 2017, Vol. 37
2017, Vol. 37文章信息
- 韩广轩.
- HAN Guangxuan.
- 潮汐作用和干湿交替对盐沼湿地碳交换的影响机制研究进展
- Effect of tidal action and drying-wetting cycles on carbon exchange in a salt marsh: progress and prospects
- 生态学报. 2017, 37(24): 8170-8178
- Acta Ecologica Sinica. 2017, 37(24): 8170-8178
- http://dx.doi.org/10.5846/stxb201611182347
-
文章历史
- 收稿日期: 2016-11-18
- 网络出版日期: 2017-08-15

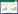




潮汐盐沼湿地处在陆海过渡带, 主要指海岸沿线受海洋潮汐周期性或间歇性影响的有盐生植物覆盖的咸水或淡咸水淤泥质滩涂, 是地球上高生产力植被类型之一[1-2]。潮汐盐沼湿地具有很高的固碳能力, 其碳的积累速度要远高于泥炭湿地[3]。研究表明, 全球潮汐盐沼的碳埋藏速率为(218±24) g m-2 a-1, 比陆地森林生态系统高40倍以上[4]。基于全球154处红树林和盐沼湿地的监测数据(除了中国和南美洲缺少监测数据外), 估算出全球红树林和盐沼湿地至少储藏44.6 Tg C a-1, 甚至更多[3]。同时, 周期性潮汐运动过程携带的大量的SO42-离子阻碍CH4的产生, 从而降低盐沼湿地CH4的排放量[5]。高的碳积累速率和低的CH4排放量, 使潮汐盐沼湿地的碳汇作用更加明显[6]。
近年来, 盐沼等滨海湿地的碳汇功能逐渐得到证实, 在减缓气候变化方面扮演着重要角色, 已得到国际组织和学术界的认可和重视。最新的统计报告称, 盐沼、海草和红树林等滨海湿地只占陆地生物量的0.05%, 但能从海洋及大气中储存和转移更多的碳(即“蓝碳”), 占全球生物吸收和固定碳总量的55%, 是地球上最密集的碳汇之一[7-8]。同时, 模型模拟表明, 气候变暖和海平面上升可能使得盐沼湿地更迅速的捕获和埋藏大气中的CO2, 因此盐沼湿地在减缓气候变化方面扮演着重要角色[8]。由联合国环境规划署(UNEP)、联合国粮食及农业组织(FAO)、联合国教科文组织海洋学委员会(IOC/UNESCO)联合发布的《蓝碳:健康海洋的固碳作用》(Blue Carbon-The Role of Healthy Oceans in Binding Carbon)报告指出, 潮汐盐沼湿地中的“蓝碳”在应对气候变化方面起着重要作用, 呼吁世界各国立即采取行动, 维持和恢复“蓝碳”, 保护海岸带生态系统的碳汇功能。
潮汐盐沼湿地与其他湿地类型最大的区别和最显著的特征是在周期性潮汐作用下出现淹没和暴露, 同时伴随盐分表聚与淋洗的干湿交替, 导致盐沼湿地系统的生物地化循环过程具有其特异性, 可能是控制盐沼湿地碳交换过程和碳收支平衡的关键因素。潮汐盐沼湿地大部分面积除在大潮期间, 平时基本上不能被海水浸没, 明显地出现以半月为周期的淹没和暴露的干湿交替, 并且在暴露期间出现土壤龟裂和盐碱化现象。但是, 周期性潮汐作用下滨海湿地碳循环的关键机理尚不清楚, 其固碳潜力、碳汇通量和封存量的数据还很少, 目前尚无一个全球公认的机制来正确认知蓝色碳汇的重要性[9]。因此, 阐明潮汐盐沼湿地碳交换过程的影响机制, 揭示盐沼湿地蓝色碳汇的形成过程与机制, 对于评估滨海湿地“蓝碳”在区域及全球碳收支的贡献具有十分重要的作用和意义。
1 盐沼湿地碳交换过程与观测方法盐沼湿地的碳交换包括垂直方向上的CO2和CH4交换和横向方向上的可溶性有机碳(DOC)、可溶性无机碳(DIC)、颗粒有机碳(POC)交换(图 1)。盐沼湿地中, 大气中的CO2通过光合作用被植物吸收并合成有机物, 这部分固定的碳称为总初级生产力(GPP), 同时植物自身消耗一部分碳用于维持生命并释放CO2(自养呼吸);植物凋落物和土壤有机碳在好氧环境下经微生物矿化分解释放CO2(异养呼吸), 自养呼吸和异养呼吸之和为生态系统呼吸(Reco)。盐沼湿地净生态系统CO2交换(NEE)是植被光合固定碳(GPP)与生态系统呼吸释放碳(Reco)之间相互平衡的结果[10]。另外, 湿地又是向大气排放CH4的碳源, 淹水时植物残体和土壤有机碳在厌氧环境下产生CH4, 通过扩散、气泡传输和植物传输释放到大气中。但在植被根际的微好氧环境以及表层以下浅层土壤氧化区域, 部分CH4被甲烷氧化菌所氧化, CH4排放量降低[11]。盐沼湿地的碳源汇功能取决于生态系统与大气间CO2和CH4交换的净收支。潮汐运动通过潮汐淹水和干湿交替直接作用于植被生产力[12]以及土壤有机碳的形成和分解[5-6], 还可以通过沉积物的供给直接影响潮间带湿地碳封存能力[3-4], 或通过影响营养物质可用性、土壤温度、盐度和氧化还原电位等环境因素间接作用于这些碳过程[13], 从而影响盐沼湿地的碳汇形成机制。另外, 作为陆地和海洋生态系统之间过渡生态系统类型, 潮汐盐沼湿地土壤有机碳在海洋潮汐和地表径流的作用下能够以DOC、DIC、POC的形式进入邻近水体。水平方向上碳迁移和输出是盐沼湿地通过水文过程实现土壤碳输出的一个主要途径[14], 在盐沼湿地碳循环中发挥着重要作用[15-16]。因此, 研究潮汐盐沼湿地的碳收支时, 应该同时考虑植被-大气界面CO2和CH4交换和土壤-水体界面的碳迁移过程, 才能准确地评估盐沼湿地的碳源汇功能。

|
| 图 1 潮汐盐沼湿地碳交换关键过程示意图 Fig. 1 Conceptual diagram for key process of carbon exchange in a tidal salt marsh GPP:总初级生产力, Gross Primary Production;Reco:生态系统呼吸, Ecosystem Respiration;NEE:净生态系统CO2交换, Net Ecosystem CO2 Exchange;DOC:溶解性有机碳, Dissolved Organic Carbon;DIC:可溶性无机碳, Dissolved Inorganic Carbon;POC:颗粒有机碳, Particulate Organic Carbon |
