 2017, Vol. 37
2017, Vol. 37文章信息
- 徐玲玲, 李昊, 王建林, 王志伟.
- XU Lingling, LI Hao, WANG Jianlin, WANG Zhiwei.
- 河北固城夏玉米比叶面积对水分梯度的响应
- Response of specific leaf area of summer maize to water gradient in Hebei Gucheng
- 生态学报. 2017, 37(23): 8101-8106
- Acta Ecologica Sinica. 2017, 37(23): 8101-8106
- http://dx.doi.org/10.5846/stxb201701130104
-
文章历史
- 收稿日期: 2017-01-13
- 修订日期: 2017-08-10





2. 哈尔滨师范大学, 哈尔滨 150025;
3. 宁夏回族自治区气象局, 银川 750002;
4. 山西省气候中心, 太原 030006
2. Harbin Normal University, Harbin 150025, China;
3. The Ningxia Hui Autonomous Region Meteorological Bureau, Yinchuan 750002, China;
4. Shanxi Climate Center, Taiyuan 030006, China
比叶面积(specific leaf area, SLA)指绿色叶面积与其干重之比[1], 其数值大小直接受叶片厚度、形状和重量等因素的影响。由于SLA与植物的生长状况和生存策略有着密切联系, 可以综合反映植物利用资源的能力以及对不同环境的适应性特征, 因而成为植物叶性状研究的首选指标[2]。同时, SLA是将叶生物量换算成叶面积的重要参数, 其模拟精度会极大地影响冠层光合作用和干物质生产模拟的准确性;近年来成为许多生态系统过程模型的一个重要输入参数[3]。因此, 开展SLA的相关研究不仅有助于阐明不同生境植物叶片生理生态对气候变化的响应机理[4], 而且对提高作物模型在产量预测和灾害损失评估等方面的精度都有着重要意义。
目前国内外学者围绕植物SLA已经开展了很多研究, 内容多侧重于不同生境植物SLA与叶片C、N、P等元素及干物质含量、土壤养分状况、不同水分梯度、光照强度等因子的相互关系[5-9]。李玉霖等[5]和韦兰英等[6]曾分别研究了沙丘和黄土高原不同退耕年限坡地的主要植物SLA, 认为不同生境植物SLA均存在显著差异。田青等[7]研究了内蒙古多伦典型草原14种植物SLA对水分梯度的响应, 发现不同水分梯度下典型草原植物SLA变化比较复杂, 二者并不存在简单线性关系。黄菊莹等[8]通过盆栽试验探讨了羊草(Leymus chinensis)SLA在不同N、P肥和水分梯度下的变化趋势, 发现一定范围内适量增加水分供给可提高绿叶SLA, 而降低绿叶N浓度。
玉米是自然界贮存能量最多的植物之一, 也是重要的粮食、饲料和经济兼用作物, 在我国北方干旱地区广泛种植。尽管目前围绕干旱如何影响玉米生长发育已经开展了许多水分控制试验, 对干旱胁迫状态下玉米SLA的变化趋势有一些初步结论[10], 但多以定性描述为主, 缺乏与土壤湿度的定量分析。土壤湿度是表征作物干旱发生程度的直接指标, 了解其与作物SLA的定量关系是建立相关模式的重要依据。因此, 本研究以河北固城农试站2013—2015年夏玉米田间水分控制试验为基础, 分不同发育时段探讨了夏玉米SLA对水分梯度的响应, 并建立了SLA与土壤湿度的定量关系, 以期为作物模型更加准确地模拟不同水分梯度下玉米生长发育状况提供理论依据。
1 材料与方法 1.1 试验设计在中国气象科学院河北固城农试站连续3年(2013—2015年)开展夏玉米田间水分控制试验。试验每年设立5个试验小区(单个小区面积约30m2), 每个试验小区设3个重复。试验采取遮雨棚遮避自然降水和灌溉的措施对夏玉米出苗-拔节、拔节-抽雄、抽雄-成熟3个主要生育阶段, 以及[(出苗-拔节)+(拔节-抽雄)]、[(出苗-拔节)+(抽雄-成熟)]、[(拔节-抽雄)+(抽雄-成熟)]3个复合生育阶段轮流进行水分控制。试验共设置4种水分梯度和1个对照处理, 夏玉米不同控水时段的土壤相对湿度(θ)分别控制为≤40%(T1)、40%—60%(T2)、60%—80%(T3)、>80%(T4)以及自然降水(T5)。
试验选用的玉米品种为郑单958;在播种前施底肥、拔节-孕穗前追肥1次, 保证土壤养分供应。试验采取人工播种方式, 按当地普遍播种时间进行播种, 行距50cm, 株距35cm, 深度约7cm, 密度约6株/m2。
1.2 试验观测项目 1.2.1 生育期观测根据农业气象观测规范, 判断并记录玉米播种、出苗、七叶、拔节、抽雄、乳熟以及成熟的日期。
