的种-面积关系和α多样性指数, 模型的拟合结果基本接近于实际观测结果, 这和白永飞的研究结论一致[25]。
据杨利民的研究结果表明, 用多度参数和生物量参数计算的多样性指数值虽有差异, 但进入平衡态的位置基本一致[23]。考虑到短花针茅荒漠草原需要大面积尺度的取样, 本研究采用较为便捷的物种多度参数计算多样性指数。结果显示多样性指数明显受取样面积大小的影响, 在曲线进入平衡点之前, 多样性指数的变化速率随样方面积的增大而增大。而在不同载畜率水平下对多样性指数的分析表明, 短花针茅草原群落的α多样性随载畜率的增加呈降低的趋势, 这一研究结果与中度干扰假说不一致, 但符合Milchunas的研究观点, 即半干旱草地植物多样性对放牧的响应呈负相关的研究结论[8]。对于干旱的荒漠草原来说, 种间竞争主要体现在水分竞争上, 放牧降低了群落覆盖度, 增加了土壤含水量的蒸发, 从而抑制了受水分胁迫物种的生长, 使植物多样性呈现随载畜率增加而降低的趋势[22, 30]。
从群落物种组成来看, 放牧引起的α多样性降低, 主要是沿载畜率梯度, 由于非优势物种的相对多度降低, 优势种短花针茅、无芒隐子草相对多度增加。表明短花针茅、无芒隐子草对放牧具有较高的忍耐性和适应能力。而非优势种对群落α多样性降低的贡献较大, 对放牧的响应更为敏感。这主要是因为大部分非优势种的适口性较好, 在轻度放牧条件下就已经受绵羊的选择性采食而开始减少甚至消失。然而, 随着放牧强度的进一步增加, 绵羊对不同物种的选择性降低, 啃食强度增加, 导致主要常见物种的多度也逐渐降低。有研究认为以生物量为参数计算多样性指数时, 由于优势种具有较高的生物量, 因此比非优势种对生态系统功能有更多的主导作用[17]。本研究以物种多度参数计算的多样性指数显示出草地生态系统中非优势物种对生态系统同样有着重要作用, 当放牧干扰时这些物种比优势种更加脆弱、容易受到破坏, 能够反映出物种对载畜率的响应机制以及对资源改变的适应策略的多样性, 因此是否拥有大量非优势种对多样性是一项重要指标。
5 结论通过上述分析和讨论, 得出以下结论:在短花针茅荒漠草原对物种数以及α多样性调查的最佳取样面积为40 m2, 种-面积关系以及α多样性指数-面积关系都符合对数增长模型:y=aln (x)+b。不同载畜率间对α多样性指数的分析受取样面积影响, 随着面积尺度的增加, 多样性指数沿载畜率梯度差异性逐渐增大。取样面积为0.16 m2到0.64 m2小尺度上可以体现α多样性指数在中、高载畜率和零、低载畜率水平间的差异性, 而α多样性指数在取样面积为160 m2的大尺度上则可以体现高载畜率以及各载畜率水平之间的差异显著性。放牧会减少群落非优势物种的相对多度, 物种数和α多样性指数随着载畜率的升高而下降, 非优势物种能够反映α多样性。
| [1] | 刘起. 保护草地资源刻不容缓. 北方经济, 1999(3): 12–13. |
| [2] | 李延梅, 牛栋, 张志强. 国际生物多样性研究科学计划与热点述评. 生态学报, 2009, 29(4): 2115–2123. |
| [3] | Parton W J, Scurlock J M O, Ojima D S, Gilmanov T G, Scholes R J, Schimel D S, Kirchner T, Menaut J C, Seastedt T, Garcia Moya E, Kamnalrut A, Kinyamario I. Observations and modeling of biomass and soil organic matter dynamics for the grass-land biome worldwide. Global Biochemical Cycles, 1993, 7(4): 785–809. DOI:10.1029/93GB02042 |
| [4] | Smith M S. Global Change Impacts on Pastures and Rangelands (Implementation Plan). Canberra:GCTE Core Project Office, 1979. |
| [5] | 刘晓媛.放牧方式对草地植被多样性与稳定性关系的影响[D].长春:东北师范大学, 2010. |
| [6] | 李永宏. 内蒙古草原草场放牧退化模式研究及退化监测专家系统雏议. 植物生态学报, 1994, 18(1): 69–79. |
| [7] | McNaughton S J. Ecology of a grazing ecosystem:the Serengeti. Ecological Monographs, 1985, 55(3): 259–294. DOI:10.2307/1942578 |
| [8] | Milchunas D G, Lauenroth W K. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs, 1993, 63(4): 327–366. DOI:10.2307/2937150 |