目前, 国内外学者主要采用涡度相关法(Eddy Covariance)和箱式法测定盐沼湿地CO2和CH4交换, 采用野外采样和室内分析测定土壤和水流中DOC、DIC、POC的浓度和迁移通量。涡度相关观测技术和理论的不断发展为探讨生态系统尺度的CO2和CH4交换的时空变化提供了新途径, 成为长期测算生态系统碳通量最可靠和切实可行的方法[17], 被认为是现今能直接测定陆地生态系统与大气间物质与能量交换通量的标准方法。目前, 国内外学者在美国德克萨斯州墨西哥湾沿岸[18]、弗吉尼亚州东海岸[19]以及Everglades[20]、法国西南部潮间带泥滩[21]等以及我国的长江口[22]、黄河口[10, 23]、闽江口[24-25]、辽河口[26]和福建漳江口开展了盐沼湿地生态系统CO2交换研究。但是当前对盐沼湿地CH4排放的观测主要基于箱式法[24-25, 27]。传统的箱法通量测定需要人工操作, 难以长期连续自动观测;同时受到空间不均一性的影响, 进行大尺度扩展有一定的困难[28]。目前箱式法经过改良后可以进行自动观测, 但是其时间分辨率较低, 难以捕捉完整潮汐运动过程对CO2和CH4交换的影响。与箱式法相比, 涡度相关技术提供了一个更大空间、更长时间和更高时间分辨率的通量观测数据[17], 有利于在生态系统尺度上量化潮汐运动和干湿交替对CO2和CH4交换的影响, 进而准确评估盐沼湿地的碳汇功能。
2 潮汐作用和干湿交替对盐沼湿地-大气间碳交换的影响潮汐盐沼湿地CO2和CH4交换受到的影响与其他湿地类型最大的区别在于, 潮汐的存在不仅影响水位, 还存在特殊的涨落潮水周期特征[29]。一方面, 潮汐过程(涨落潮和淹水)主要通过影响氧的可利用率、气体扩散率及微生物活性直接或间接影响植被的光合作用和呼吸作用[20, 30], 进而影响湿地-大气间CO2交换过程[22, 29]。同时, 潮汐淹水会使光合作用的有效叶面积减少[17], 导致光合作用强度降低, 因此淹水减少了盐沼湿地白天植被光合作用的最大速率[10];另外, 潮汐淹水使得盐沼湿地排向大气的CO2通量显著减少[23], 与涨潮前和落潮后相比淹水期湿地生态系统CO2排放量最低[24], 潮汐高度与CO2释放量呈负相关[29];NEE的减少与淹水的深度和持续时间成正比[30]。因此, 潮间带湿地淹水程度伴随潮汐而变化, 使湿地CO2交换产生与潮汐运动协同或略滞后的周期性, 持续短暂但却作用强烈[22, 30]。
另一方面, 涨落潮过程潮汐带来的富含硫酸盐等电子受体的海水入侵, 能增加电子受体与CH4产生对电子的竞争, 使盐沼湿地产CH4过程向硫酸盐还原作用过程转变, 由此产生CH4的减排效应[25, 31];在涨潮过程中, 土壤还不足以形成完全的厌氧环境, 加之涨潮引起的土壤表面水位波动使得空气中的氧气溶解, 减少CH4生产的同时, 增加了CH4的氧化[31-32]。潮汐淹水提供了CH4产生的厌氧环境, 但是水位的升高不一定造成CH4通量的增加, 还需综合考虑滨海湿地CH4的生产潜力、基底物质以及由水位改变造成的光合作用的改变。例如, 过多的淹水可能会造成CH4释放过程中的阻碍, 减少植物对于CH4的传输并延迟CH4的气泡扩散, 因此盐沼湿地涨落潮过程中CH4通量均小于涨潮前和落潮后CH4通量[24];平潮期由于CH4的部分溶解, CH4释放低于涨潮期和落潮期[32]。另外, 周期性的潮汐运动足以扰乱湿地的盐沼环境(如盐度、氧化还原电位、土壤温度、饱和度等), 从而影响植被生长与分布[12]。植物可以固定碳和通过根系分泌物和凋落物为产甲烷菌提供底物, 也会通过传输作用影响CH4的释放[25, 32]。
潮汐作用引起的干湿交替是盐沼湿地所经历的最普通和频繁的自然过程[33-34], 使土壤经历一个干旱-再湿润-淹水-湿润的物理、化学和生物变化过程。周期性干湿交替对盐沼湿地CO2和CH4的生产、吸收和传输等各个过程均可能产生深刻的影响[31], 主要表现在以下几个方面: (1)干湿交替会引起土壤收缩和膨胀, 从而破坏土壤的物理聚合[35], 影响土壤结构(团聚体结构、孔隙和通气状况), 从而影响土-气界面之间厌氧和好氧过程的发生。例如, 大潮期较高的土壤含水量, 抑制空气中O2扩散进入土壤, 而土壤原有O2很快被消耗完, 从而导致了土壤氧化还原电位(Eh)迅速降低[36]。(2)潮汐驱动下干湿交替伴随着土壤中水溶性盐分的表聚与淋洗, 导致土壤盐度周期波动, 成为盐沼湿地最显著的环境特征之一[33]。在暴露失水过程中, 孔隙水在毛细作用下向土壤表层迁移, 引起带水溶性盐分在土壤表层富集, 出现龟裂和盐碱化现象;在潮汐淹水过程中, 水分向土壤深层下渗, 根层土壤淋洗脱盐[33]。湿地土壤盐度增加可能会通过渗透压胁迫抑制土壤中微生物细胞的活性从而降低其对土壤有机质的分解速率[37], 因此土壤CO2和CH4的释放与盐度均呈负相关[38-39]。同时, 过高的盐度会通过渗透压胁迫使植物失水[18], 降低植物的光合作用和初级生产力(CO2吸收), 因而与植物活性紧密耦合的生态系统呼吸作用(CO2释放)也随盐度增加而降低[40]。(3)干湿交替对土壤微生物活性和群落结构产生重要影响。土壤干燥过程中, 土壤水势降低造成渗透休克, 从而导致细胞破裂死亡[41], 此时土壤中微生物以好氧型群落为主;干燥土壤湿润过程可能会导致土壤微生物细胞的渗透压发生变化从而使得微生物细胞溶解, 或使胞内溶质渗漏[42], 微生物群落转变为以厌氧型为主。研究表明, 干湿交替能够极大地激发微生物活性, 导致土壤有机质矿化速率的急剧增加, 从而引起短期内快速释放大量的CO2[43-45], 这种现象称为“Birch效应”。这种由干湿交替引起的土壤CO2短暂脉冲式释放很大程度上决定着长时间尺度温室气体释放的总量, 是土壤含碳温室气体释放的关键过程[45]。(4)干湿交替通过影响土壤水分和土壤盐度等环境因子进而控制植物体内的主要生理过程, 如光合作用、呼吸作用、脂质代谢、蛋白质合成等[12]。同时, 干湿交替中土壤长期暴露所带来的的高盐度是潮间带湿地的独特环境因素, 过高的盐度会通过渗透压胁迫使植物失水, 只有适应高盐度的植物才能存活, 并形成潮间带湿地多样的植物分布格局和生产力状况[18]。全球气候变化背景下, 局部地区可能经历频繁剧烈的干湿交替, 按照成因可以分为两大类:一类是降雨引起的, 发生在干旱、半干旱和地中海气候区[42, 44-45];第二类是潮汐作用引起的, 发生在滨海湿地。前人对于降雨引起的干湿交替对干旱半干旱地区碳过程的影响已开展大量工作并取得系列进展[44-46], 但是潮汐引起的干湿交替对滨海湿地碳交换过程的控制机理尚不明确, 有待深入。