1.2.2 土壤相对湿度观测统一采用土壤相对湿度(θ)来表征土壤的湿度状况, 即土壤含水量与田间最大持水量的百分比。利用土钻法从播种前一天开始观测直至收获, 每7日测定1次;并规定拔节前取样深度为0—20cm, 拔节后为0—50cm。首先根据烘干称重法求出土壤重量含水量, 然后用重量含水量除以田间最大持水量即可获得土壤相对含水量, 用土壤相对湿度(θ)来表示, 本研究中田间最大持水量取值22.7%。
1.2.3 SLA的测定在七叶、拔节、拔节后10d、拔节后20d、抽雄后10d、抽雄后20d以及成熟期, 按玉米长势好、中、差在每个小区分别随机选择1棵整株玉米, 采用Montgomery法[11]测定整株玉米的叶面积, 即利用刻度尺测定每株玉米展开叶片(绿叶)的叶长和最大叶宽, 乘以0.75, 求出各单叶面积后, 累加得全株总面积。最后将叶片放入72℃烘箱内烘48h至恒重后取出称重(干重)。此外, 为了增加样本量, 在拔节后30d和抽雄后30d还进行了加密观测。
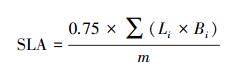
|
式中, L为叶长(cm), B为最大叶宽(cm), 下标i为叶片数, m为叶片生物量干重(g), 比叶面积SLA指单位质量的叶面积(cm2/g), 除以10换算为m2/kg。每个生育期的处理测定3次。
1.2.4 数据分析采用SPSS 19.0对夏玉米不同发育时段SLA与土壤相对湿度进行回归分析, 并对回归方程进行显著性检验, 给出检验统计量的概率P值范围和样本数n。
2 结果与分析 2.1 夏玉米SLA随发育期的变化图 1为水分条件适宜时(60%≤θ<90%), 夏玉米SLA随发育期的变化趋势。从七叶期开始夏玉米SLA随发育进程的推进总体呈下降趋势;其中七叶期到拔节期SLA迅速下降, 后期则维持稳定中略有下降。从SLA的3a均值来看, 七叶期最高(34.9m2/kg), 进入拔节期后SLA迅速下降, 维持在23.4m2/kg左右;抽雄期SLA略有下降, 为18.1—18.8m2/kg, 至成熟期夏玉米SLA降至最低, 平均为14.6m2/kg。

|
| 图 1 夏玉米SLA随发育期变化趋势(土壤水分条件适宜时) Fig. 1 The SLA change of summer maize with developing periods when soil water was sufficient |
图 2为不同水分处理下夏玉米地上绿叶面积、绿叶干重及SLA随发育期变化趋势。其中, T2为干旱胁迫处理(36%≤θ<60%), T3为水分适宜处理(60%≤θ<80%)。如图 2所示, 无论是否发生干旱, 夏玉米地上绿叶面积和绿叶干重在整个发育期的变化趋势基本一致。即绿叶面积和干重均在七叶期最低, 进入拔节期后迅速增大;不同的是T3在抽雄后10d绿叶面积达到最高(4019cm2), 对应的绿叶干重最高为22.7g, 而T2地上绿叶的生长明显受到干旱的抑制, 在抽雄后30d达到最大(1877cm2), 对应的绿叶干重最高为9.8g。图 2为T2、T3对应的SLA随发育期的变化趋势。两种水分条件下夏玉米SLA均在七叶期最高, 随着生育进程的推进SLA逐渐降低。不同的是, T2的SLA值整体略大于T3, 尤其是进入抽雄阶段之后, 这种变化趋势更为明显。由此初步推断, 尽管干旱胁迫会明显降低夏玉米地上绿叶面积和干重, 但SLA作为绿叶面积和干重的比值, 对干旱胁迫却表现出增大的趋势。

|
| 图 2 不同水分处理夏玉米绿叶面积、绿叶干重及SLA随发育期的变化趋势 Fig. 2 Changes of area, dry weight of green leaf and SLA of summer maize with developing periods under different water conditions |
图 3分发育时段探讨了夏玉米SLA与土壤相对湿度的关系, 并建立了回归方程。出苗-拔节期和抽雄-成熟期夏玉米SLA均随θ的增加呈下降趋势, 也就是说θ越小、干旱胁迫越严重, SLA越大;二者表现为显著和极显著的线性负相关关系(P值分别<0.01和<0.001)。但这两个时段夏玉米对干旱的“响应”能力存在差异。规定土壤相对湿度θ每下降1%时夏玉米生长生理生态指标的变化为干旱影响系数[12]。出苗-拔节期干旱影响SLA的系数为0.45m2/kg;抽雄-成熟期干旱影响SLA的系数仅为0.07m2/kg。而拔节-抽雄期的SLA与θ不是简单的线性关系, 表现为先增大后减小的抛物线型, 采用二次函数进行拟合, 拟合结果同样通过了显著性检验(P < 0.05)。

|
| 图 3 夏玉米不同发育时段SLA与土壤相对湿度的关系 Fig. 3 Relationship between SLA and soil relative moisture during different developing periods of summer maize |