| [9] | 王仁忠. 干扰对草地生态系统生物多样性的影响. 东北师大学报自然科学版, 1996(3): 112–116. |
| [10] | Olff H, Ritchie M E. Effects of herbivores on grassland plant diversity. Trends in Ecology & Evolution, 1998, 13(7): 261–265. |
| [11] | Hobbs R J, Huenneke L F. Disturbance, diversity, and invasion:Implications for conservation. Conservation Biology, 1992, 6(3): 324–337. DOI:10.1046/j.1523-1739.1992.06030324.x |
| [12] | 王明君, 韩国栋, 崔国文, 赵萌莉. 放牧强度对草甸草原生产力和多样性的影响. 生态学杂志, 2010, 29(5): 862–868. |
| [13] | 杨利民, 韩梅, 李建东. 中国东北样带草地群落放牧干扰植物多样性的变化. 植物生态学报, 2001, 25(1): 110–114. |
| [14] | Connell J H. Diversity in tropical rain forests and coral reefs. Science, 1978, 199(4335): 1302–1310. DOI:10.1126/science.199.4335.1302 |
| [15] | 王国杰, 汪诗平, 郝彦宾, 蔡学彩. 水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响. 生态学报, 2005, 25(7): 1649–1656. |
| [16] | 袁建立, 江小蕾, 黄文冰, 王刚. 放牧季节及放牧强度对高寒草地植物多样性的影响. 草业学报, 2004, 13(3): 16–21. |
| [17] | 杨婧, 褚鹏飞, 陈迪马, 王明玖, 白永飞. 放牧对内蒙古典型草原α、β和γ多样性的影响机制. 植物生态学报, 2014, 38(2): 188–200. |
| [18] | Arrhenius O. Species and area. Journal of Ecology, 1921, 19(1): 95–99. |
| [19] | Cain S A. The species-area curve. The American Midland Naturalist, 1938, 19(3): 573–581. DOI:10.2307/2420468 |
| [20] | Odum E P.生态学基础.孙儒泳等译.北京:人民教育出版社, 1981:144-149. |
| [21] | Magurran A E. Ecological Diversity and Its Measurement. Princeton:Princeton University press, 1988: 34–59. |
| [22] | 韩国栋, 焦树英, 毕力格图, 敖登高娃. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响. 生态学报, 2007, 27(1): 182–188. |
| [23] | 杨宝珍, 李博, 曾泗弟. 关于草原群落研究中样方面积大小的初步探讨. 植物生态与地植物学丛刊, 1964, 2(1): 111–117. |
| [24] | 杨利民, 韩梅, 李建东. 草地植物群落物种多样性取样强度的研究. 生物多样性, 1997, 5(3): 168–172. |
| [25] | 白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落α多样性研究. 生物多样性, 2000, 8(4): 353–360. |
| [26] | 张宇.不同载畜率下荒漠草原土壤水分空间异质性的研究[D].呼和浩特:内蒙古农业大学, 2014. |
| [27] | Reynolds H L, Hungate B A., Chapin Ⅲ F S, D'Antonio C M. Soil heterogeneity and plant competition in an annual grassland. Ecology, 1997, 78(7): 2076–2090. |
| [28] | 刘红梅.短花针茅草原群落特征与草地空间异质性对不同放牧制度的响应[D].呼和浩特:内蒙古农业大学, 2011. |
| [29] | Western A W, Blöschl G. On the spatial scaling of soil moisture. Journal of Hydrology, 1999, 217(3/4): 203–224. |
| [30] | 王忠武.载畜率对短花针茅荒漠草原生态系统稳定性的影响[D].呼和浩特:内蒙古农业大学, 2009. |