3 潮汐作用和干湿交替对盐沼湿地-水体间碳交换的影响作为陆地和海洋生态系统之间过渡生态系统类型, 盐沼湿地土壤-水体界面DOC、DIC、POC迁移转化及其输出是其碳收支估算中不容忽视的组成部分[47-48]。湿地DOC主要自于湿地植物、碎屑物中碳的淋溶以及沉积物中碳的释放, 而POC主要来自于湿地中生长的植物[49-50]。例如, 加拿大Ontario泥炭沼泽通过降水截获的DOC约为(1.5±0.7) gC m-2 a-1, 但每年的DOC输出量达(8.3±3.7) gC m-2 a-1[51]。美国滨海湿地输出到邻近河口的DOC通量为(180±12.6) g m-2 a-1[47], 比从淡水湿地输出的碳量大了一个数量级。土壤微生物在分解利用DOC的同时, 可以将其转变为DIC, 并通过径流与相邻水域进行交换[52], 这部分碳通量可高达土壤溶液DOC总量的40%[53]。另外, POC是生物摄食和代谢产物的主要形式, 无论在土壤孔隙还是在径流中, 其通量都仅次于DOC[54], 并在一定程度上控制着DOC的分布。通过潮汐的作用, 九龙江口湿地具有向毗邻水域输出有机碳的作用, DOC和POC的输出量分别为0.07 kg hm-2 d-1和0.01 kg hm-2 d-1, 是近海水域有机碳的源[55]。可以看出, 盐沼湿地由于高的生产力以及和潮汐的频繁物质交换, 对于临近水体而言是重要的碳源, 因此潮汐盐沼湿地的碳交换闭合研究必须包括垂直方向上的碳交换和横向方向上的碳交换。
毫无疑问, 水文条件的改变对盐沼湿地DOC、DIC和POC的产生和释放都有显著影响。首先, 周期性潮汐运动和降雨引起的地表径流控制着水位和水流速度, 直接影响盐沼湿地的氧化还原状态[15, 56]和碳的迁移速率[57]。其次, 在潮汐作用下盐沼湿地水体和土壤盐度从海到陆呈现梯度变化, 进而影响有机碳的运移和沉积[58-59]。再次, 由于潮汐和降雨导致湿地土壤表层不断经历着干湿交替过程, 干湿交替能够改变土壤结构和通气条件, 控制着好氧和厌氧呼吸作用之间的平衡, 并影响土壤有机碳含量的稳定性和土壤微生物活动, 从而改变土壤碳动态及其趋势[15, 60]。目前一些研究已经利用涡动通量塔确定了盐沼湿地和大气之间垂直方向上的碳交换[18-19, 21-22], 但是并没有把水平方向上碳交换的测量耦合在一起[16]。
4 潮汐作用和干湿交替对盐沼湿地碳汇形成机制的影响盐沼湿地不仅是CO2的“汇”与“源”, 也是CH4的重要来源, 同时DOC、DIC和POC交换及其输出也是其碳收支估算中不容忽视的组成部分, 因此碳吸收与碳排放之间的动态过程决定了盐沼湿地的碳汇功能。潮汐作用及其干湿交替影响盐沼湿地的土壤物理化学性质[2, 13, 33]、水文状况[13, 42, 61]、氧化还原状态[15, 56]以及植被生长和分布[12, 18]等诸多环境条件, 从而影响湿地碳交换通量的大小与方向[2, 31, 57], 最终影响盐沼湿地碳汇与碳源功能的相互转化。例如, 潮间带湿地在盐度较低、土壤水分较高的情况下固定CO2, 而在盐度较高、土壤水分较低的情况下则排放CO2[18]。在单独增加盐度时, 潮汐淡水湿地净生态系统生产力降低了55%, 但当盐度和水文因子被同时调控时, 净生态系统生产力却没有发生显著改变[40]。干湿交替通过影响土壤水分条件决定湿地的氧气环境, 由此使湿地在产CH4环境和氧化CH4环境两种状态间互相转化[34, 45], 因此干湿交替有调节湿地CO2吸收和CH4排放平衡的作用。另外, 湿地水位与CH4产生量呈正相关, 但却与CO2产生量有一定的负相关关系[62]。潮汐水位对盐沼湿地CO2:CH4的变化具有一定的调节作用[25], 涨潮前、涨落潮过程和落潮后3个过程盐沼湿地CO2:CH4对潮汐的响应并不一致。另外, DOC迁移和输出是盐沼湿地土壤碳输出的一个主要途径, 而潮汐作用在盐沼湿地DOC输出中发挥着关键作用。同时, 滨海湿地水平碳交换与CO2和CH4的排放密切相关。例如, 高潮时滨海淡水沼泽的土壤CO2通量与DOC浓度呈正相关[15]。湿地CH4排放的季节变化受土壤DOC的调节, 土壤DOC与CH4排放量呈显著正相关[63-64]。因此, 潮汐作用及其由此引起的干湿交替可能控制着盐沼湿地碳交换的通量大小、方向以及之间的转化, 最终决定盐沼湿地的碳汇形成机制和碳汇功能。
5 问题与展望综上所述, 潮汐盐沼湿地与其他湿地类型最大的区别和最显著的特征是在周期性潮汐作用下出现淹没和暴露, 同时伴随盐分表聚与淋洗的干湿交替, 导致盐沼湿地系统的生物地化循环过程具有其特异性[29, 33], 可能是控制盐沼湿地碳交换过程和碳收支平衡的关键因素。据预测, 2100年海平面也会比20世纪末升高0.26—0.55 m[65], 海平面上升会改变潮汐水位、淹水频率和干湿交替周期的改变, 对盐沼湿地的碳汇形成机制产生潜在影响[61]。国内外学者对盐沼湿地CO2、CH4等气体通量和水平方向上碳输出进行了较为深入的研究, 但是潮汐水动力过程及其周期性干湿交替对盐沼湿地碳交换关键过程和碳汇形成机制的影响尚不十分清楚。另外, 以往相关研究通常孤立地考虑垂直方向上CO2或CH4交换或横向方向上的DOC、DIC和POC交换对潮汐盐沼湿地的碳平衡进行评估, 显然不够准确, 这些将在很大程度上制约着对滨海盐沼湿地碳循环过程的整体理解以及盐沼湿地“蓝碳”吸存能力的评估和预测。
今后滨海盐沼湿地碳循环在以下方面的研究仍需加强:
(1) 系统研究潮汐不同阶段对盐沼湿地碳交换过程的影响。周期性潮汐作用使土壤经历一个干旱-再湿润-淹水-湿润的交替变化, 分析盐沼湿地碳交换过程对潮汐不同阶段(长期暴露后的干旱阶段-涨潮时再湿润阶段-潮汐淹水阶段-落潮后湿润阶段)的响应机制(图 2)。

|
| 图 2 依据潮汐过程, 盐沼湿地土壤水分状况划分为干旱阶段、再湿润阶段、潮汐淹水阶段和落潮后湿润阶段 Fig. 2 According to tidal process, the situation of soil moisture in a tidal salt marsh can be divided into drought stage, rewetting stage, tidal flooding and wet stage after tides |