文中河北固城夏玉米2013—2015年七叶至成熟期SLA变化范围平均为14.6—34.9m2/kg, 与前人的观测结果较为接近。刘铁梅等[13]认为冬油菜各生育期之间(除越冬期外)SLA差异显著, 变化范围为11.9—41.6m2/kg。乔玉辉等[14]通过田间试验发现越冬前冬小麦SLA基本保持在25m2/kg, 返青后从最大值40m2/kg逐渐减少到灌浆期的最小值18—20m2/kg。植物SLA随发育期的变化间接反映了不同发育阶段光合产物的积累与运转会随着生长中心的转移、更替而发生变化[15]。夏玉米从出苗开始进入营养生长阶段, 植物光合作用产生的大量营养物质主要供于叶片扩展, 叶面积不断增大, SLA也随之增加, 至七叶期达到顶峰;进入拔节期后, 茎秆开始形成, 进入营养生长和生殖生长并行的阶段, 叶面积增长放缓, SLA维持或略有下降;抽雄至成熟期以生殖生长为主, 叶片中合成的大量葡萄糖被转运到穗等贮存器官, 叶片开始衰老, SLA逐渐下降。刘铁梅等[13]的研究也发现了相似的规律, 冬油菜SLA在开花期最大, 进入角果发育期后随着光合产物逐渐向经济器官转运, SLA逐渐下降。
干旱胁迫会明显降低夏玉米地上绿叶面积和干重, 导致夏玉米生育进程延缓, 类似的结论已经在相关文献中提及[16-17]。本研究中夏玉米出苗-拔节期和抽雄-成熟期SLA与土壤相对湿度显著负相关, 干旱胁迫使SLA表现出“补偿性”增大。SLA增大即叶片变大、叶片变薄, 叶片变大可以吸收更多光能从而提高整株植物的光合产物, 在一定程度上弥补干旱胁迫导致光合产物不足引起的叶片扩张乏力, 这可能是玉米生长对逆境的适应。不同的是, 出苗-拔节期干旱影响SLA的系数为0.45m2/kg, 较抽雄-成熟期(0.07m2/kg)偏高, 这说明出苗-拔节期发生干旱胁迫时玉米叶片“补偿性”扩展的能力更大, 这也从侧面解释了为什么营养生长阶段的干旱对玉米产量的影响小于生殖生长阶段[18]。马玉平等[12]认为干旱对夏玉米叶面积扩展有明显的正效应, 土壤相对湿度距适宜湿度减小1%, SLA会上升0.8m2/kg, 与本文的结论在趋势上是一致的。张丛志等[10]利用盆栽试验也曾观测到孕穗期前严重水分胁迫下的SLA与轻度水分胁迫下的SLA比较接近, 但孕穗期后严重水分胁迫处理下的SLA显著高于轻度水分胁迫处理的现象。但也有部分观点认为, 水分胁迫会影响叶片展开, 阻碍同化物质从叶片中运输出去, 从而导致叶片变厚、SLA变小[19]。较厚的叶片中单位叶面积内的RUBP羧化酶通常比薄叶多, 因而叶片最大光合速率可能随叶片变厚而增加[20]。但若光合产物量不变, 则薄叶的面积更大可促使植物吸收更多的光能, 整株光合产物会增多。两者哪个更有利于产生更多光合产物尚需进一步探讨。拔节-抽雄期SLA变化比较复杂, 表现为先增大后减小的抛物线型。原因可能是拔节至抽雄期是夏玉米从营养生长向生殖生长转化的阶段, 植物内在的生长中心转移、干物质分配以及出现干旱胁迫时植物的生长策略等因素都会对SLA产生影响, 内在调节机制还需开展更多的生理生态方面的试验进一步研究确定。
3.2 结论从七叶期开始河北固城夏玉米SLA随发育进程的推进总体呈下降趋势, 2013—2015年夏玉米七叶至成熟期SLA变化范围平均为14.6—34.9m2/kg。尽管干旱胁迫会明显降低夏玉米地上绿叶面积和干重, 但却使SLA表现出“补偿性”增大。夏玉米出苗-拔节期、抽雄-成熟期SLA与土壤相对湿度呈显著的线性负相关关系, 干旱影响SLA的系数分别为0.45m2/kg和0.07m2/kg, 说明出苗-拔节期发生干旱胁迫时夏玉米叶片“补偿性”扩展的能力更大, 对干旱的适应能力更强;而拔节-抽雄期SLA与土壤水分的关系较为复杂, 表现为先增大后减小的抛物线型。
| [1] | Gower S T, Kucharik C J, Norman J M. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sensing of Environment, 1999, 70(1): 29–51. DOI:10.1016/S0034-4257(99)00056-5 |
| [2] | Garnier E, Shipley B, Roumet C, Laurent G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 2001, 15(5): 688–695. DOI:10.1046/j.0269-8463.2001.00563.x |
| [3] | Landsberg J J, Gower S T. Applications of Physiological Ecology to Forest Management. San Diego: Academic Press, 1997: 354–354. |