(2) 深入分析潮汐作用下盐沼湿地碳交换的微生物机制。潮汐作用下周期性干湿交替直接影响营养物质可用性[13]、土壤温度和盐度、土壤氧气环境[34, 45]、氧化还原状态[15, 56]等环境因素, 进而改变土壤微生物群系和活性以及土壤微生物群落储存、利用碳的方式[66], 最终影响湿地碳的吸收与排放。
(3) 关注潮汐水动力作用对盐沼湿地DOC、DIC和POC产生、释放以及向邻近水体输出的影响。潮汐作用下盐沼湿地碳与近海水体之间的横向碳交换通量对于滨海湿地碳库动态变化有着重要贡献, 但是关于潮汐对滨海湿地土壤碳库及碳输出的影响研究较为薄弱。
(4) 阐明潮汐作用对盐沼湿地碳汇形成机制的影响。采用数据统计与波谱解析方法, 计算潮汐运动与碳交换过程的波动规律和局部特征, 量化潮汐运动与盐沼湿地CO2和CH4交换以及水平碳交换之间的关系, 整体上评估潮汐作用对盐沼湿地碳汇功能的影响。
(5) 建立潮汐盐沼湿地碳循环模型。现有滨海湿地碳循环模型通常将整个生态系统看做一个均质的系统, 忽略了潮汐运动对盐沼湿地碳过程的影响, 因此应在模型中纳入潮汐水动力过程作为变量, 以提高模型模拟的正确性和准确性。
| [1] | Boorman L. Saltmarsh Review:an overview of coastal saltmarshes, their dynamic and sensitivity characteristics for conservation and management. JNCC Report No. 334. Peterborough, UK:JNCC, 2003. http://jncc.defra.gov.uk/page-2347-theme=print |
| [2] | 仲启铖, 王开运, 周凯, 来琦芳. 潮间带湿地碳循环及其环境控制机制研究进展. 生态环境学报, 2015, 24(1): 174–182. |
| [3] | Chmura B G, Anisfeld S C, Cahoon D R, Lynch J C. Global carbon sequestration in tidal, saline wetland soils. Global Biogeochemical Cycles, 2003, 17(4). DOI:10.1029/2002GB001917 |
| [4] | Mcleod E, Chmura G L, Bouillon S, Salm R, Björk M, Duarte C M, Lovelock C E, Schlesinger W H, Silliman B R. A blueprint for blue carbon:toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Frontiers in Ecology and the Environment, 2011, 9(10): 552–560. DOI:10.1890/110004 |
| [5] | Choi Y, Wang Y. Dynamics of carbon sequestration in a coastal wetland using radiocarbon measurements. Global Biogeochemical Cycles, 2004, 18(4). DOI:10.1029/2004GB002261 |
| [6] | Drake K, Halifax H, Adamowicz S C, Craft C. Carbon sequestration in tidal salt marshes of the Northeast United States. Environmental Management, 2015, 56(4): 998–1008. DOI:10.1007/s00267-015-0568-z |
| [7] | Nellemann C, Corcoran E, Duarte C, Valdés L, Young C D, Fonseca L, Grimsditch G. Blue carbon:the role of healthy oceans in binding carbon:a rapid response assessment. Arendal, Norway:GRID-Arendal, 2009:589-598. |
| [8] | Kirwan M L, Mudd S M. Response of salt-marsh carbon accumulation to climate change. Nature, 2012, 489(7417): 550–553. DOI:10.1038/nature11440 |
| [9] | 白雪瑞, 熊国祥. 香山科学会议第396-400次学术讨论会简述. 中国基础科学, 2011, 13(5): 25–31. |
| [10] | Han G X, Chu X J, Xing Q H, Li D J, Yu J B, Luo Y Q, Wang G M, Mao P L, Rafique R. Effects of episodic flooding on the net ecosystem CO2 exchange of a supratidal wetland in the Yellow River Delta. Journal of Geophysical Research, 2015, 120(8): 1506–1520. |
| [11] | Vann C D, Megonigal J P. Elevated CO2 and water depth regulation of methane emissions:comparison of woody and non-woody wetland plant species. Biogeochemistry, 2003, 63(2): 117–134. DOI:10.1023/A:1023397032331 |
| [12] | Parida A K, Das A B. Salt tolerance and salinity effects on plants:a review. Ecotoxicology and Environmental Safety, 2005, 60(3): 324–349. DOI:10.1016/j.ecoenv.2004.06.010 |
| [13] | Mitchell A, Baldwin D S. Effects of desiccation/oxidation on the potential for bacterially mediated P release from sediments. Limnology and Oceanography, 1998, 43(3): 481–487. DOI:10.4319/lo.1998.43.3.0481 |
| [14] | 曹磊. 山东半岛北部典型滨海湿地碳的沉积与埋藏[D]. 青岛: 中国科学院大学(海洋研究所), 2014. http://cdmd.cnki.com.cn/Article/CDMD-80068-1014308963.htm |
| [15] | Chambers L G, Osborne T Z, Reddy K R. Effect of salinity-altering pulsing events on soil organic carbon loss along an intertidal wetland gradient:a laboratory experiment. Biogeochemistry, 2013, 115(1/3): 363–383. |
| [16] | Fagherazzi S, Wiberg P L, Temmerman S, Struyf E, Zhao Y, Raymond P A. Fluxes of water, sediments, and biogeochemical compounds in salt marshes. Ecological Processes, 2013, 2: 3. DOI:10.1186/2192-1709-2-3 |
| [17] | Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems:past, present and future. Global Change Biology, 2003, 9(4): 479–492. DOI:10.1046/j.1365-2486.2003.00629.x |
| [18] | Heinsch F A, Heilman J L, Mcinnes K J, Cobos D R, Zuberer D A, Roelke D L. Carbon dioxide exchange in a high marsh on the Texas Gulf Coast:effects of freshwater availability. Agricultural and Forest Meteorology, 2004, 125(1/2): 159–172. |
| [19] | Kathilankal J C, Mozdzer T J, Fuentes J D, D'Odorico P, Mcglathery K J, Zieman J C. Tidal influences on carbon assimilation by a salt marsh. Environmental Research Letters, 2008, 3(4): 044010. DOI:10.1088/1748-9326/3/4/044010 |
| [20] | Jimenez K L, Starr G, Staudhammer C L, Schedlbauer J L, Loescher H W, Malone S L, Oberbauer S F. Carbon dioxide exchange rates from short-and long-hydroperiod Everglades freshwater marsh. Journal of Geophysical Research, 2012, 117(G4). DOI:10.1029/2012JG002117 |
| [21] | Polsenaere P, Lamaud E, Lafon V, Bonnefond J M, Bretel P, Delille B, Deborde J, Loustau D, Abril G. Spatial and temporal CO2 exchanges measured by Eddy Covariance over a temperate intertidal flat and their relationships to net ecosystem production. Biogeosciences, 2012, 9(1): 249–268. DOI:10.5194/bg-9-249-2012 |
| [22] | Guo H Q, Noormets A, Zhao B, Chen J Q, Sun G, Gu Y J, Li B, Chen J K. Tidal effects on net ecosystem exchange of carbon in an estuarine wetland. Agricultural and Forest Meteorology, 2009, 149(11): 1820–1828. DOI:10.1016/j.agrformet.2009.06.010 |
| [23] | 邢庆会, 韩广轩, 于君宝, 吴立新, 杨利琼, 毛培利, 王光美, 谢宝华. 黄河口潮间盐沼湿地生长季净生态系统CO2交换特征及其影响因素. 生态学报, 2014, 34(17): 4966–4979. |
| [24] | 仝川, 鄂焱, 廖稷, 姚顺, 王维奇, 黄佳芳, 张林海, 杨红玉, 曾从盛. 闽江河口潮汐沼泽湿地CO2排放通量特征. 环境科学学报, 2011, 31(12): 2830–2840. |
| [25] | 王维奇, 曾从盛, 仝川, 王纯. 闽江河口潮汐湿地二氧化碳和甲烷排放化学计量比. 生态学报, 2012, 32(14): 4396–4402. |