| [4] | Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, Van Der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335–380. DOI:10.1071/BT02124 |
| [5] | 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 2005, 25(2): 304–311. DOI:10.17521/cjpe.2005.0039 |
| [6] | 韦兰英, 上官周平. 黄土高原不同退耕年限坡地植物比叶面积与养分含量的关系. 生态学报, 2008, 28(6): 2526–2535. |
| [7] | 田青, 曹致中, 王国宏. 内蒙古多伦典型草原14种植物比叶面积对水分梯度变化的响应. 草原与草坪, 2008(5): 23–26. |
| [8] | 黄菊莹, 袁志友, 李凌浩. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化. 植物生态学报, 2009, 33(3): 442–448. |
| [9] | 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844–852. DOI:10.17521/cjpe.2004.0110 |
| [10] | 张丛志, 张佳宝, 赵炳梓, 张辉, 黄平, 李晓鹏, 朱强根. 玉米水分利用效率、碳稳定同位素判别值和比叶面积之间的关系. 作物学报, 2009, 35(6): 1115–1121. |
| [11] | 王得贤. 四种测定单株玉米总叶面积方法的比较. 青海农林科技, 1999(4): 20–21. |
| [12] | 马玉平、孙琳丽、马晓群. 黄淮海地区夏玉米对干旱和涝渍的生理生态反应. 干旱地区农业研究, 2016, 34(4): 85–93. DOI:10.7606/j.issn.1000-7601.2016.04.13 |
| [13] | 刘铁梅, 邹薇, 刘铁芳, 谢国生, 吴江生, 曹卫星, 李卫平, 曹凑贵, 傅廷栋. 不同冬油菜品种比叶面积的多因子分析. 作物学报, 2006, 32(7): 1083–1089. |
| [14] | 乔玉辉, 宇振荣, DriessenP M. 冬小麦叶面积动态变化规律及其定量化研究. 中国生态农业学报, 2002, 10(2): 83–85. |
| [15] | Roche P, Díaz-Burlinson N, Gachet S. Congruency analysis of species ranking based on leaf traits:which traits are the more reliable?. Plant Ecology, 2004, 174(1): 37–48. DOI:10.1023/B:VEGE.0000046056.94523.57 |
| [16] | Ribaut J M, Hoisingon D A, Deutsch J A, Jiang C, Gonzalez-de-Leon D. Identification of quantitative trait loci under drought con ditions in tropical maize. 1. Flowering parameters and the anthesis-silking interval. Theoretical and Applied Genetics,, 1996, 92(7): 905–914. DOI:10.1007/BF00221905 |
| [17] | 陶世蓉, 东先旺, 张海燕, 刘树堂, 刘培利. 土壤水分胁迫对夏玉米植株性状整齐度的影响. 西北植物学报, 2000, 20(5): 812–817. |
| [18] | 刘浏. 干旱对玉米生长发育及产量的影响. 农业科技与装备, 2015(11): 6–7. DOI:10.3969/j.issn.1674-1161.2015.11.003 |
| [19] | Bagnall D J, King R W, Farquhar G D. Temperature-dependent feedback inhibition of photosynthesis in peanut. Planta, 1988, 175(3): 348–354. DOI:10.1007/BF00396340 |
| [20] | De Vries F W T P, Jansen D M, ten Berge H F M, Bakema A. Simulation of Ecophysiological Processes of Growth in Several Annual Crops. Wageningen, the Netherlands:Pudoc, 1989: 271–271. |