| [26] | Zhou L, Zhou G S, Jia Q Y. Annual cycle of CO2 exchange over a reed (Phragmites australis) wetland in Northeast China. Aquatic Botany, 2009, 91(2): 91–98. DOI:10.1016/j.aquabot.2009.03.002 |
| [27] | 于贵瑞, 王秋凤, 朱先进. 区域尺度陆地生态系统碳收支评估方法及其不确定性. 地理科学进展, 2011, 30(1): 103–113. DOI:10.11820/dlkxjz.2011.01.013 |
| [28] | Hommeltenberg J, Mauder M, Drösler M, Heidbach K, Werle P, Schmid H P. Ecosystem scale methane fluxes in a natural temperate bog-pine forest in southern Germany. Agricultural and Forest Meteorology, 2014, 198-199: 273–284. DOI:10.1016/j.agrformet.2014.08.017 |
| [29] | 马安娜, 陆健健. 长江口崇西湿地生态系统的二氧化碳交换及潮汐影响. 环境科学研究, 2011, 24(7): 716–721. |
| [30] | Moffett K B, Wolf A, Berry J A, Gorelick S M. Salt marsh-atmosphere exchange of energy, water vapor, and carbon dioxide:effects of tidal flooding and biophysical controls. Water Resources Research, 2010, 46(10): W10525. |
| [31] | 王进欣, 王今殊, 钦佩, 张维康. 生源气体排放的潮周期动态研究:关键科学问题与不确定性. 海洋湖沼通报, 2011(4): 134–143. |
| [32] | Tong C, Wang W Q, Zeng C S, Marrs R. Methane (CH4) emission from a tidal marsh in the Min River estuary, southeast China. Journal of Environmental Science and Health, Part A:Toxic/hazardous Substances and Environmental Engineering, 2010, 45(4): 506–516. DOI:10.1080/10934520903542261 |
| [33] | 侯立军. 长江口滨岸潮滩营养盐环境地球化学过程及生态效应[D]. 上海: 华东师范大学, 2004. http://cdmd.cnki.com.cn/article/cdmd-10269-2004087322.htm |
| [34] | 王健波, 张燕卿, 严昌荣, 刘恩科. 干湿交替条件下土壤有机碳转化及影响机制研究进展. 土壤通报, 2013, 44(4): 998–1004. |
| [35] | Inglima I, Alberti G, Bertolini T, Vaccari F P, Gioli B, Miglietta F, Cotrufo M F, Peressotti A. Precipitation pulses enhance respiration of Mediterranean ecosystems:the balance between organic and inorganic components of increased soil CO2 efflux. Global Change Biology, 2009, 15(5): 1289–1301. DOI:10.1111/gcb.2009.15.issue-5 |
| [36] | Seybold C A, Mersie W, Huang J, McNamee C. Soil redox, pH, temperature, and water-table patterns of a freshwater tidal wetland. Wetlands, 2002, 22(1): 149–158. DOI:10.1672/0277-5212(2002)022[0149:SRPTAW]2.0.CO;2 |
| [37] | Setia R, Marschner P, Baldock J, Chittleborough D. Is CO2 evolution in saline soils affected by an osmotic effect and calcium carbonate. Biology and Fertility of Soils, 2010, 46(8): 781–792. DOI:10.1007/s00374-010-0479-3 |
| [38] | Weston N B, Vile M A, Neubauer S C, Velinsky D J. Accelerated microbial organic matter mineralization following salt-water intrusion into tidal freshwater marsh soils. Biogeochemistry. Biogeochemistry. Biogeochemistry, 2011, 102(1/3): 135–151. |
| [39] | Poffenbarger H J, Needelman B A, Megonigal J P. Salinity influence on methane emissions from tidal marshes. Wetlands, 2011, 31(5): 831–842. DOI:10.1007/s13157-011-0197-0 |
| [40] | Neubauer S C. Ecosystem responses of a tidal freshwater marsh experiencing saltwater intrusion and altered hydrology. Estuaries and Coasts, 2013, 36(3): 491–507. DOI:10.1007/s12237-011-9455-x |
| [41] | Magid J, Kjærgaard C, Gorissen A, Kuikman P J. Drying and rewetting of a loamy sand soil did not increase the turnover of native organic matter, but retarded the decomposition of added 14C-labelled plant material. Soil Biology and Biochemistry, 1999, 31(4): 595–602. DOI:10.1016/S0038-0717(98)00164-3 |
| [42] | Fierer N, Schimel J P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biology and Biochemistry, 2002, 34(6): 777–787. DOI:10.1016/S0038-0717(02)00007-X |
| [43] | Birch H F. The effect of soil drying on humus decomposition and nitrogen availability. Plant and Soil, 1958, 10(1): 9–31. DOI:10.1007/BF01343734 |
| [44] | Borken W, Matzner E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Global Change Biology, 2009, 15(4): 808–824. DOI:10.1111/gcb.2009.15.issue-4 |
| [45] | Schimel J P, Wetterstedt J Å M, Holden P A, Trumbore S E. Drying/rewetting cycles mobilize old C from deep soils from a California annual grassland. Soil Biology and Biochemistry, 2011, 43(5): 1101–1103. DOI:10.1016/j.soilbio.2011.01.008 |
| [46] | 欧阳扬, 李叙勇. 干湿交替频率对不同土壤CO2和N2O释放的影响. 生态学报, 2013, 33(4): 1251–1259. |
| [47] | Bergamaschi B A, Krabbenhoft D P, Aiken G R, Patino E, Rumbold D G, Orem W H. Tidally driven export of dissolved organic carbon, total mercury, and methylmercury from a mangrove-dominated estuary. Environmental Science & Technology, 2012, 46(3): 1371–1378. |
| [48] | Dinsmore K J, Billett M F, Dyson K E. Temperature and precipitation drive temporal variability in aquatic carbon and GHG concentrations and fluxes in a peatland catchment. Global Change Biology, 2013, 19(7): 2133–2148. DOI:10.1111/gcb.12209 |
| [49] | Thacker S A, Tipping E, Baker A, Gondar D. Development and application of functional assays for freshwater dissolved organic matter. Water Research, 2005, 39(18): 4559–4573. DOI:10.1016/j.watres.2005.08.020 |
| [50] | 李玲, 仇少君, 刘京涛, 刘庆, 陆兆华. 土壤溶解性有机碳在陆地生态系统碳循环中的作用. 应用生态学报, 2012, 23(5): 1407–1414. |
| [51] | Fraser C J D, Roulet N T, Moore T R. Hydrology and dissolved organic carbon biogeochemistry in an ombrotrophic bog. Hydrological Processes, 2001, 15(16): 3151–3166. DOI:10.1002/(ISSN)1099-1085 |
| [52] | Bouillon S, Borges A V, Castañeda-Moya E, Diele K, Dittmar T, Duke N C, Kristensen E, Lee S Y, Marchand C, Middelburg J J, Rivera-Monroy V H, Smith T J Ⅲ, Twilley R R. Mangrove production and carbon sinks:a revision of global budget estimates. Global Biogeochemical Cycles, 2008, 22(2): GB2013. |
| [53] | Shibata H, Mitsuhashi H, Miyake Y, Nakano S. Dissolved and particulate carbon dynamics in a cool-temperate forested basin in northern Japan. Hydrological Processes, 2001, 15(10): 1817–1828. DOI:10.1002/(ISSN)1099-1085 |
| [54] | Fiedler S, Höll B S, Freibauer A, Stahr K, Drösler M, Schloter M, Jungkunst H F. Particulate organic carbon (POC) in relation to other pore water carbon fractions in drained and rewetted fens in Southern Germany. Biogeosciences, 2008, 5(6): 1615–1623. DOI:10.5194/bg-5-1615-2008 |
| [55] | 邱悦, 叶勇. 九龙江口红树林与毗邻水域营养盐和有机碳的潮水交换. 厦门大学学报:自然科学版, 2013, 52(5): 718–721. |
| [56] | Wilson L, Wilson J, Holden J, Johnstone I, Armstrong A, Morris M. Ditch blocking, water chemistry and organic carbon flux:evidence that blanket bog restoration reduces erosion and fluvial carbon loss. Science of the Total Environment, 2011, 409(11): 2010–2018. DOI:10.1016/j.scitotenv.2011.02.036 |
| [57] | van den Berg L J L, Shotbolt L, Ashmore M R. Dissolved organic carbon (DOC) concentrations in UK soils and the influence of soil, vegetation type and seasonality. Science of the Total Environment, 2012, 427-428: 269–276. DOI:10.1016/j.scitotenv.2012.03.069 |
| [58] | Mayer L M. Surface area control of organic carbon accumulation in the continental shelf sediments. Geochemica et Cosmochimica Acta, 1994, 58(4): 1271–1284. DOI:10.1016/0016-7037(94)90381-6 |
| [59] | Zhang J, Wu Y, Jennerjahn T C, Ittekkot V, He Q. Distribution of organic matter in the Changjiang (Yangtze River) Estuary and their stable carbon and nitrogen isotopic ratios:Implications for source discrimination and sedimentary dynamics. Marine Chemistry, 2007, 106(1/2): 111–126. |
| [60] | 张雪雯, 莫熠, 张博雅, 高居娟, 高俊琴. 干湿交替及凋落物对若尔盖泥炭土可溶性有机碳的影响. 湿地科学, 2014, 12(2): 134–140. |
| [61] | IPCC. Climate Change 2013:The Physical Science Basis.Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom, New York, NY, USA:Cambridge University Press, 2013. |
| [62] | 孟伟庆, 吴绽蕾, 王中良. 湿地生态系统碳汇与碳源过程的控制因子和临界条件. 生态环境学报, 2011, 20(8/9): 1359–1366. |
| [63] | 杨文燕, 宋长春, 张金波. 沼泽湿地孔隙水中溶解有机碳、氮浓度季节动态及与甲烷排放的关系. 环境科学学报, 2006, 26(10): 1745–1750. DOI:10.3321/j.issn:0253-2468.2006.10.026 |
| [64] | Zhan M, Cao C G, Wang J P, Yang J, Cai M L, Yue L X, Shahrear A. Dynamics of methane emission, active soil organic carbon and their relationships in wetland integrated rice-duck systems in Southern China. Nutrient Cycling in Agroecosystems, 2011, 89(1): 1–13. DOI:10.1007/s10705-010-9371-7 |
| [65] | DeLaune R D, White J R. Will coastal wetlands continue to sequester carbon in response to an increase in global sea level?:a case study of the rapidly subsiding Mississippi river deltaic plain. Climatic Change, 2012, 110(1/2): 297–314. |
| [66] | Olsen Y S, Dausse A, Garbutt A, Ford H, Thomas D N, Jones D L. Cattle grazing drives nitrogen and carbon cycling in a temperate salt marsh. Soil Biology and Biochemistry, 2011, 43(3): 531–541. DOI:10.1016/j.soilbio.2010.11.018 |